Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Ebola virus disease wikipedia , lookup
Social history of viruses wikipedia , lookup
Plant virus wikipedia , lookup
Human Endogenous Retrovirus-W wikipedia , lookup
Introduction to viruses wikipedia , lookup
Endogenous retrovirus wikipedia , lookup
Negative-sense single-stranded RNA virus wikipedia , lookup
History of virology wikipedia , lookup
Oncolytic virus wikipedia , lookup
GENETICS AND FUNCTIONS OF HERPES SIMPLEX VIRUS TYPE 1
MEMBRANE PROTEINS IN VIRUS-INDUCED CELL FUSION, VIRION
MORPHOGENESIS AND EGRESS
A Dissertation
Submitted to the Graduate Faculty of the
Louisiana State University and
Agricultural and Mechanical College
in partial fulfillment of the
requirements for the degree of
Doctor of Philosophy
in
The Interdepartmental Program in
Veterinary Medical Sciences through the
Department of Pathobiological
Sciences
By
Jeffrey M. Melancon
B.Sc. Louisiana State University, 2000
December, 2004
© Copyright 2004
Jeffrey Michael Melancon
All Rights Reserved
ii
ACKNOWLEDGEMENTS
First, and foremost, I would like to thank my lovely wife Kelly for being
supportive of my goals and tolerating the long and strange hours that I spent in the
laboratory during these first few years of our marriage. I would also like to thank my
family for their encouragement and steadfast support of my pursuits.
I would like to express my gratitude to Dr. Timothy Foster, who provided
invaluable assistance and suggestions to aid in my dissertation research. In addition, I
owe thanks to all members of the Kousoulas laboratory who gave helpful advice
whenever asked, with special thanks given to Chad Petit and Rafael Luna, who helped to
shape many ideas and address the difficulties that we shared in pursuit of our goals. I
would like to thank the members of GeneLab, Thaya Geudry and Mamie Burrell, for their
assistance in DNA sequencing and primer synthesis.
I would like to thank the faculty members that served on my graduate committee,
Dr. Thomas Klei, Dr. John Larkin, Dr. Patrick DiMario, and Dr. John Chandler, for their
recommendations and advice in shaping my dissertation research and my learning
experience. Finally, I would like to thank Dr. K.G. Kousoulas for providing guidance,
encouragement, and support in the time that I have spent as a graduate student in his
laboratory, without which my development and successes in graduate school would not
have been possible.
iii
TABLE OF CONTENTS
ACKNOWLEDGEMENTS ............................................................................................. iii
LIST OF TABLES .......................................................................................................... viii
LIST OF FIGURES .......................................................................................................... ix
ABSTRACT.................................................................................................................... xvii
CHAPTER I: INTRODUCTION .....................................................................................1
STATEMENT OF PROBLEM AND HYPOTHESIS.................................................1
STATEMENT OF RESEARCH OBJECTIVES .........................................................3
LITERATURE REVIEW ..............................................................................................5
Historical Perspective of Herpesviruses........................................................5
Taxonomy of Herpesviridae ............................................................................7
Clinical Significance of Herpes Simplex Viruses .........................................9
Epidemiology................................................................................................9
Pathogenesis................................................................................................10
Mucocutaneous Infections ......................................................................10
Fetal and Neonatal Infections .................................................................12
Keratoconjunctivitis................................................................................12
Infection of an Immunocompromised Host ............................................13
Central Nervous System (CNS) Infections.............................................13
Prevention and Treatment of HSV Infection ..............................................14
Architecture of the Herpes Virion...............................................................14
The Core......................................................................................................14
The Capsid ..................................................................................................16
The Tegument .............................................................................................16
The Envelope ..............................................................................................17
Organization of the Viral Genome ..............................................................17
The Herpes Simplex Virus Lifecycle ...........................................................19
Virus Attachment and Entry .......................................................................19
Binding Receptors.......................................................................................22
Heparan Sulfate (Glycosaminoglycans) .................................................22
Entry Receptors...........................................................................................22
Tumor Necrosis Factor (TNF) Receptor Family ....................................22
Immunoglobulin Superfamily.................................................................24
3-O-sulfated Heparan Sulfate (3-OS HS) ...............................................25
Other Herpesvirus Receptors ..................................................................26
Virus-to-Cell Fusion ...............................................................................26
Host Protein Shutoff ...................................................................................27
Virion Transport to the Nucleus .................................................................28
Coordinate Gene Expression.......................................................................29
iv
Viral DNA Replication ...............................................................................33
Capsid Assembly and Packaging ................................................................35
Herpesvirus Egress......................................................................................37
Nuclear Egress: Primary Envelopment...................................................37
Egress from the Perinuclear Space: De-Envelopment............................40
Tegumentation in the Cytoplasm............................................................42
Final Envelopment at the Trans-Golgi Network/Endosomes and
Egress to Extracellular Spaces................................................................43
Formation of Light (L) Particles.............................................................45
HSV-1 Glycoproteins and Their Putative Functions .................................46
Glycoprotein B............................................................................................46
Glycoprotein C............................................................................................47
Glycoprotein D............................................................................................49
Glycoproteins E and I .................................................................................50
Glycoprotein G............................................................................................52
Glycoproteins H and L................................................................................52
Glycoprotein J.............................................................................................54
Glycoprotein K............................................................................................54
Glycoprotein M and UL49.5 (gN) ..............................................................57
Characterization of the UL20 Protein (UL20p) .........................................58
The UL20 ORF ...........................................................................................58
Membrane Topology of UL20p ..................................................................59
Interdependence with gK for Cell Surface Expression and
Internalization to the Trans-Golgi Network................................................60
Function of UL20p in the HSV-1 Lifecycle ...............................................60
Virus-Induced Syncytia Formation.............................................................61
REFERENCES..............................................................................................................63
CHAPTER II: AN α-HELICAL DOMAIN WITHIN THE CARBOXYL
TERMINUS OF HERPES SIMPLEX VIRUS TYPE 1 (HSV-1)
GLYCOPROTEIN B (gB) IS ASSOCIATED WITH CELL FUSION
AND RESISTANCE TO HEPARIN INHIBITION OF CELL
FUSION .................................................................................................................94
INTRODUCTION ........................................................................................................94
MATERIALS AND METHODS .................................................................................97
Cells and Viruses...........................................................................................97
Construction of Expression Plasmids..........................................................97
Cell Fusion Assay. .........................................................................................98
Immunofluorescence Assay and Quantitation of Fusion. .........................99
Heparin Inhibition of Viral Polykaryocyte Formation. ............................99
RESULTS ....................................................................................................................100
The Effect of gB Truncations on Virus-Independent Cell Fusion..........100
The Role of α-helical Domain H17b in Cell Fusion. ................................105
The Effect of Heparin on gB-Mediated Cell Fusion. ...............................105
The Effect of Heparin on Virus-Induced Cell Fusion..............................110
DISCUSSION ..............................................................................................................111
v
REFERENCES............................................................................................................118
CHAPTER III: HERPES SIMPLEX VIRUS TYPE 1 (HSV-1)
GLYCOPROTEIN K (gK) IS REQUIRED FOR GLYCOPROTEIN
B (gB)-MEDIATED VIRUS-INDUCED CELL FUSION, WHILE
NEITHER gB AND gK NOR gB AND UL20p FUNCTION
REDUNDANTLY IN VIRION DE-ENVELOPMENT...................................125
INTRODUCTION ......................................................................................................125
MATERIALS AND METHODS ...............................................................................128
Cells and Viruses.........................................................................................128
Plasmids .......................................................................................................128
Construction of Transformed Flp-In-CV-1 Cell Lines............................129
Construction of a Replication-Deficient Adenovirus Expressing
gB...........................................................................................................130
Construction of HSV-1 Mutants with Deletions of the UL20, gB,
and/or gK Genes...................................................................................130
Confirmation of the Targeted Mutations in pYEbac102 DNA...............132
Transfection of HSV-1 BAC DNAs. ..........................................................132
Plaque Morphology of YEbac102 Mutants. .............................................133
One-step Growth Kinetics of YEbac102 Mutants....................................133
Generation of Recombinant UL20-null and gK-null Viruses
Specifying Mutant gB. .........................................................................134
Generation of UL19 and UL20 Double-Null Viruses...............................134
Electron Microscopy...................................................................................135
RESULTS ....................................................................................................................135
Construction of the HSV-1 BACs pYEbac∆gB, pYEbac∆UL20,
pYEbac∆gK, pYEbac∆gB∆UL20, and pYEbac∆gB∆gK. ................135
Generation of Infectious Virus from pYEbac102-Based
Constructs.............................................................................................142
Plaque Morphology and Growth Kinetics of HSV-1 YEbac102
Mutants. ................................................................................................143
Ultrastructural Characterization of the YEbac∆gB∆UL20 and
YEbac∆gB∆gK Double-Null Mutant Viruses....................................148
Generation and Characterization of Recombinant gK-null and
UL20-null viruses Containing the gBsyn3 or gBamb1511
Mutation................................................................................................152
Virion Assembly and Egress Is Not Required for Either gB or gKMediated Virus-Induced Cell-to-Cell Fusion ....................................154
DISCUSSION ..............................................................................................................156
HSV-1 BAC Generated Recombinant Viruses.........................................156
gB, UL20p and gK are not Required for Virion De-Envelopment.........157
Role of gK on gB-Mediated Cell-to-Cell Fusion.......................................160
Virion Egress is not Required in Virus-Induced Cell Fusion. ................161
REFERENCES............................................................................................................162
vi
CHAPTER IV: GENETIC ANALYSIS OF THE HERPES SIMPLEX
VIRUS TYPE 1 (HSV-1) UL20 PROTEIN DOMAINS INVOLVED
IN CYTOPLASMIC VIRION ENVELOPMENT AND VIRUSINDUCED CELL FUSION................................................................................168
INTRODUCTION ......................................................................................................168
MATERIALS AND METHODS ...............................................................................171
Cells and Viruses.........................................................................................171
Construction of Transformed Flp-In-CV-1 Cell Lines............................171
Plasmids. ......................................................................................................172
UL20 Complementation Assay for Infectious Virion Production. .........173
Calculation of Complementation Ratios...................................................174
UL20 Complementation Assay for Virus-Induced Cell-to-Cell
Fusion. ...................................................................................................174
Electron Microscopy...................................................................................175
Generation of Recombinant Viruses Specifying Mutant UL20p............175
RESULTS ....................................................................................................................176
Mutagenesis of HSV-1 UL20......................................................................176
Complementation Assay for Infectious Virus Production. .....................179
Complementation for Virus-Induced Cell-to-Cell Fusion.......................182
Plaque Phenotypes of ∆20gBsyn3 and ∆20gKsyn1 Recombinants
Containing Selected UL20 Mutations. ...............................................187
Plaque Phenotypes of Selected UL20 Mutations in the Context of
Wild-Type gB and gK Genes. .............................................................189
Ultrastructural Characterization of Recombinant Viruses
Carrying Mutations Within the Amino and Carboxyl Termini
of UL20p. ..............................................................................................191
DISCUSSION ..............................................................................................................195
The Predicted Membrane Topology of UL20p. .......................................195
UL20p Domains that Function in Infectious Virus Production and
Virus-Induced Cell Fusion. .................................................................196
UL20p Mutations that Cause Virus-Induced Cell Fusion.......................201
REFERENCES............................................................................................................203
CHAPTER V: CONCLUDING REMARKS...............................................................208
SUMMARY .................................................................................................................208
CURRENT AND FUTURE RESEARCH CHALLENGES....................................212
FINAL COMMENTS .................................................................................................223
REFERENCES............................................................................................................227
APPENDIX: LETTERS OF PERMISSION ...............................................................229
VITA.................................................................................................................................232
vii
LIST OF TABLES
Table 1.1: Members of the family Herpesviridae .........................................................8
Table 2.1: Truncation of the carboxyl terminal 28 amino acid of gB causes
extensive fusion of transfected Cos cells..........................................................103
Table 2.2A: gB-H17b-targeted mutations enhance fusion of transfected Cos
cells .....................................................................................................................109
Table 2.2B: gB-H17b-targeted mutations confer resistance to heparin
inhibition of cell fusion......................................................................................109
Table 3.1: Synthetic Oligonucleotide Primers............................................................137
Table 4.1: Amino acid sequences of mutations .........................................................177
Table 4.2: Complementation results for cell fusion ...................................................184
Table 4.3: Differential effects of mutations on infectious virion production
and cell fusion ....................................................................................................197
viii
LIST OF FIGURES
Figure 1.1: Herpesvirus virion structure. Virions of herpes viruses can vary in
size from 120nm to 300nm (Roizman and Furlong, 1974). A virion
consists of: an electron-dense core containing the viral genome, an
icosadeltahedral capsid around the core, an amorphous tegument around
the capsid, and an envelope derived from cellular membranes containing
glycoprotein spikes (Roizman and Furlong, 1974) ...............................................15
Figure 1.2: Arrangement of the HSV-1 genome. (A) The top line represents
the prototypic arrangement of the HSV-1 genome with the unique long
(UL) and unique short (US) regions flanked by the terminal repeat (TR)
and internal repeat (IR) regions. (B) The bottom line shows map units of
the HSV-1 genome................................................................................................18
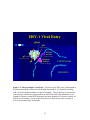
Figure 1.3: The Herpes Simplex Virus Life Cycle. The first stage of the herpes
virus life cycle consists of virus entry, capsid transport to the cell nucleus,
deposition of viral DNA into the nucleoplasm, coordinate gene expression
and viral DNA replication (black arrows). The second stage is virion
morphogenesis and egress comprised of primary envelopment at the inner
nuclear membrane, de-envelopment at the outer nuclear membrane, final
envelopment into cytoplasmic vesicles and transport to extrancellular
spaces (red arrows)................................................................................................20
Figure 1.4: Herpes Simplex Virus Entry. The first step of HSV entry is
attachment to the plasma membrane of infected cells through interaction
of gC (ligand for binding) with cell surface heparan sulfate (receptor for
binding). The second step is fusion of the viral envelope with the cell
plasma membrane and occurs following attachment of gD (ligand for
fusion) to either HVEM or a nectin (receptor for fusion) on cell surfaces.
Fusion requires the presence of gD and an entry receptor, as well as gB
and the gH/gL heterodimer. ..................................................................................21
Figure 1.5: Herpes Simplex Virus entry receptors. The three classes of cell
surface receptors for HSV entry are: the tumor necrosis factor (TNF)
receptor family consisting of HVEM, the immunoglobulin superfamily
consisting of the nectins, and 3-O-sulfated heparin sulfate. Only viral
attachment can occur in the absence of an HSV entry receptor............................23
Figure 1.6: Coordinate gene expression in Herpes Simplex Virus. 1) α-TIF, a
γ gene present in the tegument, activates initial transcription of the
α genes. 2) Autoregulation of gene expression. 3) Activation of β gene
expression. 4) Activation of γ gene expression by α and β genes, release of
ix
repression of γ genes, and replication of the viral genome. 5) γ genes turn
off α and β genes late in infection (Roizman and Knipe, 2001)...........................31
Figure 1.7: Representation of HSV-1 virion morphogenesis and egress. (I)
Mature capsids budding through the inner nuclear membrane into the
perinuclear space; (II) De-envelopment of perinuclear virions at the outer
nuclear membrane; (III) Re-envelopment of cytoplasmic capsids by
budding into cytoplasmic vesicles; (IV) Final egress to the extracellular
space. The steps at which UL20 and gK are thought to function in virion
egress are indicated by red arrows ........................................................................38
Figure 2.1: Schematic diagram of the carboxyl terminus of HSV-1 gB. The
predicted secondary structure of the gB carboxyl terminal 109 amino acids
is shown as described previously (Baghian et al., 1993; Pellett et al.,
1985). The location of the gB truncations and mutations specified by
plasmids p9-1513, p9-1528, p9-1511, pCMV-gB(∆28), pCMV-gB(∆36),
pCMV-gB(A874P), pCMV-gB(∆16), pCMV-gB(syn3) are shown. The
symbol ∆ followed by a number indicates the number of amino acids
deleted from the carboxyl terminus of gB. The location of the two
predicted α−helices H17a and H17b are shown and marked by dashed
lines .....................................................................................................................101
Figure 2.2: Truncation of the carboxyl terminal 28 amino acid of gB causes
extensive fusion of transfected Cos cells. Cells were transfected with
plasmids expressing gD, gH/gL, and plasmid A: pCMV-gB, B: p9-2080,
C: p9-1537, D: p9-1513, E: p9-1528, F: p9-1511. Transfected cells were
visualized by IFA at 36 hours after transfection (65X magnification) ...............102
Figure 2.3: Quantitation of the effect of gB truncation on fusion of
transfected Cos Cells. Nuclei in 100 IFA positive cells per assay were
counted by two observers and the average values and standard deviation
were calculated....................................................................................................103
Figure 2.4: gB-H17b-targeted mutations enhance fusion of transfected Cos
cells. Cells transfected with plasmids expressing gD, gH/gL, and A: gB,
B: gB(syn3), C: gB(∆16), D: gB(∆28), E: gB (∆36), and F: gB (A874P).
Transfected cells were visualized by IFA at 36 hours after transfection at
65X magnification...............................................................................................106
Figure 2.5: gB-H17b-targeted mutations confer resistance to heparin
inhibition of cell fusion. Cells transfected with plasmids expressing gD,
gH/gL, and A: gB, B: gB(syn3), C: gB(∆16), D: gB(∆28), E: gB (∆36),
and F: gB (A874P). Cells were incubated in the presence of heparin.
Transfected cells were visualized by IFA at 36 hours after transfection at
65X magnification...............................................................................................107
x
Figure 2.6: Quantitative comparison of fusion of transfected cells in the
presence and absence of heparin. Cell fusion represented in figure 4
(without heparin) and figure 2.5 (with heparin) was quantitated as
described in the legend of figure 3......................................................................108
Figure 2.7: Cell fusion caused by viruses specifying H17b mutated gB is
resistant to heparin inhibition. Vero cells were infected at an MOI of
0.1 with KOS/EGFP (A, B), tsb5/EGFP (C, D), amb1511/EGFP (E,F) and
MP/EGFP (G, H) in the presence (B, D, F, H) or absence (A, C, E, G) of
heparin. Infected cells were visualized by fluorescence microscopy (65X
magnification) .....................................................................................................112
Figure 3.1: Schematic of the strategy for the construction of pYEbac102
mutant BACs. (A) The top line represents the prototypic arrangement of
the HSV-1 genome with the unique long (UL) and unique short (US)
regions flanked by the terminal repeat (TR) and internal repeat (IR)
regions. (B) Shown below are the expanded genomic regions of the UL20,
UL27, and UL53 ORFs as well as the approximate locations of the sites
where insertion of the marker genes was targeted and the primers used in
diagnostic PCR to confirm the presence of each mutation. (C) PCR
fragments containing the kanamycin or GFP-Zeocin gene cassette flanked
by ~50 bp of viral sequences on both sides were used for targeted GET
recombination in E.coli to construct pYEbac102 mutant BACs with
insertion-deletion mutations in the UL20, UL27, and/or UL53 ORFs. The
approximate location of the primers used in amplification of each PCR
fragment are also shown......................................................................................138
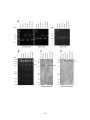
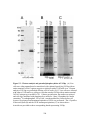
Figure 3.2: Genomic analysis of pYEbac102 mutants. (A) Diagnostic PCR to
confirm the insertion of the relevant gene cassettes in place of the deleted
genomic regions in UL20 (A, panel i) UL27 (gB) (A, panel ii), and UL53
(gK) (A, panel iii). (B) KpnI restriction fragment analysis of pYEbac102
mutant BACs in comparison to the wild-type pYEbac102. The black
arrow demarcates the position of the KpnI DNA fragment altered by the
∆gB-Kan mutation, while the gray arrow points to the missing 9,432 bp
fragment resulting from the ∆UL20-GFPZeo mutation. (C, D) Southernblot analysis of pYEbac102 mutant BACs. The KpnI restricted BACs
from Figure 2B were hybridized with either a kanamycin (C) or GFPZeocin (D) biotinylated probe. The biotinylated kanamycin probe
hybridized to an estimated 4,835 bp DNA fragment in pYEbac∆gB,
pYEbac∆gB∆gUL20 and pYEbac∆gB∆gK DNA, which was absent in the
wild-type pYEbac102 DNA (indicated by a black arrow in panel C). The
biotinylated GFP-Zeocin probe identified a predicted 10,992 bp DNA
fragment (gray arrow) in pYEbac∆UL20 and pYEbac∆gB∆gUL20 DNA
corresponding to the ∆UL20-GFPZeo mutation, and a predicted 8,956 bp
fragment (white arrow) in pYEbac∆gK and pYEbac∆gB∆gK DNA
corresponding to the ∆gK-GFPZeo mutation (D)...............................................140
xi
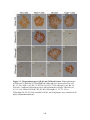
Figure 3.3: Plaque phenotypes of gB, gK and UL20-null viruses. Plaque
phenotypes of YEbac102, YEbac∆UL20, YEbac∆gB and YEbac∆gK were
observed on Vero (A1, B1, C1, D1), Fd20-1 (A2, B2, C2, D2), D6 (A3,
B3, C3, D3) and FcgK-1 (A4, B4, C4, D4) cells. Confluent cell
monolayers were infected with the wild-type YEbac102 (A1, A2, A3, A4),
YEbac∆UL20 (B1, B2, B3, B4), YEbac∆gB (C1, C2, C3, C4) or
YEbac∆gK (D1, D2, D3, D4) at an MOI of 0.001, and viral plaques were
visualized at 48 hpi by immunohistochemistry...................................................144
Figure 3.4: Plaque phenotypes of double-null viruses and rescued viruses.
Plaque phenotypes of YEbac∆gB∆gUL20 and YEbac∆gB∆gK on Vero
(A1, B1), Fd20-1 (A2, B2), D6 (A3, B3), and FcgK-1 (A4, B4) cells, or
Ad5-gB infected Fd20-1 (A5, B5) and FcgK-1 (A6, B6) cells are shown.
Confluent cell monolayers were infected with YEbac∆gB∆gUL20 (A1,
A2, A3, A4, A5, A6) or YEbac∆gB∆gK (B1, B2, B3, B4, B5, B6) at an
MOI of 0.001. The plaque phenotypes of the rescued gB, gK and UL20null (D, E, F) and double-null (G, H) viruses in comparison to the wildtype YEbac102 (C) are shown on Vero cells. Viral plaques were
visualized at 48 hpi by immunohistochemistry...................................................146
Figure 3.5: Viral replication kinetics. Comparison of virus replication
characteristics of YEbac102 (●),YEbac∆UL20 (■),YEbac∆gK
(▲),YEbac∆gB (○),YEbac∆gB∆UL20 (□), and YEbac∆gB∆gK (∆) on
Vero cells. One step kinetics of infectious virus production were
calculated after infection at an MOI of 5 followed by incubation at 37°C .........147
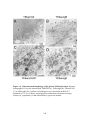
Figure 3.6: Ultrastructural morphology of gB, gK and UL20-null viruses.
Electron micrographs of Vero cells infected with YEbac102 (A),
YEbac∆gB (B), YEbac∆UL20 (C) or YEbac∆gK (D). Confluent cell
monolayers were infected at an MOI of 5, incubated at 37°C for 24 hours,
and prepared for transmission electron microscopy. Nuclear (n),
cytoplasmic (c) and extracellular (e) spaces are marked ....................................149
Figure 3.7: Ultrastructural morphology of gB/gK and gB/UL20 double-null
viruses. Electron micrographs of Vero cells infected with
YEbac∆gB∆gUL20 (A1, A2, A3, A4, A5) or YEbac∆gB∆gK (B1, B2, B3,
B4). Confluent cell monolayers were infected at an MOI of 5, incubated at
37°C for 24 hours, and prepared for transmission electron microscopy.
Panels A4 and B3 show partially-enveloped capsids. Panels A5 and B4
show a tegument-like accumulation on membranes that are folded
irregularly. Nuclear (n), cytoplasmic (c) and extracellular (e) spaces are
marked.................................................................................................................150
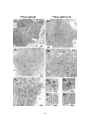
Figure 3.8: Plaque morphology of gK and UL20-null viruses carrying
syncytial mutations in gB. Plaque phenotypes of gK-null (A, B, C, D, E,
xii
F) and UL20-null (G, H, I, J, K, L) recombinant viruses containing
specific gB mutations on Vero (A, C, E, G, I, K) and FcgK-1 (B, D, F) or
Fd20-1 (H, J, L) cells. Confluent cell monolayers were infected with the
gK-null viruses containing: wild-type gB (A, B), gBsyn3 (C, D), or
gBamb1511 (E, F); or UL20-null viruses containing wild-type gB (G, H),
gBsyn3 (I, J), or gBamb1511 (K, L) at an MOI of 0.001, and viral plaques
were visualized at 48 hpi by immunohistochemistry. Insets are shown at
2.4x magnification to facilitate visualization of the non-syncytial
phenotype of the small plaques ...........................................................................153
Figure 3.9: Plaque phenotypes of ∆1920, ∆1920-syn20 and ∆1920-syn3
viruses on Vero (A, C, E) and FcgK-1 (B, D, F) cells. Confluent cell
monolayers were infected with the ∆1920 (A, B), ∆1920-syn20 (C, D) or
∆1920-syn3 (E, F) at an MOI of 0.001, and viral plaques were visualized
at 48 hpi by immunohistochemistry....................................................................155
Figure 3.10: Schematic of HSV-1 virion morphogenesis and egress. (I)
Mature capsids budding through the inner nuclear membrane into the
perinuclear space; (II) De-envelopment of perinuclear virions at the outer
nuclear membrane; (III) Re-envelopment of cytoplasmic capsids by
budding into cytoplasmic vesicles; (IV) Final egress to the extracellular
space....................................................................................................................159
Figure 4.1: Predicted membrane topology of UL20p and location of the 15
cluster-to-alanine mutations. Membrane topology was predicted using
the TMPred and SOSUI algorithms (19, 20). UL20p domains where
cluster-to-alanine mutations are located are indicated by a shaded oval.
Naming of cluster mutations is based on the first amino acid mutated in
each cluster. Transmembrane region (TM), Cluster mutant (CL) ......................178
Figure 4.2: Schematic of the strategy for the construction of UL20-mutant
genes used for complementation and isolation of recombinant viruses.
(A) The top line represents the prototypic arrangement of the HSV-1
genome with the unique long (UL) and unique short (US) regions flanked
by the terminal repeat (TR) and internal repeat (IR) regions. (B) Shown
below is and expanded genomic region of the UL20-null virus between
map units 0.239 and 0.305 containing the UL19, CMV-EGFP, UL20.5,
UL21, and UL22 genes. (C) The p20F2BX-UL20mut plasmid denotes the
expression and recombination plasmids used for complementation assays
and construction of the recombinant viruses carrying each UL20 mutation.
(D, E) Mutated UL20 genes were inserted into the BamHI restriction site
of p20F2BX.........................................................................................................180
Figure 4.3: Complementation ratios of mutant UL20p genes. Vero cells were
transfected with plasmids encoding wild-type or mutant UL20 genes under
the UL20 promoter and then infected with the HSV-1(KOS) UL20-null
xiii
(∆20) virus. (A) Bar graph showing complementation ratios for UL20
cluster-to-alanine mutants and carboxyl terminal truncations. (B)
Complementation ratios of the CL49, Y49A, S50A, and R51A mutants.
(C) Complementation ratios of the CL153, E153A, T154A, S156A, and
D158A mutants. (D) Complementation ratios of the CL209, R209A,
T212A, and R213A mutants. The error bars shown represent the
maximum and minimum complementation ratios obtained from three
independent experiments, and the bar height represents the average
complementation ratio.........................................................................................181
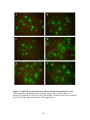
Figure 4.4: Complementation for virus-induced cell-to-cell fusion of UL20null viruses containing syncytial mutations in gK(syn1) or gB(syn3).
Vero cells were transfected with plasmids encoding wild-type or mutant
UL20 genes and then infected with either the ∆20gKsyn1 or ∆20gBsyn3
viruses. Twenty-four hpi, cell fusion was determined by visualization of
syncytia formation by fluorescence microscopy. The extent of syncytial
formation (complementation) obtained with the negative control plasmid,
p∆20NE (A) and positive control plasmid, p20R (B) are shown for
reference purposes. Representative images for the CL38, 204t, 211t, and
216t mutants (C, E, G, I, respectively), and for the CL49, Y49A, S50A,
and R51A (D, H, F, J, respectively) are shown...................................................185
Figure 4.5: Plaque phenotypes of ∆20gKsyn1 (A, B, C, D, E, F) and
∆20gBsyn3 (G, H, I, J, K, L) derived recombinant viruses containing
selected UL20 mutations on Vero cells. Confluent cell monolayers were
infected with the recombinant viruses containing wild-type UL20 (A, G),
or the UL20 mutants CL38 (B, H), CL41 (C, I), CL46 (D, J), CL49 (E, K),
or 216t (F, L), and viral plaques were visualized by immunohistochemistry
at 24 hpi...............................................................................................................188
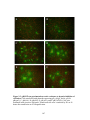
Figure 4.6: Domains of UL20p associated with UL20p-induced cell fusion.
Plaque phenotypes of recombinant viruses containing selected UL20
mutations are shown on Vero (A1, B1, C1, D1, E1, F1, G1, H1, I1, J1)
and G5 (A2, B2, C2, D2, E2, F2, G2, H2, I2, J2) cells. Confluent cell
monolayers were infected with recombinant viruses containing wild-type
UL20 (A1, A2), or the UL20 mutants CL49 (B1, B2), Y49A (C1, C2),
S50A (D1, D2), R51A (E1, E2), 216t (F1, F2), CL209 (G1, G2), R209A
(H1, H2), T212A (I1, I2), or R213A (J1, J2) at an MOI of 0.001, and viral
plaques were visualized at 30 hpi. White arrows indicate syncytia
formation .............................................................................................................190
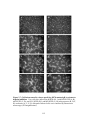
Figure 4.7: Electron micrographs of Vero cells infected with ∆20DIV5 (A),
∆20-rescue (B) or 216t (C, D, E, F) viruses. Confluent cell monolayers
were infected at an MOI of 5, incubated at 37°C for 24 hours, and
prepared for transmission electron microscopy. Panel D shows a higher
magnification of panel C. Panel F shows a partially-enveloped capsid
xiv
often seen in UL20-null infected cells. Nuclear (n), cytoplasmic (c) and
extracellular (e) spaces are marked .....................................................................192
Figure 4.8: Electron micrographs of Vero cells infected with the CL49 (A, B),
Y49Α (C), S50A (D) or R51A (E) viruses. Confluent cell monolayers
were infected at an MOI of 5, incubated at 37°C for 24 hours, and
prepared for transmission electron microscopy. Nuclear (n), cytoplasmic
(c) and extracellular (e) spaces are marked.........................................................194
Figure 5.1: gK localization at the TGN mediated by selected UL20p
mutants. Vero cells were co-transfected with gKD1V5 as well as with
plasmids encoding wild-type or mutant UL20p. 36 hours posttransfection, cells were washed thoroughly, fixed, and processed for
confocal microscopy. After permeabilization, rabbit αFLAG pAb was
used to identify UL20, and sheep αTGN46 pAb was used to identify the
TGN.....................................................................................................................214
Figure 5.2: gK cell surface expression and endocytosis to the TGN mediated
by selected UL20p mutants. Vero cells were co-transfected with
gKD1V5 as well as with plasmids encoding wild-type or mutant UL20p.
24 hours post-transfection, cells were incubated under live conditions with
aV5 (gK) mAb for 6 hours. Cells were washed thoroughly, fixed, and
processed for confocal microscopy. After permeabilization, rabbit
αFLAG pAb was used to identify UL20, and sheep αTGN46 pAb was
used to identify the TGN.....................................................................................216
Figure 5.3: Western analysis and potential phosphorylation of UL20p. (A)
Vero cells were either untransfected or transfected with a plasmid
specifying UL20p with an amino terminal 3xFLAG epitope tag prior to
infection with a UL20-null virus. Western analysis of UL20p was
performed utilizing a FLAG mAb. (B, C) Vero cells were infected with
either a UL20-null or a wild-type virus that encodes UL20p with an amino
terminal 3xFLAG epitope tag at an MOI of 10. 11 hours post infection,
the media was replaced and cells were incubated with phosphate-free
media for 1 hour. Phosphate-free media containing 32P (orthophosphate,
250 µCi/mL) was then added for 6 hours before cell lysates were prepared
and immunoprecipitation of UL20 was performed. The 32P profiles of the
total lysate (B) and the UL20 immunoprecipitations (C) are shown above.
Asterisks are provided to show corresponding bands representing UL20p........220
Figure 5.4: The proposed Alphaherpesvirus Fuseosome. A schematic
representation of interactions between the core integral membrane proteins
responsible for regulation of HSV-1 virus-induced cell-to-cell fusion is
shown. UL20p is thought to interact directly with both gB and gK (blue
arrows). There are also thought to be functional interactions that must
xv
take place between gB and gD or gH/gL (green arrow) in order to activate
the fusogenic potential of gB. .............................................................................226
xvi
ABSTRACT
The Herpes Simplex Virus life cycle contains a number of membrane fusion
events that must function properly to ensure a productive infection, including: virus
attachment and entry into susceptible cells, de-envelopment at the outer nuclear lamellae,
and virus-induced cell-to-cell fusion. A virus-free cell fusion assay was recently
developed in order to attempt to understand the underlying mechanisms that are
responsible for viral fusion events and was utilized in order to investigate the effect of
mutations targeted to the carboxyl terminus of gB. We showed that the predicted
α−helical domain H17b within the carboxyl-terminus of gB is involved in both virusinduced and virus-free fusion systems, and heparin was shown to be a specific inhibitor
of gB-mediated fusion in both systems, while resistance to heparin inhibition of gBmediated cell fusion was associated with the predicted α−helical structure H17b. An
important difference between virus-free and virus-induced membrane fusion is that virusexpressed gB mediates an insignificant amount of cell-to-cell fusion, while transiently
expressed gB causes extensive cell-to-cell fusion. gK was recently shown to inhibit cell
fusion resulting from transiently expressed gB, gD, gH and gL and hypothesized to
function as a negative regulator of membrane fusion. However, we show that gK can not
solely act as a negative regulator of gB-mediated membrane fusion, since gK is
demonstrated to be absolutely required for virus-induced cell fusion to occur, suggesting
a more complicated relationship between gK and gB. A recent publication from our
laboratory showed that syncytial mutations in either gB or gK failed to cause fusion in
the absence of the UL20 gene, suggesting that the UL20 protein was essential for virus-
xvii
induced cell fusion. Absence of the UL20 gene also caused accumulation of unenveloped
capsids into the cytoplasm, indicating that UL20p functioned in cytoplasmic stages of
virion envelopment. We delineated via site-directed mutagenesis the functional domains
of UL20p involved in infectious virus production and virus-induced cell fusion, revealing
that the role of UL20p in virus-induced cell fusion can be functionally separated from its
role in cytoplasmic virion morphogenesis.
xviii
CHAPTER I
INTRODUCTION
STATEMENT OF PROBLEM AND HYPOTHESIS
Herpes Simplex Viruses are known to be the etiologic agents responsible for
many human diseases including mucocutaneous oral and genital lesions,
keratoconjunctivitis, and viral encephalitis. During the herpesvirus life cycle there are a
number of membrane fusion events that are required for productive infection, including
those occurring during virus entry and egress from infected cells. In addition, certain
viral mutants exhibit the ability to cause extensive cell-to-cell fusion. The virus-induced
cell-to-cell fusion phenomenon has the potential to provide insight into the mechanisms
involved in all herpesvirus membrane fusion events. While the components involved in
virus entry are relatively well understood, the mechanisms involved in regulating
membrane fusion during virion egress and virus-induced cell-to-cell fusion are not well
defined.
During virion egress, there are multiple steps where the capsid must traverse or
acquire cellular membranes. HSV-1 is thought to acquire an initial envelope when
mature capsids formed in the nucleus of cells bud into the perinuclear space.
Subsequently, a putative fusion event between viral envelopes and the outer nuclear
membrane, termed de-envelopment, results in capsid release into the cytoplasm. Final
envelopment is thought to occur when capsids bud into cytoplasmic vesicles derived from
the TGN or endosomes, which facilitate egress of enveloped virions to extracellular
spaces (Mettenleiter, 2002b). Currently, it is not known whether viral glycoproteins
1
involved in virus entry or virus-induced cell fusion also function in the de-envelopment
step at the outer nuclear membrane. In addition, the mechanisms involved in final
envelopment are unclear.
Syncytia formation resulting from virus-induced cell-to-cell fusion can be viewed
as an aberrant manifestation of the interactions of altered membranes in herpesvirus
infected cells with the unaltered membranes of neighboring cells. Genetic analysis has
shown that mutations that cause extensive virus-induced cell-to-cell fusion map to at least
four and possible more loci within the viral genome: the UL20 gene (Baines et al., 1991;
MacLean et al., 1991; Melancon, Foster, and Kousoulas, 2004), the UL24 gene (Jacobson
et al., 1998; Sanders, Wilkie, and Davison, 1982), the UL27 gene encoding glycoprotein
B (gB) (Bzik et al., 1984; Pellett et al., 1985), and the UL53 gene coding for glycoprotein
K (gK) (Bond and Person, 1984; Debroy, Pederson, and Person, 1985; Hutchinson et al.,
1992b; Pogue-Geile et al., 1984; Pogue-Geile and Spear, 1987; Ruyechan et al., 1979),
with the great majority of syncytial mutations arising in either UL27 (gB) or UL53 (gK).
Virus-induced cell-to-cell fusion has been extensively studied for several reasons:
as a probe of the structure and function of cellular membranes during infection reflected
in their behavior, as a tool for analysis of the functions of viral membrane proteins, and as
a model of the initial interaction between HSV and susceptible cells that results in fusion
of the viral envelope with the cellular plasma membrane (Campadelli-Fiume and
Serafini-Cessi, 1985; Roizman, 1962; Spear, 1985b; Spear, 1993). Recently, it was
shown that transiently expressed gB, gD, gH, and gL are necessary and sufficient to
induce membrane fusion in a virus-free membrane fusion system, reflecting the core
requirements to activate the herpesvirus fusion machinery (Turner et al., 1998).
2
STATEMENT OF RESEARCH OBJECTIVES
The goal of this research was to investigate the structure and functions of the
Herpes Simplex Virus Type 1 UL20 protein (UL20p), glycoprotein B (gB), and
glycoprotein K (gK) in virion morphogenesis and egress, and virus-induced cell fusion
The specific aims of this research were:
I.
To analyze the features in the carboxyl terminus of gB required for the regulation
of syncytia formation in a virus-free fusion assay:
1.
To delineate gB carboxyl terminal domains involved in cell-to-cell fusion
by generating a panel of site specific and truncation mutations in the
carboxyl-terminus of gB and determine their effect on cell-to-fusion
resulting from transiently expressed gB, gD, and the gH/gL heterodimer.
2.
To determine whether soluble heparin, a specific inhibitor of gB-mediated
virus-induced cell fusion, can inhibit gB-mediated cell fusion in the
transient co-expression system (virus-free) in the presence or absence of
gB-carboxyl terminal mutations that alter gB’s fusogenic properties.
3.
To facilitate a determination of the relevance of the results obtained in the
virus-free fusion system by comparison to the effects of similar gB
mutations in virus-induced cell-to-cell fusion.
II.
To analyze whether gB, gK, and UL20p function redundantly in virion deenvelopment from the perinuclear space during virion morphogenesis and egress:
1.
To generate UL20-null, gB-null, gK-null, UL20/gB double-null, and
gK/gB double-null viruses through the use of a bacterial artificial
chromosome (BAC) containing the entire HSV-1 genome.
3
2.
To characterize the ultrastructural phenotypes of the UL20/gB double-null
and gK/gB double-null viruses.
III.
To investigate the requirement of gK or UL20p in virus-induced cell fusion
caused by syncytial mutations and truncations in the carboxyl terminus of gB:
1.
To introduce a gBsyn3 and gBamb1511 mutation into the previously
constructed UL20/gB double-null and gK/gB double-null viruses.
2.
To characterize the syncytial phenotypes of the resulting UL20-null and
gK-null viruses containing syncytial gB mutations.
IV.
To delineate the functional domains of UL20p involved in cytoplasmic virion
envelopment and virus-induced cell fusion:
1.
To generate a panel of single and multiple (cluster) alanine substitutions as
well as carboxyl-terminal truncations targeting specific areas of interest
and conserved regions of the UL20 protein.
2.
To analyze the ability of each mutant UL20p gene to complement the
UL20-null defect for infectious virion production and virus-induced cell
fusion caused by gB and gK syncytial mutations.
3.
To compare complementation for virus replication results with the
phenotypes of recombinant viruses containing mutants of interest.
4.
To investigate the ultrastructural phenotypes of viruses specifying mutant
UL20 genes exhibiting defects in virion egress and spread.
Overall, the results obtained from this research indicate that: the carboxyl
terminus of gB contains specific domains that regulate gB-mediated membrane fusion
phenomena; UL20p, gB, and gK are interdependent for virus-induced cell-to-cell fusion;
4
and specific domains of UL20p function in cell fusion and virion egress. The work is
presented in individual chapters in a manuscript format having a specific title for the
central theme of each chapter:
Chapter II:
An alpha-helical domain within the carboxyl terminus of herpes simplex
virus type 1 (HSV-1) glycoprotein B (gB) is associated with cell fusion
and resistance to heparin inhibition of cell fusion.
Chapter III:
Herpes Simplex Virus Type 1 (HSV-1) Glycoprotein K (gK) is Required
for Glycoprotein B (gB)-mediated Virus-Induced Cell Fusion, while
neither gB and gK nor gB and UL20p Function Redundantly in Virion DeEnvelopment
Chapter IV:
Genetic analysis of the herpes simplex virus type 1 UL20 protein domains
involved in cytoplasmic virion envelopment and virus-induced cell fusion.
LITERATURE REVIEW
Historical Perspective of Herpesviruses
The first description of Herpes Simplex Virus (HSV) can be traced back to the
ancient Greeks. Hippocrates used the term “herpes” to describe lesions that appeared to
creep or crawl along the skin (Wildy, 1973). Descriptions of lesions resembling HSV
were also found on a Sumerian Tablet dated to the 3rd Millenium BC and the Ebers
Papyrus, circa 1500 BC (Whitley, 2001). Herodotus is noted as the first to describe an
association between the cutaneous lesions and fever caused by HSV, and Galen
recognized that recurrent HSV lesions develop at the same anatomical location (Whitley,
2001). During the 18th century, Bateman accurately described the nature of HSV
infection as a “restricted group of localized vesicles with a short, self-limiting course”
5
(Bateman, 1814). The first description of the link between HSV and the genital organs
did not appear until De Morbis Venereis was published by John Astruc, physician for
King Louis XIV, in 1736, after studying the afflictions of French prostitutes (Astruc,
1736).
During the late 19th and early 20th century, human volunteers were often used to
test the transmission of infectious agents, and Vidal showed that HSV was infectious by
passing it from human to another (Vidal, 1873). Gruter, in a switch from human to
animal studies, demonstrated that HSV could be transmitted from rabbit to rabbit, and he
is widely credited with the isolation of HSV by the virology community (Gruter, 1924).
In 1939, Burnett and Williams published an article describing the nature of latency,
noting that HSV seems to persist for life and can be reactivated under stressful conditions
to produce visible lesions (Burnet and Williams, 1939).
The development of tissue culture technology was critical in the isolation of other
members of the human herpesvirus family. Between 1952 and 1956, varicella zoster
virus (VZV), the causative agent of chicken pox, and cytomegalovirus (CMV) were
isolated (Craig et al., 1957; Rowe et al., 1956; Smith, 1956; Weller and Stoddard, 1952).
The eventual cultivation of lymphoblastoid tumor cells and B lymphocytes led to the
isolation and study of Epstein-Barr virus (EBV) (Epstein, Achong, and Barr, 1964). In
the 1990s, cultivation of T lymphocytes led to the isolation of human herpesviruses 6A,
6B, and 7 (Frenkel et al., 1990; Lopez et al., 1988; Salahuddin et al., 1986). More
recently, Representational Differential Analysis (RDA) led to the discovery of human
herpesvirus 8 (Chang et al., 1994).
6
Taxonomy of Herpesviridae
Identification of the new and apparently related viruses led to a scientific desire
for classification. However, it was not until 1981 that the current herpesvirus
classification came into being. All herpesviruses examined to date are capable of
establishing a latent infection in their natural hosts in a specific set of cells, which varies
from one virus to another. Other biological properties vary, such as the length of the
reproductive cycle, and these were used as the basis of classification, before DNA
sequences of the viruses were known. Members of the family Herpesviridae were
initially classified by the Herpesvirus Study Group into three subfamilies: the
Alphaherpesvirinae, the Betaherpesvirinae, and the Gammaherpesvirinae (Roizman,
Bartha, and Biggs, 1973; Roizman et al., 1992; Van Regenmortel et al., 2000). DNA
sequence data has since supported and expanded the platform on which the classification
system rests. Table 1.1 shows the classification of the nine known human herpesviruses
(bold type) as well as other commonly studied herpesviruses (Roizman et al., 1992; Van
Regenmortel et al., 2000).
Alphaherpesvirinae were classified based on their variable host range, short
reproductive cycle, rapid spread in tissue culture, efficient destruction of infected cells,
and the ability to establish latent infections primarily in sensory ganglia. The subfamily
consists of the genera Simplexvirus, Varicellovirus, Marek’s disease-like virus, and
Infectious laryngotracheitis-like virus (Van Regenmortel et al., 2000).
Betaherpesvirinae were characterized by a limited host range, long reproductive
cycle, and slow infection progression in tissue culture. Cells that are infected often
become enlarged (cytomegalia), and the viruses can maintain latency in secretory
7
Table 1.1: Members of the family herpesviridae
Subfamily
Alphaherpesvirinae
Designation
Vernacular Name
Human herpesvirus 1 (HHV-1)
Human herpesvirus 2 (HHV-2)
Human herpesvirus 3 (HHV-3)
Cercopithecine herpesvirus 1 (CeHV-1)
Gallid herpesvirus 1 (GaHV-1)
Gallid herpesvirus 2 (GaHV-2)
Suid herpesvirus 1 (SuHV-1)
Felid herpesvirus 1 (FeHV-1)
Ictalurid herpesvirus 1 (IcHV-1)
Betaherpesvirinae
Gammaherpesvirinae
Human herpesvirus 5 (HHV-5)
Cercopithecine herpesvirus 8 (CeHV-8)
Murid herpesvirus 1 (MuHV-1)
Murid herpesvirus 2 (MuHV-2)
Suid herpesvirus 2 (SuHV-1)
Felid herpesvirus 2 (FeHV-1)
Human herpesvirus 6A (HHV-6A)
Human herpesvirus 6B (HHV-6B)
Human herpesvirus 7 (HHV-7)
Human herpesvirus 4 (HHV-4)
Human herpesvirus 8 (HHV-8)
Herpes simplex virus type 1 (HSV-1)
Herpes simplex virus type 2 (HSV-2)
Varicella-zoster virus (VZV)
Herpesvirus B, Simian Herpesvirus
Infectious laryngotracheitis virus
Marek's disease herpesvirus 2
Pseudorabies virus, Aujesky's disease
Feline herpesvirus 1, Feline rhinotracheitis
herpesvirus
Channel catfish herpesvirus
Cytomegalovirus (CMV)
Rhesus monkey cytomegalovirus
Mouse cytomegalovirus
Rat cytomegalovirus
Pig cytomegalovirus
Cat cytomegalovirus
Roseolovirus
Epstein-Barr virus (EBV)
Karposi's sarcoma-associated
herpesvirus (KSHV)
glands, lymphoreticular cells, kidneys, and other tissues. The subfamily consists of the
genera Cytomegalovirus, Muromegalovirus, and Roseolovirus (Van Regenmortel et al.,
2000).
Gammaherpesvirinae were classified by a limited host range and ability to
replicate in lymphoblastoid cells, with some viruses also causing lytic infection in some
types of epithelial and fibroblastic cells. Viruses are usually specific for either B or T
lymphocytes, and latent virus is frequently demonstrated in lymphoid tissue. The
8
Gammaherpesvirinae subfamily consists of the genera Lymphocryptovirus (EBV), and
Rhadinovirus (Van Regenmortel et al., 2000).
Clinical Significance of Herpes Simplex Viruses
Epidemiology
Infections caused by HSV occur worldwide in both developed countries and
underdeveloped countries (Black, 1975). There are no known animal carriers for HSV;
therefore, humans remain solely responsible for transmitting virus to other humans. Virus
transmission from an infected to a susceptible individual occurs during close personal
contact. The frequency of person to person contact appears to be the major mediator of
infections (Whitley, 2001). Due to HSV infection rarely resulting in fatality and the
nature of latency, more than half of the world’s population probably have recurrent HSV
infections, enabling the transmission of HSV. Initial HSV infection usually occurs in
children less than 5 years old and is most often asymptomatic. The mouth area is the
most common location of infection (Whitley, 2001). Primary infection leads to the
shedding of virus from the mouth and stool for an average of 7 to 10 days (Amir et al.,
1997), and neutralizing antibodies appear between 4 and 7 days after the onset of HSV
infection and peak at approximately 3 weeks post infection (Buddingh et al., 1953).
The primary factors affecting the rate of HSV infection are location,
socioeconomic status, and age. In some studies, by the age of 15, over 95% of children
were found to possess antibodies to HSV (Bader et al., 1978; Black et al., 1974). HSV-1
seroprevalence is much more common than its counterpart HSV-2 (Fleming et al., 1997;
Nahmias, Lee, and Beckman-Nahmias, 1990). Because HSV-2 is usually acquired
through sexual contact, antibodies to HSV-2 are rarely found before the onset of sexual
9
activity. While most genital infections are caused by HSV-2, there is an ever-increasing
proportion attributable to HSV-1 (Corey et al., 1983; Kalinyak, Fleagle, and Docherty,
1977). Genital HSV-1 infections are usually both less severe than HSV-2 and less prone
to recurrence (Corey et al., 1983; Reeves et al., 1981).
Pathogenesis
Initially, virus must come in contact with a mucosal surface or abraded skin for
HSV infection to be initiated. After primary infection, viral replication at the infected
location, usually oral or genital mucosal tissue, results in the infection of sensory nerve
endings; and virus is then transported to the dorsal root ganglia (Baringer and Swoveland,
1973; Bastian et al., 1972). In HSV-1 infection, the trigeminal ganglia becomes
colonized and harbors latent virus; whereas in HSV-2 infection the sacral ganglia is the
site of latency (Whitley, 2001). After the establishment of latency, certain stimuli can
cause reactivation to occur, and virus becomes evident at mucocutaneous sites as vesicles
or ulcers. A more severe primary infection can result in a higher rate of HSV
reactivation.
Cellular changes induced by viral infection include enlargement of infected cells
and the appearance of condensed chromatin within the nuclei, followed by degradation of
the nuclei. Cells lose intact plasma membranes and form multinucleated giant cells. In
infected dermal regions, there is an intense inflammatory response, and the intensity
decreases substantially with recurrent disease (Whitley, 2001).
Mucocutaneous Infections
Primary HSV-1 infection can be either totally asymptomatic or can result in a
symptoms including fever, sore throat, vesicular or ulcerative lesions. However,
10
asymptomatic infection is generally the rule rather than the exception (Whitley, 2001).
The duration of symptomatic disease in children is generally 2 to 3 weeks, with a fever of
101°F to 104°F. The onset of a recurrent HSV-1 infection is generally marked pain,
burning, tingling, or itching, which generally lasts for less than 6 hours, and is followed
by vesicle formation within 24 to 48 hours (Spruance and Crumpacker, 1982; Spruance et
al., 1984; Spruance et al., 1977). Usually, 3 to 5 vesicles appear at the border of the lip
and last no longer than 48 hours. Pain is most severe at the outset of vesicle formation
and resolves in 4 to 5 days. The frequency of recurrences varies greatly among
individuals, and the factors that influence reactivation are poorly defined but may include
fever, stress, and exposure to UV light (Segal et al., 1974; Ship, Miller, and Ram, 1977).
In HSV-2, the most severe clinical symptoms are encountered with primary
infection, characterized by the appearance of macules and papules followed by vesicles,
pustules, and ulcers. The duration of lesions and viral secretion averages about 3 weeks.
Men and women experience both similar and dissimilar symptoms (Corey, 1982; Corey
et al., 1983). Pre-existing immunity to HSV-1 can have a beneficial effect in reducing
the severity of HSV-2 primary infections (Allen and Rapp, 1982; Corey et al., 1981;
Kaufman et al., 1973). Recurrent HSV-2 is milder than initial infection and is
characterized by the appearance of 3 to 5 vesicles (Adams et al., 1976). Symptoms
usually last 7 to 10 days, and virus is shed for an average of 2 to 5 days. The biggest
problem involving recurrent genital herpes is the frequency of recurrences, which varies
by individual. Recurrences usually occur several times per year; and, whether
symptomatic or asymptomatic, transmission of the infection to sexual partners can occur
with intimate contact (Corey et al., 1983).
11
Fetal and Neonatal Infections
Neonatal HSV infections occur at a rate of about 1 in 3000 deliveries per year
(Nahmias, Keyserling, and Kerrick, 1983; Nahmias, Keyserling, and Lee, 1989), and
infection occurs far less frequently than genital infections in the adult population. The
type of maternal genital infection at the time of delivery is directly tied to the risk of fetal
infection. If the mother has a primary infection the risk of transmission is approximately
30%, while the risk for recurrent infection is 3% or less (Brown et al., 1991). The most
common route of infection, 75% to 80%, is intrapartum contact of the fetus with infected
maternal secretions (Whitley, 2001). The clinical symptoms of neonatal HSV infection
are a reflection of the site and extent of viral replication, with infection being almost
always symptomatic and often lethal. Babies with HSV infection can have disease:
localized to the skin, eye, and mouth; encephalitis with or without skin involvement; or
disseminated infection involving multiple organs (Nahmias et al., 1970; Whitley et al.,
1981). The highest mortality rate occurs in babies with disseminated infection. In
addition, frequently occurring HSV-2 cutaneous lesions defined a group at risk for
neurological problems (Whitley et al., 1991).
Keratoconjunctivitis
There are 300,000 new cases of HSV eye infection annually, ranking second
behind trauma as the cause of corneal blindness (Binder, 1977). Primary herpetic
keratoconjunctivitis can occur in either a single eye or both eyes, and healing of the
cornea can take as long as one month even with antiviral therapy. Recurrent HSV eye
infections occur at a similar rate to HSV-1 mucocutaneous infections and most often
12
involve only a single eye. Repeated attacks can last for weeks or months and progressive
disease can result in vision loss (Whitley, 2001).
Infection of an Immunocompromised Host
Patients who are immunocompromised due to immunotherapy, malnutrition or
acquired immunodeficiency syndrome are at risk for severe HSV infections, and these
patients may develop progressive disease involving the respiratory tract, esophagus, or
the gastrointestinal tract (Korsager et al., 1975; Montgomerie et al., 1969). Recurrent
HSV infection can occur in theses patients at multiple sites and healing occurs over an
average of 6 weeks (Whitley et al., 1984). The repeated treatment required for these
patients can lead to viral mutants resistant to antiviral therapy.
Central Nervous System (CNS) Infections
HSV is the most common cause of sporadic, fatal encephalitis in this country
(Olson et al., 1967). Some studies estimate a rate as high as 1250 cases per year in the
United States (Whitley, 2001). Encephalitis is caused when the virus spreads past the
dorsal root ganglia, in which latency is usually established, to the CNS. The mechanisms
responsible for this aberrant event in the virus life cycle are unclear. The manifestations
of HSV encephalitis include primarily focal encephalitis along with fever, altered
behavior, and localized neurological findings. There is usually evidence of a localized
temporal lobe disease (Whitley et al., 1977; Whitley et al., 1981). In untreated patients,
mortality exceeds 70% and only 2.5% of patients return to normal neurological function
(Whitley, 2001).
13
Prevention and Treatment of HSV Infection
The two methods for control of HSV infections are antiviral therapy and
prevention. Antivirals such as acyclovir and valaciclovir are effective in limiting the
extent of HSV infection and therefore helpful in limiting spread to uninfected individuals.
However, post-exposure antiviral treatment does not prevent lifelong infection of an
individual. Prevention of HSV infection is mainly achieved through avoiding contact
with infectious secretions. Vaccination would be the ideal method of HSV prevention;
however, to date no HSV vaccine has been clinically successful.
Architecture of the Herpes Virion
Virions of herpes viruses can vary in size from 120 nm to 300 nm (Roizman
and Furlong, 1974), and consist of: an electron-dense core, an icosadeltahedral capsid
around the core, an amorphous tegument around the capsid, and an outer envelope
containing glycoprotein spikes (Roizman and Furlong, 1974). The variability in the size
of herpes virions is due mainly to variability in the makeup of the tegument and the state
of the envelope. A model of the virion architecture is presented below in Figure 1.1.
The Core
The core of a mature herpes virion contains the viral DNA in the form of a torus
that may appear to be suspended by a proteinaceous spindle to the capsid (Falke, Siegert,
and Vogell, 1959; Furlong, Swift, and Roizman, 1972; Nazerian, 1974). The toroidal
structure is 50 nm high, with an inside diameter of 18 nm and an outside diameter of 70
nm. The arrangement of the viral DNA in the torus is not known.
14
Figure 1.1: Herpesvirus virion structure. Virions of herpes viruses can vary in size
from 120 nm to 300 nm (Roizman and Furlong, 1974). A virion consists of: an electrondense core containing the viral genome, an icosadeltahedral capsid around the core, an
amorphous tegument around the capsid, and an envelope derived from cellular
membranes containing glycoprotein spikes (Roizman and Furlong, 1974).
15
The Capsid
The capsid is approximately 125 nm in diameter in the form of an
icosadeltahedron and features 162 capsomeres, characteristic of all herpesviruses.
Capsomeres are either pentons or hexons, consisting of 5 or 6 VP5 (major capsid protein)
monomers, respectively. In the capsid, the pentons are located on the icosahedral 5-fold
vertices, and the hexons make up the faces and edges. Hexons also contain 6 copies of
VP26, attachedto the upper edge of VP5 and form a continuous ring around each hexon
(Zhou et al., 1995). A heterotrimeric complex known as the triplex connects the
capsomeres; the triplex consists of two copies of VP23 and one copy of VP19C and acts
as a sort of scaffold for the capsid (Spencer et al., 1998).
The Tegument
The tegument is contained between the capsid and the virion envelope and
appears fibrous on negative staining (Morgan et al., 1959; Morgan, Rose, and Mednis,
1968; Wildy and Watson, 1962). The tegument can be distributed asymmetrically and its
thickness can vary depending on the location of the virion particle within the infected
cell. There is less tegument that is more symmetrically arranged in perinuclear virions
than in virions in cytoplasmic vesicles that contain more tegument distributed more
asymmetrically (Falke, Siegert, and Vogell, 1959). Tegument proteins are important in
various aspects of the virus life cycle and are believed to have key functions in the early
events of infection and virion egress. There is ordered tegument density around the
pentons, suggesting symmetry where the capsid and tegument interact (Zhou et al., 1999).
This density may be due to the VP1-3 protein, an extremely large 336 kDa protein,
thought to be involved in nucleocapsid attachment to the nuclear pore facilitating DNA
16
release into the nucleoplasm (Batterson, Furlong, and Roizman, 1983; Knipe et al., 1981;
Ojala et al., 2000). However, VP1-3 null mutants also accumulate newly assembled,
DNA-filled capsids in the cytoplasm of infected cells, indicating that VP1-3 is involved
in various stages of the virus life cycle (Desai, 2000).
The Envelope
The outer covering of the herpesvirus, the envelope, has a typical trilaminar
appearance (Epstein, 1962) and appears to be made up of altered cellular membranes
(Armstrong, Pereira, and Andrewes, 1961; Falke, Siegert, and Vogell, 1959; Morgan,
Rose, and Mednis, 1968). The herpesvirus envelope contains numerous glycoprotein
extrusions, while the amounts of each glycoprotein vary. HSV specifies at least 11
different glycoproteins, and the copy number of each glycoprotein can well exceed 1,000
per virion. Envelope glycoproteins gB, gD, gH, and gL have been shown to be required
for virion entry into susceptible cells.
Organization of the Viral Genome
The viral DNA of herpesviruses is linear and double stranded, but the DNA
becomes circular immediately after release from capsids into the nucleoplasm of the
infected cells. The length of the genome of different herpesviruses varies between 120 to
250 kbp, with the size of HSV-1 determined to be 152,261 bp (McGeoch et al., 1988).
This variability is different than polymorphism in the genome length of individual
viruses, which is due to terminal and internal repeated sequences that can vary in copy
number, leading to variations in genome length of more than 10 kbp. The total G+C
content of herpesviruses varies from 31% to 75%, and this percentage can vary across the
genome (Roizman and Pellett, 2001). HSV-1 and HSV-2 contain approximately 68%
17
and 69% G+C content, respectively (Becker, Dym, and Sarov, 1968; Kieff,
Bachenheimer, and Roizman, 1971).
The sequence arrangement of herpesvirus genomes varies on the presence and
location of reiterated sequences that allow rearrangement to occur. In Herpes Simplex
Virus genomes, the sequences from both termini are repeated in an inverted orientation
and juxtaposed internally. As a result, the genome is divided into two regions, consisting
of the unique long (UL) and unique short (US) regions flanked by inverted repeats (Figure
1.2)
UL
TR
IRL IRS
A
B
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
US
TR
0.9
1.0
Figure 1.2. Arrangement of the HSV-1 genome. (A) The top line represents
the prototypic arrangement of the HSV-1 genome with the unique long (UL) and
unique short (US) regions flanked by the terminal repeat (TR) and internal repeat
(IR) regions. (B) The bottom line shows map units of the HSV-1 genome.
Both components are able to invert relative to the other to form four isomers; and, as
predicted, DNA purified from infected cells contains four equivalent populations, which
differ in the relative orientation of the unique long and unique short regions (Roizman
and Pellett, 2001).
The majority of herpesvirus genes contain: a promoter region 50 to 200 bp
upstream of a TATA box, a transcription initiation site 20 to 25 bp downstream of the
TATA box, a 5’ untranslated leader sequence of 30 to 300 bp, a single major open
reading frame (ORF) with a translation initiation codon meeting the host requirement for
18
efficient initiation, 10 to 30 bp of 3’ untranslated sequence, and a polyadenylation signal
with standard flanking sequences (Roizman and Pellett, 2001). Some exceptions include
genes without a TATA box or genes with a second in-frame initiator methionine (Chou
and Roizman, 1986; Markovitz, Filatov, and Roizman, 1999). Most transcriptional gene
products are not spliced, although every herpesvirus expresses a few spliced genes.
Herpesviruses also produce noncoding RNAs, such as the HSV-1 latency associated
transcript (LAT) (Roizman and Pellett, 2001). The different members of the herpesvirus
family encode between 70 and 200 genes, estimated using various methods (Roizman and
Pellett, 2001). HSV-1 encodes about 90 gene products, with at least 84 of the
transcriptional units encoding proteins (Roizman and Knipe, 2001).
The Herpes Simplex Virus Lifecycle
Virus Attachment and Entry
Herpesvirus entry is a multistep process involving multiple viral glycoproteins
acting as ligands for multiple receptors on the surface of the target cell. Entry is the most
critical step in the HSV life cycle and greatly determines the tropism and pathology of
each member of the herpesvirus family. The wide host range of HSV and narrow host
range of EBV can be in part explained by the ability of each virus to utilize a different
array of cell surface binding and entry receptors. Entry of HSV occurs is three distinct
stages: the first step involves virus binding to the surface of the cell, the second step
involves an interaction of gD (HSV-1) with an entry receptor, and the third step involves
fusion of the viral envelope with the plasma membrane of the cell, releasing the capsidtegument complex into the cytoplasm of the infected cell (Figure 1.4).
19
Herpes Simplex Virus Life Cycle
α
β
γ
Figure 1.3: The Herpes Simplex Virus Life Cycle. The first stage of the herpes virus
life cycle consists of virus entry, capsid transport to the cell nucleus, deposition of viral
DNA into the nucleoplasm, coordinate gene expression and viral DNA replication (black
arrows). The second stage is virion morphogenesis and egress comprised of primary
envelopment at the inner nuclear membrane, de-envelopment at the outer nuclear
membrane, final envelopment into cytoplasmic vesicles and transport to extrancellular
spaces (red arrows).
20
HSV-1 Viral Entry
gH/gL
gD
“Ligand for Fusion”
HVEM,
nectins
gC
“Receptor for Fusion”
“Ligand for Binding”
gB
Heparan
Sulfate
CYTOPLASM
“Receptor for Binding”
NUCLEUS
Figure 1.4: Herpes Simplex Virus Entry. The first step of HSV entry is attachment to
the plasma membrane of infected cells through interaction of gC (ligand for binding)
with cell surface heparan sulfate (receptor for binding). The second step is fusion of the
viral envelope with the cell plasma membrane and occurs following attachment of gD
(ligand for fusion) to either HVEM (herpesvirus entry mediator) or a nectin (receptor for
fusion) on cell surfaces. Fusion requires the presence of gD and an entry receptor, as
well as gB and the gH/gL heterodimer.
21
Binding Receptors
Heparan Sulfate (Glycosaminoglycans)
Initial contact of herpesviruses with cells usually occurs through binding
receptors, in which engagement of the virus with receptor is reversible and changes in the
virion envelope required for fusion and entry do not occur. Herpes Simplex viruses
initial attachment to cells involves the interaction of glycoprotein C, and glycoprotein B
to a lesser extent, with cell surface glycosaminoglycans, usually heparan sulfate (Shieh et
al., 1992; WuDunn and Spear, 1989). Heparan sulfate is synthesized as the
glycosaminoglycan component of heparan sulfate proteoglycans, and it is widely
distributed on cell surfaces in mammals. The presence of heparan sulfate is not essential
for virus entry, but it greatly increases the efficiency of virus entry into cells (Banfield et
al., 1995). Similarly, although glycoprotein C confers the greatest efficiency for virus
attachment to cell surfaces, increasing the efficiency of virus binding by approximately
10 fold, it is not essential for either virus entry or replication (Heine et al., 1974).
Entry Receptors
Binding of the HSV glycoprotein D to an entry receptor sets in motion an
irreversible chain of events leading to the fusion of the virion envelope with the plasma
membrane of the target cell. There have been three classes of HSV entry receptors
classified to date, indicated below and shown in Figure 1.5.
Tumor Necrosis Factor (TNF) Receptor Family
HVEM / Herpes Virus Entry Mediator A (HveA) is a member of the TNF
receptor family and is expressed in a variety of different cell types including
lymphocytes, other leukocytes, epithelial cells, and fibroblasts. While HVEM is utilized
22
HSV-1 Cell-Surface Entry Receptors
Tumor necrosis factor
(TNF) receptor family
HVEM
(HveA)
3-O-sulfated
heparan sulfate
Immunoglobulin Superfamily
V
V
C2
C2
C1
C1
Nectin-1α
(HveC)
Nectin-1β
(HIgR)
V
V
V
C2
C2
C2
C1
C1
C1
Nectin-2α
(HveB)
Nectin-2δ
Nectin-3
3-O-sulfated
heparan
sulfate
Figure 1.5: Herpes Simplex Virus entry receptors. The three classes of cell surface
receptors for HSV entry are: the tumor necrosis factor (TNF) receptor family consisting
of HVEM, the immunoglobulin superfamily consisting of the nectins, and 3-O-sulfated
heparin sulfate. Only viral attachment can occur in the absence of an HSV entry
receptor.
23
by herpes simplex viruses as an entry receptor, the natural ligands for HVEM include
LIGHT and lymphotoxin-alpha (Mauri et al., 1998). LIGHT can function as a second
signal for T-cell activation, and the interactions between LIGHT and HVEM are the
focus of active investigations of the regulation of immune responses (Kwon et al., 2003).
HVEM serves as an excellent entry mediator for both HSV-1 and HSV-2. Certain studies
have shown that HSV-1 entry of activated T-cells is principally mediated by HVEM,
although in a variety of other cell types HVEM was shown not to be the primary mediator
of virus entry (Montgomery et al., 1996).
Immunoglobulin Superfamily
The second family of herpes simplex virus entry receptors is part of the
immunoglobulin superfamily and includes several isoforms present in both human and
nonhuman cells encoded by mRNAs that can vary due to alternative splicing. Members
of this family have been shown to act as intercellular adhesion molecules localized at
adhesion junctions, in which the carboxyl-terminal domains bind to L-afadin, a PDZbinding protein that anchors the receptors to the cytoskeleton and adherens junctions.
These receptors have been named “nectins” on account of their cellular function
(Takahashi et al., 1999). Nectins are highly conserved among mammalian species in
respect to structure, function, and the ability to mediate HSV entry (Milne et al., 2001;
Shukla et al., 2000). The nectins are expressed in a variety of cell types including
epithelial cells, fibroblasts, and neurons (Takai and Nakanishi, 2003). In addition, the
distribution of this class of receptors in human tissues reflects the susceptibility of cells to
infection and probably accounts for both virus entry and cell-to-cell spread (Roizman and
Knipe, 2001).
24
Nectin-1α (Herpes Virus Entry Mediator C, HveC) and nectin-1β (Herpesvirus
Immunoglobulin-like receptor, HIgR) are two mRNA splicing variants containing a
common ectodomain. They are expressed on epithelial, fibroblastic, neural and
hematopoietic cells, in keratinocytes, and in human tissues that are the target of HSV
infection including skin, brain, and spinal ganglia (Cocchi et al., 1998; Geraghty et al.,
1998). Nectin-1α and nectin-1β have the ability to mediate entry of all HSV-1 strains,
HSV-2, Pseudorabies virus (PrV) and bovine herpes virus 1 (BHV-1) (Geraghty et al.,
1998).
Nectin-2α (Herpes Virus Entry Mediator B, HveB) and nectin-2δ are also mRNA
splice variants, and they mediate the entry of HSV-2, PrV, and certain viable mutant
forms of HSV-1 but not wild-type HSV-1 (Lopez et al., 2000; Warner et al., 1998).
Nectin-1 and nectin-2 are related to nectin-3 and nectin-4, and the poliovirus receptor
(Takai and Nakanishi, 2003).
Nectin-3 has been shown to be expressed on J cells, which lack HVEM, nectin-1,
and nectin-2. An HSV-1 mutant, designated HSV-1 (JMP), was able to enter into J cells
expressing nectin-3, but wild-type HSV-1 was not (Cocchi et al., 2004). HSV-1 (JMP)
contains mutations in glycoprotein K as well as glycoprotein D. The mutations in both
gK and gD were shown to be required for HSV-1 (JMP) entry into J cells (Cocchi et al.,
2004).
3-O-sulfated Heparan Sulfate (3-OS HS)
Modification of heparan sulfate by D-glucosaminyl 3-O-sulfotransferase
isoforms-3 and/or -5 creates 3-O-sulfated Heparan Sulfate (3-OS HS), generating a gDbinding site that allows heparan sulfate proteoglycans to function as entry receptors for
25
HSV-1. 3-O-sulfated heparan sulfates are broadly distributed on human cells and tissue
and mediate HSV-1 but not HSV-2 entry (Shukla et al., 1999).
Other Herpesvirus Receptors
Only alphaherpesviruses (except VZV) encode members of the gD family.
Betaherpesvirus and gammaherpesviruses rely on other viral proteins to serve as ligands
for cellular receptors. EBV uses gp42, which becomes a member of a gH-gL-gp42
complex, to serve as a ligand for human leukocyte antigen (HLA) class II molecules on B
lymphocytes. Human CMV encodes glycoprotein O (gO), which is unrelated to gp42 but
also forms a complex with gH-gL (Huber and Compton, 1998). In addition, HHV-6A
encodes glycoprotein Q (gQ), which forms a complex with gH-gL (Mori et al., 2003).
However, it is unclear whether or not gO or gQ are required for entry in CMV or HHV6A, respectively.
Virus-to-Cell Fusion
After binding of gD to a HSV-1 virus entry receptor, the last step in virus entry is
fusion of the virion envelope with the plasma membrane of the target cell (Morgan, Rose,
and Mednis, 1968). The current belief is that gD (Ligas and Johnson, 1988), gB
(Sarmiento, Haffey, and Spear, 1979), and the gH/gL heterodimer (Forrester et al., 1992)
are required for the virus-to-cell fusion process to occur. Since gD is required for entry
receptor binding, it would be logical to hypothesize that receptor binding triggers a
conformational change in gD that translates to a change in gB and gH/gL, resulting in
activation of the HSV-1 membrane fusion machinery. In some cells types, notably CHO
cells that express an entry receptor such as nectin-1 and HeLa cells, endocytosis and
acidification of endosomes are required for efficient virus entry. Interestingly, it was
26
found that endocytic uptake of HSV virions from the cell surface is rapid and
independent of any known gD receptor. In addition, efficient entry through endocytosis
requires cellular PI 3-kinase activity and the viral glycoproteins gB, gD, and gH/gL
(Nicola, McEvoy, and Straus, 2003; Nicola and Straus, 2004). While these two entry
pathways are spatially distinct, they are still functionally similar in that both require the
same set of viral glycoproteins and a gD entry receptor.
Host Protein Shutoff
HSV shuts off the synthesis of cellular protein products in two stages. First, the
virus mediates degredation of the host cell mRNA. Second, there is inhibition of further
synthesis and processing of host mRNA. The virion host shutoff (vhs) function involves
structural components of the virion and does not require de novo protein synthesis after
infection (Fenwick and Walker, 1978; Nishioka and Silverstein, 1977; Nishioka and
Silverstein, 1978a; Nishioka and Silverstein, 1978b). The isolation of vhs- mutants,
which failed to shutoff host cell protein synthesis in HSV-1 infected cells, allowed for
more detailed study of the viral requirements for host shutoff (Read and Frenkel, 1983).
The UL41 ORF was identified as being responsible for the phenotype in the vhs- mutants
(Fenwick, Morse, and Roizman, 1979; Kwong, Kruper, and Frenkel, 1988; Oroskar and
Read, 1987). The UL41 protein (VHS) is expressed as a γ1 gene (Frink, Anderson, and
Wagner, 1981) and is translated to a 58,000 or 59,500 kDa phosphoprotein (Read, Karr,
and Knight, 1993). In addition, VHS was conclusively shown to be capable of
functioning in the absence of any other viral proteins (Zelus, Stewart, and Ross, 1996).
VHS was shown to function at least in part by inducing endoribonucleolytic cleavage of
mRNA in vitro (Elgadi and Smiley, 1999), and the 5’ end of mRNA is degraded before
27
the 3’ end (Karr and Read, 1999). VHS appears to form a complex with the translation
factor eIF-4H, and the interaction is required for RNase activity possibly through
targeting to polyribosomes. The VHS and eIF-4H complex appears to decap cellular
mRNA from the 5’ end (Roizman and Knipe, 2001).
In the context of a vhs- mutant, host protein synthesis is not shut off early in
infection and α and β gene expression is extended in comparison to a wild-type virus.
These effects are due to the fact that VHS accelerates the degradation of both cellular and
viral mRNAs. As a result, during the context of a HSV-1 infection, cellular mRNA
levels diminish and as viral mRNA levels accumulate it is preferentially translated. In
this way, VHS may facilitate the transition from α to β to γ gene expression through
shortening the life of the viral mRNAs (Kwong and Frenkel, 1987; Oroskar and Read,
1987). As a γ1 gene product, VHS accumulates late in infection, but does not degrade
viral mRNA as would be expected. An interaction between VP16 and VHS at late times
blocks the degradation of RNA by VHS (Lam et al., 1996). As a result, the accumulation
of VHS into tegument complexes accomplishes two crucial functions: it brings VHS into
the infected cell to shutoff host protein synthesis, and it regulates the activity of VHS late
in infection when mostly viral mRNA is present.
Virion Transport to the Nucleus
After fusion of the virion envelope with the plasma membrane of the infected cell,
the capsid with its associated tegument complex is deposited into the cytoplasm. Some
tegument proteins disassociate from the complex and remain in the cytoplasm, while
others, such as VP16 (αTIF) are transported to the nucleus to effect their functions. The
remaining capsid-tegument complex is then transported along the cell’s microtubule
28
network to a nuclear pore. Studies have shown that dynein, the microtubule dependent
motor, is bound to capsids after entry (Sodeik, Ebersold, and Helenius, 1997), and these
results prompted the theory that incoming capsids bind to microtubules and utilize the
cell’s dynein motor to transport them to nuclear pores. Other experiments have shown
that microtubules in proximity to the plasma membrane become disrupted after entry,
suggesting that the presence of the capsid-tegument complex may destroy normal cellular
microtubule interactions (Ward et al., 1998).
After intracytoplasmic transport to the nucleus, capsids accumulate at the nuclear
envelope and become associated with nuclear pore complexes. Experiments have shown
that the VP1/2 gene is required for DNA release at the nuclear pore (Batterson, Furlong,
and Roizman, 1983; Knipe, Ruyechan, and Roizman, 1979), and antibodies to nuclear
pore transport components block capsid binding (Ojala et al., 2000). It is believed that
binding to the nuclear pore produces a structural change in the HSV capsid, resulting in
viral DNA release into the nucleus and an empty capsid docked at the nuclear pore.
Coordinate Gene Expression
During the course of an HSV infection, there are more than 80 gene products
expressed from the viral genome in a highly regulated cascade fashion (Figure 1.6) in a
number of coordinately expressed groups (Honess and Roizman, 1974). Transcription of
viral DNA takes place in the nucleus, and the host RNA polymerase II is responsible for
the transcription of all viral genes during infection (Alwine, Steinhart, and Hill, 1974;
Costanzo et al., 1977). There are several viral genes that play an important role in the
regulation of gene expression at different times post-infection.
29
HSV encodes a function responsible for transactivation of α genes immediately
after infection (Post, Mackem, and Roizman, 1981), and this was termed the α gene
transactivating factor (α-TIF). α-TIF was shown to be a component of the tegument
capable of inducing the α genes, which contain the “TAATGArATT” promoter response
element, which binds Oct-1. VP16 was identified as the viral protein responsible for the
α-TIF function (Campbell, Palfreyman, and Preston, 1984). After entry, VP16 is
released from its interaction with VHS and the tegument complex. VP16 then binds to a
cellular protein called the host cell factor (HCF) or C1 (Katan et al., 1990; Kristie and
Sharp, 1990), and HCF carries VP16 into the nucleus, at which time the VP16-HCF
complex binds to Oct-1 that is bound to viral DNA. This event forms the activator
complex that is responsible for transactivation of α genes (La Boissiere, Hughes, and
O'Hare, 1999). In addition to the Oct-1 binding sites, the promoters for α genes also
contain binding sites for other cellular transcriptional activators upstream of a TATA box
that may contribute to a basal level of α gene expression in rapidly dividing cells in the
absence of VP16 (Roizman and Knipe, 2001). The viral requirement for VP16 may be
different in resting cells, such as neuronal cells, where cellular transcription factors are
not available.
After being deposited into the nucleus of the infected cell, the HSV-1 viral
genome localizes to nuclear ND10 structures where the transcription of α genes takes
place (Maul, Ishov, and Everett, 1996). At 2 to 4 hours post infection, the viral α genes
are expressed at peak levels. There are six viral α genes: ICP0, ICP4, ICP22, ICP27,
ICP47, and US1.5. Five of the six α genes stimulate viral β gene expression in at least
some cell types. In particular, ICP4 is required for all post-α gene expression (Clements,
30
1
2
on
on
α
on
off
4
5
γ1...γ2
on
4
off
off
3
on
on
β
5
4
off
Figure 1.6: Coordinate gene expression in Herpes Simplex Virus. 1) α-TIF, a γ gene
product present in the tegument, activates initial transcription of the α genes.
2) Autoregulation of gene expression. 3) Activation of β gene expression. 4) Activation
of γ gene expression by α and β genes, release of repression of γ genes, and replication of
the viral genome. 5) γ genes turn off α and β genes late in infection (Roizman and Knipe,
2001).
31
Watson, and Wilkie, 1977; Dixon and Schaffer, 1980), and its effect is exerted at the
transcriptional level (Godowski and Knipe, 1986). However, the mechanism by which
ICP4 exerts its transcriptional control over β gene expression is unclear. ICP4 is also
responsible for down regulation of α gene products including itself and ICP0, and the
“pre α” gene products ORF P and ORF O. In this case, specific consensus binding sites
appear to be responsible for ICP4 mediated transcriptional regulation (Faber and Wilcox,
1986; Gelman and Silverstein, 1987; Kristie and Roizman, 1984; Kristie and Roizman,
1986; Muller, 1987). In addition, ICP4 has different isoforms dependent on different post
translational modifications, and it is possible that the different functions may be
somewhat dependent on the specific isoform present at different times during the course
of the infection. ICP0 promotes viral infection and gene expression, especially at a low
multiplicity of infection (MOI) where its absence leads to a virus yield that is 100 fold
less than a wild type virus (Sinclair et al., 1994; Stow and Stow, 1986). ICP0 is a
nonspecific transactivator that induces the expression of the HSV α, β, and γ genes.
Because ICP0 does not bind DNA directly, it appears to act indirectly in modulation of
transcription (Everett, Orr, and Elliott, 1991).
The HSV-1 β genes are produced at peak levels between 4 and 8 hours post
infection. Expression of the viral β genes requires the presence of functional ICP4, but is
not dependent on viral DNA synthesis. β gene products include proteins involved in viral
DNA replication and nucleotide metabolism. These viral proteins promote viral DNA
replication, which results in expression of the γ class of genes. The β genes can be
divided into two general groups: β1 genes, which are expressed shortly after the synthesis
of α proteins; and β2 genes, which are expressed with more of a delay after α gene
32
expression (Roizman and Knipe, 2001). β1 genes are exemplified by the single-stranded
DNA binding protein, ICP8, and the large subunit of ribonucleotide reductase, ICP6. β2
genes are exemplified by the viral thymidine kinase encoded by UL23. Some β2 genes
require ICP27 for expression, and this dependence may correlate with the later expression
of these genes (Roizman and Knipe, 2001).
The HSV-1 γ (late) genes are produced at peak levels only after viral DNA
replication has started, and require ICP4, ICP27 and ICP8 for efficient levels of
transcription. The transition from β to γ gene expression is also marked by a change in
the nuclear localization of transcription from sites near ND10 domains to replication
compartments as evidenced by the localization of ICP4, RNA polymerase II, and ICP22
(Knipe et al., 1987; Leopardi et al., 1997; Rice et al., 1994). γ gene products include
structural proteins, glycoproteins, and tegument components required to prepare newly
infected cells for an efficient infection. The γ genes have been subdivided into two
groups based on timing of expression and their dependence on viral DNA replication: γ1
(leaky-late) genes, which are expressed relatively early in infection and transcription is
increased only a few fold after DNA replication has occurred; and γ2 genes, which do not
accumulate in appreciable amounts until after DNA replication and are not expressed in
the presence of inhibitors of viral DNA synthesis (Wagner, 1985). Typical γ1 genes
include the major capsid protein ICP5, gB, gD, and ICP34.5, whereas typical γ2 genes
include gC, UL41 (VHS), UL36, UL38, UL20, and gK.
Viral DNA Replication
After the β genes have been expressed and translated, there are several proteins
that are localized to the nucleus where they assemble on the parental viral DNA in
33
punctuate “prereplicative sites” located near nuclear ND10 structures (Ishov and Maul,
1996; Uprichard and Knipe, 1996). Initially, viral DNA replication initiates on the
circular viral DNA, creating a “theta” structure, which as replication progresses
transitions to a rolling circle mechanism of replication producing head-to-tail
concatemers of viral DNA (Jacob, Morse, and Roizman, 1979). At this point, replication
takes place in “replication compartments” that consist of accumulating DNA molecules
and replication complexes (Quinlan, Chen, and Knipe, 1984).
There are seven viral proteins absolutely required for viral DNA replication and
sufficient to replicate a viral origin transfected into cells. These are the viral DNA
polymerase (UL30) (Purifoy, Lewis, and Powell, 1977), its accessory protein (UL42)
(Conley et al., 1981), an origin-binding protein (UL9), the single stranded DNA binding
protein (ICP8), and the helicase-primase complex that consists of three proteins: UL5,
UL8, and UL52 (Challberg, 1986; Wu et al., 1988). Host cell factors may also be
involved in DNA synthesis, and host enzymes that include the DNA polymerase αprimase, DNAligase, and topoisomerase II are almost certainly also required. There are
also three origins of replication located on the viral genome: oriS, a palindromic
sequence of 45 bp that is located in c sequences bounding the S component and present in
two copies; and oriL, a palindromic sequence of 144 bp that is located between the
transcriptional units of ICP8 and the DNA polymerase (UL29 and UL30, respectively)
(Deb and Doelberg, 1988; Frenkel et al., 1976; Knopf, 1986; Locker, Frenkel, and
Halliburton, 1982; Lockshon and Galloway, 1986; Mocarski and Roizman, 1982; Stow,
1982; Vlazny, Kwong, and Frenkel, 1982; Weller et al., 1985). The reason for the
presence of three origins of replication is not clear, although it may reflect the
34
evolutionary history of the virus. Only one origin of replication is needed for replication
to occur (Igarashi et al., 1993; Polvino-Bodnar, Orberg, and Schaffer, 1987).
The basic model for the replication of HSV viral DNA proceeds as follows. First,
the parental viral DNA is circularized upon being deposited into the nucleus of the
infected cell. After α and β gene expression, UL9 binds to specific elements in either
oriL or oriS and begins to unwind the viral DNA. UL9 then recruits the ssDNA binding
protein ICP8 to the unwound portion of the viral DNA. At this point, UL9 and ICP8
recruit the remaining five proteins to the replication forks. The helicase–primase and
viral DNA polymerase complexes assemble at each replication fork and initiate theta
form replication. Through an unknown mechanism, replication switches from theta form
to the rolling circle form of replication, and UL9 is not required for rolling circle
replication because it is not origin dependent. The rolling circle replication forms long
head-to-tail concatamers of viral DNA, which become cleaved into individual units
during packaging of viral DNA into capsids (Roizman and Knipe, 2001).
Capsid Assembly and Packaging
After DNA replication has started, the γ genes are transcribed, including those
encoding HSV-1 capsid proteins. First, empty shells containing an internal scaffold are
assembled, with the internal scaffold lost upon viral DNA packaging into the capsid.
Then the mature capsid is able to proceed along the viral egress pathway. At least some
of the initial stages of capsid assembly occur in the cytoplasm of the infected cell
(Nicholson et al., 1994; Rixon et al., 1996). The major capsid protein (VP5), the outer tip
of hexons (VP26), and a triplex protein (VP23) are not capable of nuclear localization on
their own; however, VP5 can be carried into the nucleus by VP19C, another capsid
35
triplex protein, or by pre-VP22a, a scaffolding protein. VP23 localizes to the nucleus
only in the presence of VP19C, while VP26 localizes to the nucleus only when it is
expressed with both VP5 and VP19C or pre-VP22a (Nicholson et al., 1994; Rixon et al.,
1996). Electron microscopic studies have shown that final envelopment occurs in the
nucleus; and three types of capsids, called A, B, and C capsids have been identified by
sucrose density gradient ultracentrifugation (Gibson and Roizman, 1972). All three types
of capsids are about 120 nm in diameter with an outer shell composed of hexons and
pentons made up of VP5. The individual capsomeres are linked by triplex structures
consisting of VP19C and VP23, the two minor capsid proteins. Each triplex consists of
one molecule of VP19C and two molecules of VP23 (Newcomb et al., 1993). C-capsids
are mature capsids that contain packaged viral DNA and can proceed to become
infectious virions by budding through the nuclear membrane into the perinuclear space
(Perdue et al., 1976). In contrast, A and B capsids lack viral DNA, but B capsids are
filled with VP22a and VP21, the cleaved scaffolding proteins, and a viral protease VP24
(Gibson and Roizman, 1972; Newcomb et al., 1993). The internal proteins VP22a,
VP21, and VP24 are removed upon packaging of viral DNA to form C capsids (Davison,
Rixon, and Davison, 1992; Gibson and Roizman, 1972). A capsids do not contain viral
DNA or scaffolding proteins and are believed to be an abortive result of failed attempts at
DNA packaging.
In terms of the order of assembly, once in the nucleus, VP5-pre-VP22a complexes
come together as a result of self assembly of pre-VP22a. The triplex proteins VP19C and
VP23 are then added to form a partial capsid. As hexons and pentons are added, the
structure assembles into a round procapsid (Newcomb et al., 1996). At this point, the
36
procapsid undergoes a structural transformation and becomes polyhedral (Church and
Wilson, 1997; Thomsen et al., 1995; Trus et al., 1996). It is unknown at this time
whether the round or polyhedral B capsids are the structure in which viral DNA is
packaged (Lee, Irmiere, and Gibson, 1988; Perdue et al., 1974; Trus et al., 1996).
Encapsidation of viral DNA is a process in which unit length monomers of viral DNA are
cleaved from concatamers and packaged into preformed B capsids, as originally shown
for pseudorabies virus (Ladin, Blankenship, and Ben-Porat, 1980; Ladin et al., 1982).
Cleavage of concatamers occurs at specific sites and requires the pac1 and pac2
packaging signals (Deiss, Chou, and Frenkel, 1986; Smiley, Duncan, and Howes, 1990;
Varmuza and Smiley, 1985). The process of encapsidation of viral DNA into capsids is
complex and requires several gene products, including the UL6, UL15, UL25, UL28,
UL32, UL33, UL36 and UL37 gene products, but the mechanism of viral DNA
packaging is not well defined (Roizman and Knipe, 2001).
Herpesvirus Egress
Nuclear Egress: Primary Envelopment
After encapsidation of genomic DNA, mature nucleocapsids aquire a primary
envelope by budding through the inner nuclear membrane into the perinuclear space
(Vlazny, Kwong, and Frenkel, 1982). Two conserved herpesvirus proteins, UL31 and
UL34 have recently been shown to be involved in the budding process. UL34 is Cterminally anchored membrane protein present in the inner and outer nuclear membrane
(Klupp, Granzow, and Mettenleiter, 2000), while UL31 is a nuclear phosphoprotein also
present in the nuclear membrane of infected cells (Fuchs et al., 2002; Reynolds et al.,
2001). The UL31 protein requires UL34 for proper nuclear targeting (Fuchs et al., 2002),
37
TRANS GOLGI
NETWORK (TGN)
IV
GOLGI
TGN / ENDOSOME
DERIVED VESICLE
ENDOPLASMIC
RETICULUM (ER)
III
UL20p
gK
II
PERINUCLEAR
SPACE
I
NUCLEUS
Figure 1.7: Representation of HSV-1 virion morphogenesis and egress. (I) Mature
capsids budding through the inner nuclear membrane into the perinuclear space; (II) Deenvelopment of perinuclear virions at the outer nuclear membrane; (III) Re-envelopment
of cytoplasmic capsids by budding into cytoplasmic vesicles; (IV) Final egress to the
extracellular space. The steps at which UL20 and gK are thought to function in virion
egress are indicated by red arrows.
38
whereas the UL34 protein appears to possess an intrinsic nuclear targeting signal (Klupp,
Granzow, and Mettenleiter, 2000). However, the nuclear targeting of UL34 is increased
by the presence of UL31 (Fuchs et al., 2002; Reynolds et al., 2001). Analysis of UL31
and UL34 deletion mutants has indicated that the absence of either protein results in a
drastic impairment in primary envelopment with capsids trapped within the nucleus
(Chang et al., 1997; Fuchs et al., 2002; Klupp, Granzow, and Mettenleiter, 2000;
Reynolds et al., 2001; Roller et al., 2000). In addition, partial dismantling of the nuclear
lamina has been observed after HSV infection and may be required so that intranuclear
capsids are able to gain access to the inner nuclear membrane (Scott and O'Hare, 2001).
Multiple lines of evidence indicate that UL31 and UL34 play a role in dismantling the
nuclear lamina in infected cells. Overexpression of the UL31 protein in uninfected cells
was sufficient to relocalize lamin A/C from the nuclear rim into nucleoplasmic
aggregates, while overexpression of UL34 was sufficient to relocalize some lamin A/C
into the cytoplasm (Reynolds, Liang, and Baines, 2004). More importantly, both UL31
and UL34 have been shown to directly bind lamin A/C, suggesting that the UL31 and
UL34 proteins modify the conformation of the nuclear lamina in infected cells, possibly
by direct interaction with lamin A/C. Given that the nuclear lamina potentially excludes
nucleocapsids from envelopment sites at the inner nuclear membrane, the lamina
alteration may reflect a role of the UL31/UL34 protein complex in perturbing the lamina
to promote nucleocapsid egress from the nucleus (Baines 2004). It is unclear whether
additional primary envelope and tegument proteins exist that are required for primary
envelopment to take place. The UL11 protein has been proposed to function in primary
39
envelopment and increases the efficiency of envelopment at this step (Baines and
Roizman, 1992; MacLean et al., 1992).
Egress from the Perinuclear Space: De-Envelopment
The subsequent steps in HSV-1 egress following primary envelopment have been
in dispute for some time. Two models were proposed to explain virion egress. The first
model, sometimes called the “lumenal” pathway, suggests that perinuclear virions retain
their primary envelope and integrity as they leave the cell through the secretory pathway
(Campadelli-Fiume et al., 1991; Darlington and Moss, 1968; Johnson and Spear, 1982;
Johnson et al., 2001). In this model, virion glycoproteins are modified in transit to the
plasma membrane and perinuclear virions should contain the entire complement of
tegument and envelope proteins present on mature extracellular virions. The alternative
model proposes that the primary envelope of perinuclear virions fuses with the outer
nuclear lamellae, resulting in loss of the primary envelope and some tegument
components and translocation of the capsid into the cytoplasm of the infected cells
(Figure 1.7, step II) (Enquist et al., 1998; Mettenleiter, 2000). In this “de-envelopment
re-envelopment” pathway, final tegumentation and envelopment (re-envelopment) occur
in cytoplasmic compartments (Figure 1.7, step III). In addition, in this model perinuclear
and intracytoplasmic/extracellular virions should differ in composition.
There is evidence that supports both the “lumenal” and the alternative “deenvelopment re-envelopment” pathways. However, the vast majority of recent evidence
supports the “de-envelopment re-envelopment” model of HSV-1 virus egress. The first
line of evidence supporting the alternative model is that primary envelope and primary
tegument of perinuclear virions clearly differ in ultrastructural appearance from the final
40
envelope and final tegument of extracellular virions when observed by electron
microscopy (Gershon et al., 1994; Granzow et al., 2001). In addition, while UL31 and
UL34 localize on nuclear membranes and perinuclear virions, the UL31 and UL34
proteins were not detected in extracellular virions (Reynolds et al., 2002). In further
support, the major tegument proteins UL46 and UL49 are present in
intracytoplasmic/extracellular virions but absent from perinuclear virions (Klupp,
Granzow, and Mettenleiter, 2000; Mettenleiter, 2002b). Also, the phospholipid
composition on the final virion envelope differs substantially from the composition of the
nuclear membrane, explained only by a two-step envelopment model of HSV virion
egress (van Genderen et al., 1994). Electron microscopic analysis has also demonstrated
the fusion of primary envelopes with the outer nuclear membrane with many
herpesviruses including HSV-1 (Granzow et al., 2001; Harms et al., 2000). As a result,
the two-step “de-envelopment re-envelopment” model of virion egress is congruent with
both biochemical and morphological data and constitutes a unified model for the
morphogenesis of herpesviruses (Mettenleiter, 2002b).
The mechanism of virion de-envelopment from the perinuclear space is unclear.
Deletion of the major glycoproteins involved in other membrane fusion events does not
affect the de-envelopment process as indicated by multiple studies (Cai et al., 1987;
Granzow et al., 2001; Jayachandra, Baghian, and Kousoulas, 1997; Steven and Spear,
1997). It is possible that these glycoproteins function in a redundant manner, and that
only deletion of multiple genes encoding glycoproteins would have an observable effect
on virion de-envelopment at the outer nuclear membrane. This possibility is addressed
later in Chapter III.
41
Tegumentation in the Cytoplasm
While the complexity of the herpesvirus capsid is not much different than that of
other icosahedral viral capsids such as the picornavirus capsid, the complexity of the
herpesvirus tegument is drastically increased. At least 15 proteins have been identified
that are part of the HSV-1 tegument. Even more proteins make up the tegument of other
herpesviruses such as VZV or CMV (Gibson, 1996; Spengler et al., 2001). Recent data
indicate that tegumentation follows an intrincate pattern of protein-protein interactions
that contains significant levels of redundancy built in, at least as observed in cell culture
(Mettenleiter, 2002b). The tegument proteins interact with the capsid on one side and the
viral envelope proteins on the other side in order to link the structural components to the
final envelope of the HSV-1 virion and to secure the integrity of the virus particle (Figure
1.7) (Mettenleiter, 2002b). For a long time the herpesvirus tegument was considered
unstructured; however, cryoelectron microscopic analysis has indicated that at least the
innermost portion of the tegument located adjacent to the capsid exhibits icosahedral
symmetry, resulting from the interaction of a large tegument protein, presumably UL36,
with the pentons of the capsid (Zhou et al., 1999). The UL36 gene product has been
shown to interact with the major capsid protein VP5, which forms both the pentons and
hexons (Machtiger et al., 1980; McNabb and Courtney, 1992; Newcomb et al., 1993).
So, it appears that the first layer of tegument around the capsid is structured and
composed of UL36. UL37 has been shown to interact with UL36 in coimmunoprecipitation and yeast two-hybrid experiments, and capsids that accumulate in
the absence of UL37 contain UL36 (Klupp et al., 2002). UL36 and UL37 appear to be the
only tegument proteins conserved in all herpesvirus subfamilies, and the absence of the
42
HSV-1 UL36 and UL37 proteins abolishes virus maturation (Desai et al., 2001; Desai,
2000; Klupp et al., 2001). Unfortunately, the subsequent steps in capsid tegumentation
are still largely undefined. Virion morphogenesis still proceeds in the absence of several
other tegument proteins, including UL13, US3 (Purves et al., 1987), UL41, UL46, UL47
(Rafield and Knipe, 1984; Roizman and Knipe, 2001), and UL49 (Mettenleiter, 2002b).
So far the most dramatic effect has been seen upon deletion of UL48 (α-TIF), which is
responsible for transducing α gene promoters and makes up a major part of the virus
tegument (Batterson and Roizman, 1983; Heine et al., 1974). The absence of UL48 may
interfere with a later step in virion assembly, presumably affecting tegumentation and reenvelopment in the cytoplasm (Mossman et al., 2000). Interestingly, UL48 has been
shown to interact with other tegument components, UL49 and UL41 (VHS) (Smibert et
al., 1994). Mutant forms of UL41 that do not bind UL48 do not become incorporated
into the virion (Read, Karr, and Knight, 1993). Cross-linking studies have indicated that
UL48 may interact with gB, gD, and gH (Zhu and Courtney, 1994), although these
interactions have not been confirmed through more stringent methods.
Final Envelopment at the Trans-Golgi Network/Endosomes and Egress to
Extracellular Spaces
Following tegumentation in the cytoplasm, HSV-1 capsids bud into cytoplasmic
vesicles derived from the trans-Golgi Network (TGN) or endosomes, acquiring their final
envelope (Figure 1.7, step III). Although the mechanism of secondary envelopment is not
well defined, the study of mutants that contain deletions of certain glycoproteins has shed
light on some requirements of the final envelopment event. By far the most dramatic
effect on secondary envelopment and final egress is caused by the deletion of either
UL20 or gK, and ultrastructural study of UL20-null and gK-null infected cells reveals a
43
dramatic accumulation of unenveloped and aberrantly enveloped capsids in the cytoplasm
of infected cells with a marked absence of extracellular virus. With regard to gK-null and
UL20-null virus titers, there is also a corresponding lack of infectious virus in the
supernates of infected cells, with close to wild-type levels of intracellular virus (Foster et
al., 2004a; Jayachandra, Baghian, and Kousoulas, 1997). The ultrastructural phenotypes
of UL20-null and gK-null viruses are covered in detail in Chapter III and Chapter IV.
Following the final envelopment step, nascent virions must still egress to
extracellular spaces. The current belief is that cellular transport machinery is utilized in
this step, although the mechanisms are unclear. It is apparent that the late stages in viral
egress may differ depending on the cell type that is infected. For example, deletion of gE
and gI, which have been shown to interact and form a complex, does not impair
productive replication of HSV-1 (Enquist et al., 1998). However, in polarized epithelial
cells, wild-type HSV-1 virions are sorted predominantly to cell junctions, while gE/gI
null virions are non-specifically released into supernatants (Dingwell and Johnson, 1998).
Delivery of virus particles to cell junctions would be expected to enhance virus spread
and enable viruses to avoid host immune defenses.
Deletion of multiple genes encoding envelope glycoproteins has revealed that
certain aspects of final envelopment and egress may feature redundant mechanisms, each
of which is sufficient for function, at least in cell culture. For instance, while neither
gE/gI or gM is required for productive replication in PrV or HSV-1, simultaneous
deletion of gE/gI and gM results in drastic inhibition of plaque formation and replication
in PrV (Brack et al., 1999). In addition, simultaneous deletion of UL11 and gM resulted
in similar egress defect in PrV (Kopp et al., 2004). In contrast, simultaneous deletion of
44
gE/gI and gM does not have a major effect in HSV-1 (Browne, Bell, and Minson, 2004);
however, a triple mutant lacking gD, gE, and gI had a severe defect in the final
envelopment step. The authors proposed that HSV gD and the gE/gI heterodimeric
complex act in a redundant fashion to anchor the virion envelope onto tegument-coated
capsids (Farnsworth, Goldsmith, and Johnson, 2003). In contrast, it is also possible that
deletion of multiple glycoproteins indirectly disrupts the integrity of many different
protein-protein interactions required for the final envelopment step and that the resulting
phenotype results from the compounded effects of an abnormal glycoprotein profile.
More study of the reason for defects in double and triple-null viruses is required before
conclusions can be drawn regarding mechanisms of final envelopment.
Formation of Light (L) Particles
The presence of egressing capsids in the cytoplasm is not required for tegument
assembly and subsequent envelopment to take place. The formation of extracellular
herpesvirus light (L) particles that lack capsids, consist of only tegument and envelope,
and are formed independent of normal virus maturation have been thoroughly studied in
HSV-1 (McLauchlan and Rixon, 1992; Rixon, Addison, and McLauchlan, 1992). L
particles appear to contain the full complement of tegument proteins as well as an
authentic envelope containing all appropriate glycoproteins. Studies of PrV have shown
that L particle formation occurs in the absence of UL36 and UL37, which are tegument
components that are critical for capsid tegumentation. It is possible that normal
tegumentation may be centered on UL36 and UL37 interactions with the capsid and that
UL49 may be required for proper interactions with viral glycoproteins in the final
budding and envelopment steps. In the absence of capsids, tegument assembly could
45
proceed anchored on UL49, resulting in the formation of L particles (Mettenleiter,
2002b).
HSV-1 Glycoproteins and Their Putative Functions
Herpes simplex viruses specify at least eleven virally encoded glycoproteins: gB,
gC, gD, gE, gG, gH, gI, gJ, gK, gL, gM, and potentially gN (UL49.5). HSV also encode
several non-glycosylated membrane associated proteins: UL20, US9, UL24, UL43, and
UL34. Herpesvirus membrane proteins function in several important roles including virus
entry via fusion of the viral envelope with cellular membranes, intracellular virion
morphogenesis and egress, cell-to-cell spread, and virus-induced cell fusion
(Mettenleiter, 2002a; Mettenleiter, 2002b; Roizman and Knipe, 2001; Spear, 2004; Spear
and Longnecker, 2003). At least 11 membrane proteins are present on the surface of the
virion envelope (Roizman and Knipe, 2001).
Glycoprotein B
The HSV-1 UL27 gene encodes the 904 aa glycoprotein B (gB). gB is highly
conserved across all subfamilies of herpesviruses. HSV-1 gB is homotrimeric (aa) type I
membrane glycoprotein composed of a 696-aa ectodomain that is N-glycosylated at
multiple sites (Claesson-Welsh and Spear, 1986; Claesson-Welsh and Spear, 1987;
Highlander et al., 1991; Laquerre, Person, and Glorioso, 1996; Spear, 1993; Whitley,
2001), a 69-aa transmembrane domain, and a 109-aa carboxy-terminal domain. The
cytoplasmic domain of gB is the longest among HSV-1 glycoproteins, implying a crucial
role for this domain in gB-mediated functions. A variety of evidence indicates that gB
plays important roles in membrane fusion phenomena during virus entry and virusinduced cell fusion: HSV-1 mutant viruses lacking gB are not able to enter into cells (Cai
46
et al., 1987) due to a post-attachment defect that can be resolved by polyethelene glycol
mediated fusion of viral envelopes with cellular membranes (Cai, Gu, and Person, 1988);
Single amino acid substitutions and truncations of the carboxyl terminus of gB cause
extensive virus-induced cell fusion (Baghian et al., 1993; Bzik et al., 1984; Cai et al.,
1988; Gage, Levine, and Glorioso, 1993). Transient co-expression of gB with gD, gH and
gL causes cell-to-cell fusion, which is substantially increased by carboxyl terminal
truncations of gB (Foster, Melancon, and Kousoulas, 2001; Haan, Lee, and Longnecker,
2001; Klupp, Nixdorf, and Mettenleiter, 2000; Pertel, 2002). These results suggest a
direct role for gB in membrane fusion and suggest that perturbations of the carboxyl
terminal domains of gB facilitate gB-mediated cell-to-cell fusion. gB is viewed as the
primary component of herpesvirus membrane fusion machinery.
Glycoprotein C
The 1,536 bp HSV-1 UL44 gene encodes for the 511 aa long precursor of
glycoprotein C (gC) (Frink et al., 1983). gC contains a 25 aa signal sequence at the Nterminus, a long 453 aa extracellular domain, a 23 aa transmembrane anchoring domain,
and a short 10 aa C-terminal cytoplasmic tail (Homa et al., 1986). The first contact of
virions with the surface of susceptible cells is through glycosaminoglycans (GAGs). gC
can mediate the initial binding of HSV-1 virus to a cell surface GAG, heparan sulfate
(HS). The heparan sulfate binding site of gC has been localized to the N-terminal 120 aa
(Tal-Singer et al., 1995). The aminoglycoside neomycin interferes with the binding of
virus to HS mediated by gC (Herold et al., 1994). Although gC is dispensable for the
infection of cultured cells, its presence can increase the efficiency of virus binding almost
10-fold, at least for HSV-1. The main function of gC binding to GAGs seems to be
47
concentration of the virus on cell surfaces, enabling the more stable interaction of gD
with an entry receptor.
Another important function of gC is its ability to inactivate complement to
facilitate immune evasion by HSV-1. The gC of many herpesviruses has the ability to
bind C3b (Eisenberg et al., 1987; Friedman et al., 1984; Huemer et al., 1993; Huemer et
al., 1995). The presence of gC protects the virus and infected cell from antibody (Ab)independent complement neutralization and cell lysis (Lubinski et al., 1999; Lubinski et
al., 1998), and gC is an important virulence factor in vivo (Isaacs, Kotwal, and Moss,
1992; Kotwal et al., 1990; Sahu et al., 1998). HSV-1 mutant viruses lacking gC are
rapidly inactivated by human complement, resulting in up to a 5000 fold loss of titer
(Friedman et al., 1996). While Ab is not required for neutralization of a gC null virus,
the presence of Ab enhances neutralization (Friedman et al., 1996; Harris et al., 1990).
An unresolved issue regarding gC function is the relationship of gC to virusinduced syncytia formation. Several spontaneously arising syncytial virus strains were
found to be gC-deficient. In particular the MP strain variant MP10311 contains a
syncytial mutation in gK and is gC-deficient (Bartoletti et al., 1985). In general, the
syncytial phenotype of these strains was found to be more effectively expressed in the
absence of gC, especially in Hep-2 cells (Bond, Person, and Warner, 1982; Goodman and
Engel, 1991; Manservigi, Spear, and Buchan, 1977; Pertel and Spear, 1996; Tognon et
al., 1984). One possible explanation of the enhancement of the syncytial phenotype is
that in the absence of gC, because extracellular virus does not bind to cell surfaces as
rapidly, virus-induced cell fusion acts as the preferential method of cell-to-cell spread of
HSV-1.
48
Glycoprotein D
The 1182 bp long US6 gene specifies a 394 aa precursor to glycoprotein D (gD).
gD consists of a 25 aa signal peptide, a long 315 aa ectodoamin containing three
glycosylation sites (McGeoch et al., 1985; Watson et al., 1982), a 22 aa transmembrane
domain, and a 32 aa C-terminal cytoplasmic domain (Minson et al., 1986). The
ectodomain of gD has 6 cysteine residues at positions 66, 106, 118, 127, 189, 202
forming disulfide bonds (Long et al., 1992). Importantly, gD has been shown to act as the
viral ligand for all known HSV-1 entry receptors (Spear, Eisenberg, and Cohen, 2000).
As a result, gD is absolutely required for virus entry and virus-induced cell-to-cell fusion
to occur. As discussed earlier, HSV-1 gD has the ability to bind HVEM, nectin-1, nectin2, and other cell surface receptors to mediate virus entry. In addition, cells that express
gD are resistant to HSV infection in a dose-dependent manner due to a saturation of the
corresponding entry receptors (Campadelli-Fiume et al., 1988; Johnson, Burke, and
Gregory, 1990). X-ray structures of HSV-1 gD alone and in complex with HVEM
revealed that a portion of gD assumes an Ig-like fold with unconventional disulfidebonding patterns (Carfi et al., 2001). There is an N-terminal extension from the Ig-like
fold that forms a hairpin loop in the complex with HVEM, but is disordered in the
crystals of gD alone. The contacts in gD for HVEM have been localized to amino acids 7
to 15 and 24 to 32 within the N-terminal hairpin (Spear and Longnecker, 2003).
Mutagenic analysis of HSV-1 gD revealed that the first 32 amino acids of the N-terminal
extension have a critical role in functional interactions of gD with all the HSV entryfusion receptors, except for nectin-1, and that the amino acid sequence within this region
49
governs whether nectin-2 can be recognized as an entry and fusion receptor (Yoon et al.,
2003; Zago and Spear, 2003).
Glycoproteins E and I
The 1652 bp long US8 gene encodes the 550 aa glycoprotein E (gE) (Lee, Para,
and Spear, 1982), while the 1172 bp long US7 gene encodes 390 aa glycoprotein I (gI)
(Longnecker et al., 1987). HSV, VZV, and PrV express a heterodimer of gE and gI that
functions to mediate cell-to-cell spread of virus in epithelial and neuronal tissues (Balan
et al., 1994; Card et al., 1992; Cohen and Nguyen, 1997; Dingwell et al., 1994; Dingwell
and Johnson, 1998; Jacobs et al., 1993; Johnson and Feenstra, 1987; Johnson et al., 1988;
Kimura et al., 1998; Kritas, Pensaert, and Mettenleiter, 1994; Kudelova et al., 1991;
Mettenleiter et al., 1987b; Mulder et al., 1994; Neidhardt, Schroder, and Kaerner, 1987;
Tirabassi and Enquist, 1998; Tirabassi et al., 1997; Whealy et al., 1993). HSV and PRV
gE/gI complexes are required for efficient spread of viruses between certain cultured
epithelial cells, neurons, and other polarized cells with extensive cell junctions, but are
not needed for spread between highly transformed, nonpolarized cells, such as Vero or
HeLa cells, which do not form cell junctions (Dingwell et al., 1994; Dingwell, Doering,
and Johnson, 1995; Mettenleiter et al., 1987a; Tirabassi et al., 1997; Wisner et al., 2000;
Zsak et al., 1992). For example, plaques formed by a gE-negative HSV mutant on
monolayers of a keratinocyte cell line included eightfold fewer cells than plaques
produced by wild-type HSV-1, yet there was no difference in cell-to-cell spread in
monolayers of HeLa cells (Wisner et al., 2000). Moreover, PRV and HSV gE/gI
complexes are required for spread within synaptically connected neuronal circuitry in the
peripheral and central nervous systems (Babic et al., 1996; Dingwell, Doering, and
50
Johnson, 1995; Mulder et al., 1994; Tirabassi and Enquist, 1998; Tirabassi et al., 1997;
Whealy et al., 1993).
gE and gI are extensively complexed in virus-infected cells (Johnson and
Feenstra, 1987; Johnson et al., 1988), and it is the gE/gI complex that functions in cell-tocell spread (Dingwell et al., 1994; Dingwell, Doering, and Johnson, 1995; Johnson and
Feenstra, 1987; Johnson et al., 1988; Olson and Grose, 1998; Tirabassi et al., 1997;
Zuckermann et al., 1988). In contrast to their effects on cell-to-cell spread, HSV and PRV
gE/gI complexes do not appear to be required for entry of cell-free virus, i.e., virus
particles applied to the apical surfaces of cells (Dingwell et al., 1994; Mettenleiter et al.,
1987a). HSV-1 gE/gI localizes specifically to the TGN during early phases of infection
but moves out to cell junctions at intermediate to late times. One mechanism by which
gE/gI facilitates cell-to-cell spread of HSV-1 involves the sorting of newly assembled
virions to lateral cell surfaces and cell junctions. Mutant HSV-1 lacking gE accumulated
more extensively in the cytoplasm, at apical cell surfaces, and in cell culture supernatants
than did wild-type HSV-1 particles, which were found predominantly at cell junctions
(Alconada et al., 1999; Kimura et al., 1998).
The gE/gI complex is also known as the HSV Fc Receptor (FcR) because of its
high affinity for the Fc portion of IgG. Through binding of the Fc portion of IgG, the
gE/gI complex contributes to the immune evasion capabilities of HSV-1, reducing the
effectiveness of antibody-dependent cellular cytotoxicity (ADCC) targeting the virus and
infected cells (Eberle et al., 1995).
51
Glycoprotein G
The 716 bp long US4 gene encodes the 238 aa glycoprotein G (gG). During
infection, gG is incorporated into nuclear and cytoplasmic membranes in the cell (Frame,
Marsden, and McGeoch, 1986; Sullivan and Smith, 1987). The relevant function of gG
in the herpesvirus life cycle has been difficult to identify. Mutant HSV-1 lacking US4
showed no discernible phenotypic abnormalities relative to the wild-type HSV-1 virus in
non-polarized cells in culture, and the absence of gG resulted in only marginal
attenuation of the virus in certain in vivo models (Atkinson, Barr, and Timbury, 1978).
However, at the primary site of infection, HSV-1 is required to infect the apical surfaces
of mucosal epithelial cells. Recombinant viruses lacking gG were shown to be defective
in the gC-dependent infection of the apical surfaces of polarized epithelial cells in culture
and also in infection of the apical surfaces of corneal epithelial cells in vivo (Tran et al.,
2000). It is not known whether the gG function in infection is mediated in a cooperative
manner with gC, whereby gG would assist with gC binding, or in an independent manner,
whereby gG would be necessary in a second step that renders the gC interaction
irreversible.
Glycoproteins H and L
The 2517 bp UL22 gene encodes the 838 aa glycoprotein H (gH). gH is a type I
membrane glycoprotein containing an 18 aa signal peptide, a long 785 aa ectodomain, a
single 21 aa transmembrane hydrophobic domain close to the C-terminus, and a 14 aa Cterminal cytoplasmic tail. The 675 bp UL1 gene encodes the 224 aa glycoprotein L. gL
contains a 25 aa signal peptide; however, unlike other herpesvirus glycoproteins, gL does
not contain a transmembrane domain. It appears that gL is not an integral membrane
52
protein; rather, its membrane association and incorporation into virus particles is
dependent on its interaction with gH (Dubin and Jiang, 1995). Infected cell membranes
contain a heterodimer of gH and gL (gH/gL). When gH is expressed in transfected cells
in the absence of gL, the resulting gH polypeptide is neither folded or processed
correctly. The malformed gH remains in the endoplasmic reticulum and undergoes selfaggregation (Foa-Tomasi et al., 1991; Forrester et al., 1991; Roberts et al., 1991).
Similarly, cells infected with a gL-null virus do not produce virions containing gH.
When cells are infected with a gH-null virus, gL is neither correctly processed nor
incorporated into the plasma membrane or viral envelope (Hutchinson et al., 1992a;
Roop, Hutchinson, and Johnson, 1993). Due to the lack of a transmembrane region, the
gL polypeptide is secreted into the medium (Dubin and Jiang, 1995). When both UL22
and UL1 are cotransfected into cells, the antigenic conformation of the gH/gL
heterodimer is identical to that of virus infected cells, indicating that no other viral
proteins are necessary for heterodimer formation. The mutual interaction of both
glycoproteins is mediated by the N-terminal region of the first 69 aa of gL after cleavage
of the 25 aa signal peptide (Roop, Hutchinson, and Johnson, 1993), while gH interacts
through a central region of the its extracellular domain. A properly formed gH/gL
heterodimer is required for both virus entry and virus-induced cell-to-cell fusion.
Although viruses lacking gH/gL are unable to enter cells, they are able to attach to the
cell surface. Therefore, the role of gH/gL in virus entry is during the virus envelopeplasma membrane fusion event and is not required for virion attachment or receptor
binding. An interesting result concerning the mechanism of gH/gL functions was
obtained when using anti-gL mAbs directed to the C-terminal portion of gL. Certain
53
mAbs inhibited virus-induced cell-to-cell fusion resulting from syncytial mutations but
not virus entry, indicating a fundamentally different mechanism may be at work in the
two processes (Novotny, Parish, and Spear, 1996).
Glycoprotein J
The 279 bp US5 gene encodes the 92 aa glycoprotein J (gJ) (Ghiasi et al., 1998).
gJ contains a predicted signal peptide as well as a hydrophobic transmembrane domain.
The current function of gJ is unknown. Deletion viruses that lack the US5 ORF show no
phenotypic deficiencies either in vitro or in vivo (Balan et al., 1994). More specifically, a
gJ-null HSV-1 was capable of multiplying from an inoculation site in mice and entering
and replicating in the peripheral and central nervous system (Balan et al., 1994).
However, it has been shown that another gene in the unique long (UL) region of the viral
genome, UL27.5, has amino acid sequences common to US5. UL27.5 was discovered
due to the fact that antibodies generated against the 23 kDa gJ cross reacted with an
unknown 43 kDa product (Chang et al., 1998). In contrast to gJ, UL27.5 accumulates in
the cytoplasm of infected cells; nevertheless, it may be possible that UL27.5 and gJ could
function redundantly and an observable change in phenotype would not be seen unless
both US5 and UL27.5 were disrupted.
Glycoprotein K
The 1017 bp UL53 gene encodes the 338 aa precursor of glycoprotein K (gK)
(Debroy, Pederson, and Person, 1985; Pertel and Spear, 1996). gK contains a cleavable
30 aa signal sequence as well as two asparagine residues at positions 48 and 58 that are
glycosylated by N-linked mannose (Hutchinson et al., 1992b; Ramaswamy and Holland,
1992). gK is extremely hydrophobic and was originally proposed to contain four
54
transmembrane domains, with both the N-terminus and C-terminus predicted to lie on the
lumenal/extracellular side of cellular membranes (Debroy, Pederson, and Person, 1985).
In contrast, experiments with in vitro translated gK in the presence of microsomal
membranes predicted gK to contain three transmembrane domains, with the C-terminal
tail located intracellularly (Debroy, Pederson, and Person, 1985; Mo and Holland, 1997;
Ramaswamy and Holland, 1992). However, recent experiments by our laboratory
utilizing epitope tags inserted into specific regions of gK confirmed the original
prediction for the membrane topology of gK (Foster, Alvarez, and Kousoulas, 2003).
Original attempts to isolate gK-deficient viruses through substitution of UL53
with the lacZ gene were unsuccessful and it was determined that gK was essential for
HSV replication in vivo (MacLean et al., 1991). A subsequent attempt at the isolation of
a F-strain gK-null virus was successful, resulting in an insertion-deletion mutation in gK
that expressed a fusion protein containing the N-terminal 112 aa of gK. The aberrant
virions were significantly less infectious and did not reach the extracellular space, with a
high number of naked capsids and aberrant virions in the cytoplasm (Hutchinson and
Johnson, 1995). Furthermore, cells infected with the F strain gK-null virus caused cell
fusion in 143TK- cells. However, a KOS strain containing a more precise deletion of the
UL53 gene resulted in a slightly different phenotype (Jayachandra, Baghian, and
Kousoulas, 1997). While the egress of virions in the KOS gK-null virus was still
severely impaired relative to a wild-type virus, the KOS gK-null did not cause cell fusion
in 143TK- cells. A KOS virus that was constructed to mirror the F strain insertiondeletion resulted in syncytia formation in 143TK- cells, so the syncytial phenotype of the
“gK-null” viruses was attributed to the expression of the N-terminal 112 aa of gK
55
(Jayachandra, Baghian, and Kousoulas, 1997). Importantly, a precise insertion-deletion
gK mutation recently constructed in an F-strain BAC was not syncytial and showed a
similar egress defect, confirming previous results (see Chapter III).
Studies of gK localization in infected cells had previously shown that gK was
exclusively localized in the perinuclear spaces and contained unprocessed carbohydrates
added in the rough endoplasmic reticulum, indicating that gK was not transported to the
Golgi complex and never reached the cell surface (Hutchinson, Roop-Beauchamp, and
Johnson, 1995). Interestingly, while spontaneous mutations leading to syncytia
formation can arise in UL20 (Baines et al., 1991; MacLean et al., 1991), UL24 (Jacobson,
Martin, and Coen, 1989; Sanders, Wilkie, and Davison, 1982), UL27 (gB) (Bzik et al.,
1984; Pellett et al., 1985), and UL53 (gK) (Bond and Person, 1984; Debroy, Pederson,
and Person, 1985; Pogue-Geile et al., 1984; Ruyechan et al., 1979), a large majority of
syncytial mutations map to the UL53 (gK) gene. The number of syncytial mutations
found in gK would seem to be at odds with data indicating that gK never reaches the
surface of infected cells; however, data obtained in our laboratory utilizing viruses
engineered to contain epitope tags in the putative extracellular domains of gK indicates
that gK does indeed reach the surface of infected cells, resolving the paradox of how gK
could potentially function in virus induced cell-to-cell fusion (Foster, Alvarez, and
Kousoulas, 2003). In transiently transfected cells, gK localized in the ER and did not the
cell surface. It was originally reported for PrV that gK is not correctly processed in a
UL20 deletion mutant, and that coexpression of gK and UL20 restored gK processing at
least partially (Dietz et al., 2000). In support of these original findings, our laboratory
showed that coexpression of gK and UL20 in transfected cells restored the transport of
56
gK to cell surfaces (Foster, Alvarez, and Kousoulas, 2003). Nevertheless, while
coexpression of UL20 also restored the transport of syncytial gK genes in transfected
cells, syncytia formation did not result in these experiments, indicating that additional
viral proteins are needed to activate the membrane fusion potential of gK (Foster,
Alvarez, and Kousoulas, 2003).
Glycoprotein M and UL49.5 (gN)
The 1422 bp UL10 gene encodes the 473 aa glycoprotein M (gM), which contains
eight predicted hydrophobic transmembrane regions (Babic et al., 1996; Baines and
Roizman, 1993). gM is a component of the virion envelope and is present in the
cytoplasmic membranes of infected cells (MacLean, Robertson, and Jamieson, 1993).
Viruses deficient in the expression of gM exhibit a 10 to 20 fold reduction in virus titers
in Vero or BHK cells and form slightly smaller plaques (MacLean et al., 1991; MacLean,
Robertson, and Jamieson, 1993). The 276 bp UL49.5 (UL49A) gene encodes a 91 aa
integral membrane protein (Baines and Roizman, 1993). The UL49.5 polypeptide
contains a 23 aa cleaved signal peptide and a C-terminal hydrophobic transmembrane
domain (Barnett et al., 1992). There are also potential sites for O-glycosylation, leading
to the naming of UL49.5 as glycoprotein N (gN) in some cases. In PrV, the UL49.5
homolog was found to be O-glycosylated and incorporated into virions as a 14kDa
envelope protein (Jons et al., 1996). However, the UL49.5 gene product showed no
evidence of glycosylation in HSV-1 infected cells, and appeared to be abundantly present
in virions, linked to the tegument by disulfide bonds (Adams et al., 1998). While initial
attempts at deletion of UL49.5 were unsuccessful and indicated the possibility that
UL49.5 was required for virus replication (Barker and Roizman, 1992), a more recent
57
attempt utilizing a more precise deletion resulted in a UL49.5-deficient virus that showed
no phenotypic differences in plaque yield or morphology and only a marginal two-fold
difference in virus yield (Adams et al., 1998).
Glycoprotein M and UL49.5 (gN) are conserved throughout alpha, beta-, and
gammaherpesviruses and have been shown to form a complex in a number of
herpesviruses analyzed to date (Jons, Dijkstra, and Mettenleiter, 1998; Koyano et al.,
2003; Lake, Molesworth, and Hutt-Fletcher, 1998; Mach et al., 2000; Wu, Zhu, and
Letchworth, 1998). The PrV gM has been shown to act as an inhibitor of cell fusion
resulting from cotransfection of either the PrV gB, gD and gH/gL or the F protein of
bovine respiratory syncytial virus, indicating a general mechanism of fusion inhibition by
gM (Klupp, Nixdorf, and Mettenleiter, 2000). While no definitive interaction has been
shown between the HSV-1 gM and UL49.5, an enhancement of function has been
demonstrated when both are present, inasmuch as cotransfection of the HSV-1 gM and
UL49.5 significantly inhibited cell fusion caused by the HSV-1 gB, gD and gH/gL or the
Molony murine leukemia virus envelope protein, while transfection of the HSV-1 gM
alone did not inhibit membrane fusion (Koyano et al., 2003). A requirement of both gM
and gN for inhibition of cell fusion was also shown for HHV-8 (Koyano et al., 2003). At
this point in time, the mechanism by which gM and UL49.5 mediate inhibition of cell-tocell fusion is unknown.
Characterization of the UL20 Protein (UL20p)
The UL20 ORF
The UL20 open reading frame is situated between the UL19 gene and the recently
confirmed UL20.5 gene in the unique long region of the HSV-1 genome. The UL20 gene
58
is expressed from the complementary strand of the viral DNA. The 669 bp UL20 gene
encodes the 222 aa UL20 protein (UL20p). UL20 is expressed as a γ1 (late) gene that
does not require viral DNA replication for expression (Ward et al., 1994) and is
conserved in the alphaherpesviruses.
Membrane Topology of UL20p
Based on its predicted amino acid sequence, the UL20 protein was suggested to
be an integral membrane protein containing two or three hydrophobic transmembrane
domains (McGeoch et al., 1988). However, when the hydrophobic and membranespanning domains of UL20p were calculated by using the TMPred and SOSUI computer
algorithms (Hirokawa, Boon-Chieng, and Mitaku, 1998; Hofmann and Stoffel, 1993) and
used to derive a predicted membrane spanning model of UL20p, four hydrophobic
regions were predicted, with no predicted signal peptide. This putative UL20p model
features four membrane spanning regions (Fig. 4.1), placing both the 66 aa amino
terminus and the 14 aa carboxyl terminus of UL20p within the cytoplasm (domains I and
V). In addition, a third small 10 aa domain is predicted to be located intracellularly
(domain III), while the two other domains are predicted to be located extracellularly
(domains II and IV). Domain II is predicted to contain 7 aa, while domain IV is
predicted to contain 32 aa. As the largest extracellular domain, domain IV holds the best
chance for interactions with the extracellular portions of other membrane proteins. This
predicted model of the UL20p membrane topology has been partially confirmed in our
laboratory with FLAG epitope tags located either on the N-terminus of UL20p, which is
intracellular, or inside of domain IV, which is present on the extracellular/lumenal side of
cellular membranes (Foster et al., 2004b).
59
Interdependence with gK for Cell Surface Expression and Internalization to the TransGolgi Network
Initial reports on the cellular localization of the UL20 protein indicated that it was
not expressed at the cell surface but was present in virions purified from extracellular
fluid and from the cytoplasm. The UL20 protein was found to be present in the nuclear
membranes, in the Golgi apparatus, and dispersed in the cytoplasm but was not detected
in the plasma membranes of infected cells (Ward et al., 1994). In contrast, recent
experiments performed by our laboratory showed that independently expressed gK or
UL20p failed to be transported from the ER in Vero cells. Similarly, infection of Vero
cells with either a gK-null or UL20-null virus resulted in ER entrapment of UL20p or gK,
respectively. In contrast, transiently coexpressed gK and UL20p predominantly localized
to the TGN, and TGN-localized gK and UL20p to originated from internalized gK and
UL20p expressed at cell surfaces. In all experiments, UL20p and gK colocalized,
consistent with the hypothesis that gK and UL20p directly interact, and that this
interaction is important for their TGN localization and function in the virus life cycle
(Foster et al., 2004b).
Function of UL20p in the HSV-1 Lifecycle
The original description of the role of UL20p in the virus life cycle was
predicated on the observation that an F-strain UL20-null virus, R7225, had a perinuclear
block in virus egress, resulting in a large accumulation of virions between the inner and
outer nuclear membrane as well as an abnormal level of unenveloped capsids in the
cytoplasm. In addition, the R7225 virus formed small syncytia on 143TK- cells (Baines
et al., 1991). However, at the time it was unknown that the UL20.5 gene was located
adjacent to UL20, and the R7225 virus contains an aberrantly fused gene consisting of
60
the 5’ half of the UL20.5 ORF and the 3’ half of the UL20 gene. As such, it was
unknown whether or not the description of the UL20-null phenotype could be attributed
to the aberrant gene.
To resolve the true phenotype of an HSV-1 virus lacking the UL20 gene, our
laboratory constructed an insertion-deletion mutation into the HSV-1 KOS-strain UL20
gene that consisted of removal of the 5’ region of the UL20 ORF, while maintaining the
integrity of both the UL20.5 gene and the promoter for the major capsid protein, UL19.
In accordance with the R7225 virus, the KOS strain UL20-null virus exhibited a defect in
virion egress; however, the block in virion egress consisted solely of an accumulation of
unenveloped capsids in the cytoplasm, and no accumulation of perinuclear virions was
observed (Foster et al., 2004a). Furthermore, the KOS UL20-null virus did not form
syncytial plaques on 143TK- cells. The observable phenotypes of the KOS UL20-null
virus were in agreement with the phenotype described for a PrV virus lacking UL20
(Fuchs et al., 1997), indicating that the previously described phenotype of the R7225
virus was in all likelihood due to the presence of the aberrantly fused gene. In addition,
we found that the UL20 protein was required for virus-induced cell-to-cell fusion
resulting from syncytial mutations in either gB or gK, implying that UL20 may serve to
regulate membrane fusion events in HSV-1 (Foster et al., 2004a). In Chapter IV, we
investigate the domains of UL20 that function in gB and/or gK-induced syncytia
formation as well as in virion egress.
Virus-Induced Syncytia Formation
While wild-type herpesviruses usually cause infected cells to round up and clump
together, some viral mutants in both HSV-1 and HSV-2 cause cells to fuse into large
61
polykaryocytes containing many nuclei (Ejercito, Kieff, and Roizman, 1968; Hoggan and
Roizman, 1959; Ruyechan et al., 1979). Virus-induced cell-to-cell fusion has been
extensively studied for several reasons: as a probe of the structure and function of
cellular membranes during infection reflected in their behavior, as a tool for analysis of
the functions of viral membrane proteins, and as a model of the initial interaction
between HSV and susceptible cells that results in fusion of the viral envelope with the
cellular plasma membrane (Campadelli-Fiume and Serafini-Cessi, 1985; Roizman, 1962;
Spear, 1985a; Spear, 1985b).
Syncytia formation can be viewed as an aberrant manifestation of the interactions
of altered membranes in herpesvirus infected cells with the unaltered membranes of
neighboring cells. Genetic analysis has shown that mutations that cause extensive virusinduced cell-to-cell fusion map to at least four and possible more loci within the viral
genome: the UL20 gene (Baines et al., 1991; MacLean et al., 1991; Melancon, Foster,
and Kousoulas, 2004), the UL24 gene (Jacobson, Martin, and Coen, 1989; Sanders,
Wilkie, and Davison, 1982), the UL27 gene encoding glycoprotein B (gB) (Bzik et al.,
1984; Pellett et al., 1985), and the UL53 gene coding for glycoprotein K (gK) (Bond and
Person, 1984; Debroy, Pederson, and Person, 1985; Hutchinson, Roop-Beauchamp, and
Johnson, 1995; Pogue-Geile et al., 1984; Pogue-Geile and Spear, 1987; Ruyechan et al.,
1979), with the great majority of syncytial mutations arising in either UL27 (gB) or UL53
(gK).
The mechanisms involved in virus-induced cell fusion are unclear, but some
progess has been made on understanding the components required for cell fusion
resulting from syncytial mutations in UL27 or UL53. In one set of experiments, the
62
requirements for syncytia formation caused by a mutation at codon 855 of gB (gBANG)
were analyzed. In Vero cells, gC, gG, gJ, and UL43 were shown to be dispensable for
syncytium formation at both high and low multiplicities of infection, while gD, gH, gL,
gE, gI, and gM were all required for the fusion of cellular membranes (Davis-Poynter et
al., 1994). These results confirmed that the requirements for virion entry and cell-to-cell
fusion are not identical. While gD and gH/gL, like gB, are essential for both virion entry
and virus-induced cell-to-cell fusion, gE, gI, and gM, on the other hand, are dispensable
for virus penetration, yet can play a role in cell-to-cell spread (Davis-Poynter et al.,
1994). In addition, it has been shown that syncytial mutations in gK failed to cause virusinduced cell fusion in the absence of gB, indicating that gB was essential for gKassociated cell-to-cell fusion (Cai, Gu, and Person, 1988). Recently, our laboratory
demonstrated a requirement for UL20 in virus-induced cell fusion caused by syncytial
mutations in either gB or gK. However, the role of gK in syncytia formation caused by
gB syncytial mutations had not been studied, and in Chapter III we investigate this
possibility.
REFERENCES
Adams, H. G., Benson, E. A., Alexander, E. R., Vontver, L. A., Remington, M. A., and
Holmes, K. K. (1976). Genital herpetic infection in men and women: clinical
course and effect of topical application of adenine arabinoside. J Infect Dis 133
Suppl, A151-9.
Adams, R., Cunningham, C., Davison, M. D., MacLean, C. A., and Davison, A. J. (1998).
Characterization of the protein encoded by gene UL49A of herpes simplex virus
type 1. J Gen Virol 79 ( Pt 4), 813-23.
Alconada, A., Bauer, U., Sodeik, B., and Hoflack, B. (1999). Intracellular traffic of
herpes simplex virus glycoprotein gE: characterization of the sorting signals
required for its trans-Golgi network localization. J Virol 73(1), 377-87.
Allen, W. P., and Rapp, F. (1982). Concept review of genital herpes vaccines. J Infect
Dis 145(3), 413-21.
63
Alwine, J. C., Steinhart, W. L., and Hill, C. W. (1974). Transcription of herpes simplex
type 1 DNA in nuclei isolated from infected HEp-2 and KB cells. Virology 60(1),
302-7.
Amir, J., Harel, L., Smetana, Z., and Varsano, I. (1997). Treatment of herpes simplex
gingivostomatitis with aciclovir in children: a randomised double blind placebo
controlled study. Bmj 314(7097), 1800-3.
Armstrong, J. A., Pereira, H. G., and Andrewes, C. H. (1961). Observations on the virus
of infectious bovine rhinotracheitis, and its affinity with the Herpesvirus group.
Virology 14, 276-85.
Astruc, J. (1736). "De Morbis Venereis Libri Sex." G. Cavelier, Paris.
Atkinson, M. A., Barr, S., and Timbury, M. C. (1978). The fine structure of cells infected
with temperature-sensitive mutants of herpes simplex virus type 2. J. Gen. Virol.
40, 103-119.
Babic, N., Klupp, B., Brack, A., Mettenleiter, T. C., Ugolini, G., and Flamand, A. (1996).
Deletion of glycoprotein gE reduces the propagation of pseudorabies virus in the
nervous system of mice after intranasal inoculation. Virology 219(1), 279-84.
Bader, C., Crumpacker, C. S., Schnipper, L. E., Ransil, B., Clark, J. E., Arndt, K., and
Freedberg, I. M. (1978). The natural history of recurrent facial-oral infection with
herpes simplex virus. J Infect Dis 138(6), 897-905.
Baghian, A., Huang, L., Newman, S., Jayachandra, S., and Kousoulas, K. G. (1993).
Truncation of the carboxy-terminal 28 amino acids of glycoprotein B specified by
herpes simplex virus type 1 mutant amb1511-7 causes extensive cell fusion. J.
Virol. 67(4), 2396-401.
Baines, J. D., and Roizman, B. (1992). The UL11 gene of herpes simplex virus 1 encodes
a function that facilitates nucleocapsid envelopment and egress from cells. J.
Virol. 66(8), 5168-74.
Baines, J. D., and Roizman, B. (1993). The UL10 gene of herpes simplex virus 1 encodes
a novel viral glycoprotein, gM, which is present in the virion and in the plasma
membrane of infected cells. J. Virol. 67(3), 1441-52.
Baines, J. D., Ward, P. L., Campadelli-Fiume, G., and Roizman, B. (1991). The UL20
gene of herpes simplex virus 1 encodes a function necessary for viral egress. J.
Virol. 65(12), 6414-24.
Balan, P., Davis-Poynter, N., Bell, S., Atkinson, H., Browne, H., and Minson, T. (1994).
An analysis of the in vitro and in vivo phenotypes of mutants of herpes simplex
virus type 1 lacking glycoproteins gG, gE, gI or the putative gJ. J. Gen. Virol.
75(Pt 6), 1245-58.
64
Banfield, B. W., Leduc, Y., Esford, L., Visalli, R. J., Brandt, C. R., and Tufaro, F. (1995).
Evidence for an interaction of herpes simplex virus with chondroitin sulfate
proteoglycans during infection. Virology 208(2), 531-9.
Baringer, J. R., and Swoveland, P. (1973). Recovery of herpes-simplex virus from human
trigeminal ganglions. N Engl J Med 288(13), 648-50.
Barker, D. E., and Roizman, B. (1992). The unique sequence of the herpes simplex virus
1 L component contains an additional translated open reading frame designated
UL49.5. J Virol 66(1), 562-6.
Barnett, B. C., Dolan, A., Telford, E. A., Davison, A. J., and McGeoch, D. J. (1992). A
novel herpes simplex virus gene (UL49A) encodes a putative membrane protein
with counterparts in other herpesviruses. J Gen Virol 73 ( Pt 8), 2167-71.
Bartoletti, A. M., Tognon, M., Manservigi, R., and Mannini-Palenzona, A. (1985).
Characterization of virus obtained from MDBK cells persistently infected with a
variant of herpes simplex virus type 1 strain MP [HSV-1(MP)]. Virology 141(2),
306-10.
Bastian, F. O., Rabson, A. S., Yee, C. L., and Tralka, T. S. (1972). Herpesvirus hominis:
isolation from human trigeminal ganglion. Science 178(58), 306-7.
Bateman, T. (1814). "A Practical Synopsis of Cutaneous Diseases According to the
Arrangement of Dr. Willan." Longman, Hurst, Orme, and Brown, London.
Batterson, W., Furlong, D., and Roizman, B. (1983). Molecular genetics of herpes
simplex virus. VIII. further characterization of a temperature-sensitive mutant
defective in release of viral DNA and in other stages of the viral reproductive
cycle. J Virol 45(1), 397-407.
Batterson, W., and Roizman, B. (1983). Characterization of the herpes simplex virionassociated factor responsible for the induction of alpha genes. J. Virol. 46(2), 3717.
Becker, Y., Dym, H., and Sarov, I. (1968). Herpes simplex virus DNA. Virology 36(2),
184-92.
Binder, P. S. (1977). Herpes simplex keratitis. Surv Ophthalmol 21(4), 313-31.
Black, F. L. (1975). Infectious diseases in primitive societies. Science 187(4176), 515-8.
Black, F. L., Hierholzer, W. J., Pinheiro, F., Evans, A. S., Woodall, J. P., Opton, E. M.,
Emmons, J. E., West, B. S., Edsall, G., Downs, W. G., and Wallace, G. D. (1974).
Evidence for persistence of infectious agents in isolated human populations. Am J
Epidemiol 100(3), 230-50.
65
Bond, V. C., and Person, S. (1984). Fine structure physical map locations of alterations
that affect cell fusion in herpes simplex virus type 1. Virology 132(2), 368-76.
Bond, V. C., Person, S., and Warner, S. C. (1982). The isolation and characterization of
mutants of herpes simplex virus type 1 that induce cell fusion. J. Gen. Virol. 61
(Pt 2), 245-54.
Brack, A. R., Dijkstra, J. M., Granzow, H., Klupp, B. G., and Mettenleiter, T. C. (1999).
Inhibition of virion maturation by simultaneous deletion of glycoproteins E, I, and
M of pseudorabies virus. J Virol 73(7), 5364-72.
Brown, Z. A., Benedetti, J., Ashley, R., Burchett, S., Selke, S., Berry, S., Vontver, L. A.,
and Corey, L. (1991). Neonatal herpes simplex virus infection in relation to
asymptomatic maternal infection at the time of labor. N Engl J Med 324(18),
1247-52.
Browne, H., Bell, S., and Minson, T. (2004). Analysis of the requirement for glycoprotein
m in herpes simplex virus type 1 morphogenesis. J Virol 78(2), 1039-41.
Buddingh, G. J., Schrum, D. I., Lanier, J. C., and Guidry, D. J. (1953). Studies of the
natural history of herpes simplex infections. Pediatrics 11(6), 595-610.
Burnet, F. M., and Williams, S. W. (1939). Herpes simplex: new point of view. Med J
Aust 1, 637-640.
Bzik, D. J., Fox, B. A., DeLuca, N. A., and Person, S. (1984). Nucleotide sequence of a
region of the herpes simplex virus type 1 gB glycoprotein gene: mutations
affecting rate of virus entry and cell fusion. Virology 137(1), 185-90.
Cai, W., Person, S., Warner, S. C., Zhou, J., and DeLuca, N. A. (1987). Linker-insertion
nonsense and restriction-site deletion mutations of the gB glycoprotein gene of
herpes simplex virus type 1. J. Virol. 61, 714-721.
Cai, W. H., Gu, B., and Person, S. (1988). Role of glycoprotein B of herpes simplex virus
type 1 in viral entry and cell fusion. J. Virol. 62(8), 2596-604.
Cai, W. Z., Person, S., DebRoy, C., and Gu, B. H. (1988). Functional regions and
structural features of the gB glycoprotein of herpes simplex virus type 1. An
analysis of linker insertion mutants. J. Mol. Biol. 201(3), 575-88.
Campadelli-Fiume, G., Arsenakis, M., Farabegoli, F., and Roizman, B. (1988). Entry of
herpes simplex virus 1 in BJ cells that constitutively express viral glycoprotein D
is by endocytosis and results in degradation of the virus. J. Virol. 62(1), 159-67.
Campadelli-Fiume, G., Farabegoli, F., Di, G. S., and Roizman, B. (1991). Origin of
unenveloped capsids in the cytoplasm of cells infected with herpes simplex virus
1. J. Virol. 65(3), 1589-95.
66
Campadelli-Fiume, G., and Serafini-Cessi, F. (1985). Processing of the oligosaccharide
chains of herpes simplex virus type 1 glycoproteins. In "The herpesviruses" (B.
Roizman, Ed.), Vol. 3, pp. 357-382. Plenum, New York.
Campbell, M. E., Palfreyman, J. W., and Preston, C. M. (1984). Identification of herpes
simplex virus DNA sequences which encode a trans-acting polypeptide
responsible for stimulation of immediate early transcription. J Mol Biol 180(1), 119.
Card, J. P., Whealy, M. E., Robbins, A. K., and Enquist, L. W. (1992). Pseudorabies
virus envelope glycoprotein gI influences both neurotropism and virulence during
infection of the rat visual system. J Virol 66(5), 3032-41.
Carfi, A., Willis, S. H., Whitbeck, J. C., Krummenacher, C., Cohen, G. H., Eisenberg, R.
J., and Wiley, D. C. (2001). Herpes simplex virus glycoprotein D bound to the
human receptor HveA. Mol Cell 8(1), 169-79.
Challberg, M. D. (1986). A method for identifying the viral genes required for
herpesvirus DNA replication. Proc Natl Acad Sci U S A 83(23), 9094-8.
Chang, Y., Cesarman, E., Pessin, M. S., Lee, F., Culpepper, J., Knowles, D. M., and
Moore, P. S. (1994). Identification of herpesvirus-like DNA sequences in AIDSassociated Kaposi's sarcoma. Science 266(5192), 1865-9.
Chang, Y. E., Menotti, L., Filatov, F., Campadelli-Fiume, G., and Roizman, B. (1998).
UL27.5 is a novel gamma2 gene antisense to the herpes simplex virus 1 gene
encoding glycoprotein B. J Virol 72(7), 6056-64.
Chang, Y. E., Van Sant, C., Krug, P. W., Sears, A. E., and Roizman, B. (1997). The null
mutant of the U(L)31 gene of herpes simplex virus 1: construction and phenotype
in infected cells. J Virol 71(11), 8307-15.
Chou, J., and Roizman, B. (1986). The terminal a sequence of the herpes simplex virus
genome contains the promoter of a gene located in the repeat sequences of the L
component. J Virol 57(2), 629-37.
Church, G. A., and Wilson, D. W. (1997). Study of herpes simplex virus maturation
during a synchronous wave of assembly. J Virol 71(5), 3603-12.
Claesson-Welsh, L., and Spear, P. G. (1986). Oligomerization of herpes simplex virus
glycoprotein B. J. Virol. 60, 803-806.
Claesson-Welsh, L., and Spear, P. G. (1987). Amino-terminal sequence, synthesis, and
membrane insertion of glycoprotein B of herpes simplex virus type 1. J. Virol. 61,
1-7.
67
Clements, J. B., Watson, R. J., and Wilkie, N. M. (1977). Temporal regulation of herpes
simplex virus type 1 transcription: location of transcripts on the viral genome.
Cell 12(1), 275-85.
Cocchi, F., Menotti, L., Di Ninni, V., Lopez, M., and Campadelli-Fiume, G. (2004). The
herpes simplex virus JMP mutant enters receptor-negative J cells through a novel
pathway independent of the known receptors nectin1, HveA, and nectin2. J Virol
78(9), 4720-9.
Cocchi, F., Menotti, L., Mirandola, P., Lopez, M., and Campadelli-Fiume, G. (1998). The
ectodomain of a novel member of the immunoglobulin subfamily related to the
poliovirus receptor has the attributes of a bona fide receptor for herpes simplex
virus types 1 and 2 in human cells. J Virol 72(12), 9992-10002.
Cohen, J. I., and Nguyen, H. (1997). Varicella-zoster virus glycoprotein I is essential for
growth of virus in Vero cells. J Virol 71(9), 6913-20.
Conley, A. J., Knipe, D. M., Jones, P. C., and Roizman, B. (1981). Molecular genetics of
herpes simplex virus. VII. Characterization of a temperature-sensitive mutant
produced by in vitro mutagenesis and defective in DNA synthesis and
accumulation of gamma polypeptides. J Virol 37(1), 191-206.
Corey, L. (1982). The diagnosis and treatment of genital herpes. Jama 248(9), 1041-9.
Corey, L., Adams, H. G., Brown, Z. A., and Holmes, K. K. (1983). Genital herpes
simplex virus infections: clinical manifestations, course, and complications. Ann
Intern Med 98(6), 958-72.
Corey, L., Holmes, K., Benedetti, J., and Critchlow, C. (1981). Clinical course of genital
herpes: Implications for therapeutic trials. In "The human herpesviruses, an
interdisciplinary perspective" (W. R. Dowdle, A. J. Nahmias, and R. F. Schinazi,
Eds.), pp. 496-502. Elsevier, Amsterdam.
Costanzo, F., Campadelli-Fiume, G., Foa-Tomasi, L., and Cassai, E. (1977). Evidence
that herpes simplex virus DNA is transcribed by cellular RNA polymerase B. J
Virol 21(3), 996-1001.
Craig, J. M., Macauley, J. C., Weller, T. H., and Wirth, P. (1957). Isolation of
intranuclear inclusion producing agents from infants with illnesses resembling
cytomegalic inclusion disease. Proc Soc Exp Biol Med 94(1), 4-12.
Darlington, R. W., and Moss, L. H. D. (1968). Herpesvirus envelopment. J. Virol. 2(1),
48-55.
Davison, M. D., Rixon, F. J., and Davison, A. J. (1992). Identification of genes encoding
two capsid proteins (VP24 and VP26) of herpes simplex virus type 1. J Gen Virol
73 ( Pt 10), 2709-13.
68
Davis-Poynter, N., Bell, S., Minson, T., and Browne, H. (1994). Analysis of the
contributions of herpes simplex virus type 1 membrane proteins to the induction
of cell-cell fusion. J. Virol. 68(11), 7586-90.
Deb, S., and Doelberg, M. (1988). A 67-base-pair segment from the Ori-S region of
herpes simplex virus type 1 encodes origin function. J Virol 62(7), 2516-9.
Debroy, C., Pederson, N., and Person, S. (1985). Nucleotide sequence of a herpes
simplex virus type 1 gene that causes cell fusion. Virology 145(1), 36-48.
Deiss, L. P., Chou, J., and Frenkel, N. (1986). Functional domains within the a sequence
involved in the cleavage-packaging of herpes simplex virus DNA. J Virol 59(3),
605-18.
Desai, P., Sexton, G. L., McCaffery, J. M., and Person, S. (2001). A null mutation in the
gene encoding the herpes simplex virus type 1 UL37 polypeptide abrogates virus
maturation. J Virol 75(21), 10259-71.
Desai, P. J. (2000). A null mutation in the UL36 gene of herpes simplex virus type 1
results in accumulation of unenveloped DNA-filled capsids in the cytoplasm of
infected cells. J Virol 74(24), 11608-18.
Dietz, P., Klupp, B. G., Fuchs, W., Kollner, B., Weiland, E., and Mettenleiter, T. C.
(2000). Pseudorabies virus glycoprotein K requires the UL20 gene product for
processing. J Virol 74(11), 5083-90.
Dingwell, K. S., Brunetti, C. R., Hendricks, R. L., Tang, Q., Tang, M., Rainbow, A. J.,
and Johnson, D. C. (1994). Herpes simplex virus glycoproteins E and I facilitate
cell-to-cell spread in vivo and across junctions of cultured cells. J. Virol. 68(2),
834-45.
Dingwell, K. S., Doering, L. C., and Johnson, D. C. (1995). Glycoproteins E and I
facilitate neuron-to-neuron spread of herpes simplex virus. J. Virol. 69(11), 708798.
Dingwell, K. S., and Johnson, D. C. (1998). The herpes simplex virus gE-gI complex
facilitates cell-to-cell spread and binds to components of cell junctions. J Virol
72(11), 8933-42.
Dixon, R. A., and Schaffer, P. A. (1980). Fine-structure mapping and functional analysis
of temperature-sensitive mutants in the gene encoding the herpes simplex virus
type 1 immediate early protein VP175. J Virol 36(1), 189-203.
Dubin, G., and Jiang, H. (1995). Expression of herpes simplex virus type 1 glycoprotein
L (gL) in transfected mammalian cells: evidence that gL is not independently
anchored to cell membranes. J Virol 69(7), 4564-8.
69
Eberle, F., Dubreuil, P., Mattei, M. G., Devilard, E., and Lopez, M. (1995). The human
PRR2 gene, related to the human poliovirus receptor gene (PVR), is the true
homolog of the murine MPH gene. Gene 159(2), 267-72.
Eisenberg, R. J., Ponce de Leon, M., Friedman, H. M., Fries, L. F., Frank, M. M.,
Hastings, J. C., and Cohen, G. H. (1987). Complement component C3b binds
directly to purified glycoprotein C of herpes simplex virus types 1 and 2. Microb
Pathog 3(6), 423-35.
Ejercito, P. M., Kieff, E. D., and Roizman, B. (1968). Characterization of herpes simplex
virus strains differing in their effects on social behavior of infected cells. J. Gen.
Virol. 53, 634-644.
Elgadi, M. M., and Smiley, J. R. (1999). Picornavirus internal ribosome entry site
elements target RNA cleavage events induced by the herpes simplex virus virion
host shutoff protein. J Virol 73(11), 9222-31.
Enquist, L. W., Husak, P. J., Banfield, B. W., and Smith, G. A. (1998). Infection and
spread of alphaherpesviruses in the nervous system. Adv Virus Res 51, 237-347.
Epstein, M. A. (1962). Observations on the mode of release of herpes virus from infected
HeLa cells. J Cell Biol 12, 589-97.
Epstein, M. A., Achong, B. G., and Barr, Y. M. (1964). Virus Particles in Cultured
Lymphoblasts from Burkitt's Lymphoma. Lancet 15, 702-3.
Everett, R. D., Orr, A., and Elliott, M. (1991). High level expression and purification of
herpes simplex virus type 1 immediate early polypeptide Vmw110. Nucleic Acids
Res 19(22), 6155-61.
Faber, S. W., and Wilcox, K. W. (1986). Association of the herpes simplex virus
regulatory protein ICP4 with specific nucleotide sequences in DNA. Nucleic
Acids Res 14(15), 6067-83.
Falke, D., Siegert, R., and Vogell, W. (1959). [Electron microscopic findings on the
problem of double membrane formation in herpes simplex virus]. Arch Gesamte
Virusforsch 9, 484-96.
Farnsworth, A., Goldsmith, K., and Johnson, D. C. (2003). Herpes simplex virus
glycoproteins gD and gE/gI serve essential but redundant functions during
acquisition of the virion envelope in the cytoplasm. J Virol 77(15), 8481-94.
Fenwick, M., Morse, L. S., and Roizman, B. (1979). Anatomy of herpes simplex virus
DNA. XI. Apparent clustering of functions effecting rapid inhibition of host DNA
and protein synthesis. J Virol 29(2), 825-7.
Fenwick, M. L., and Walker, M. J. (1978). Suppression of the synthesis of cellular
macromolecules by herpes simplex virus. J Gen Virol 41(1), 37-51.
70
Fleming, D. T., McQuillan, G. M., Johnson, R. E., Nahmias, A. J., Aral, S. O., Lee, F. K.,
and St Louis, M. E. (1997). Herpes simplex virus type 2 in the United States,
1976 to 1994. N Engl J Med 337(16), 1105-11.
Foa-Tomasi, L., Avitabile, E., Boscaro, A., Brandimarti, R., Gualandri, R., Manservigi,
R., Dall'Olio, F., Serafini-Cessi, F., and Fiume, G. C. (1991). Herpes simplex
virus (HSV) glycoprotein H is partially processed in a cell line that expresses the
glycoprotein and fully processed in cells infected with deletion or ts mutants in
the known HSV glycoproteins. Virology 180(2), 474-82.
Forrester, A., Farrell, H., Wilkinson, G., Kaye, J., Davis-Poynter, N., and Minson, T.
(1992). Construction and properties of a mutant of herpes simplex virus type 1
with glycoprotein H coding sequences deleted. J Virol 66(1), 341-8.
Forrester, A. J., Sullivan, V., Simmons, A., Blacklaws, B. A., Smith, G. L., Nash, A. A.,
and Minson, A. C. (1991). Induction of protective immunity with antibody to
herpes simplex virus type 1 glycoprotein H (gH) and analysis of the immune
response to gH expressed in recombinant vaccinia virus. J Gen Virol 72 ( Pt 2),
369-75.
Foster, T. P., Alvarez, X., and Kousoulas, K. G. (2003). Plasma membrane topology of
syncytial domains of herpes simplex virus type 1 glycoprotein K (gK): the UL20
protein enables cell surface localization of gK but not gK-mediated cell-to-cell
fusion. J Virol 77(1), 499-510.
Foster, T. P., Melancon, J. M., Baines, J. D., and Kousoulas, K. G. (2004a). The Herpes
Simplex Virus Type 1 UL20 Protein Modulates Membrane Fusion Events during
Cytoplasmic Virion Morphogenesis and Virus-Induced Cell Fusion. J Virol
78(10), 5347-57.
Foster, T. P., Melancon, J. M., and Kousoulas, K. G. (2001). An alpha-helical domain
within the carboxyl terminus of herpes simplex virus type 1 (HSV-1) glycoprotein
B (gB) is associated with cell fusion and resistance to heparin inhibition of cell
fusion. Virology 287(1), 18-29.
Foster, T. P., Melancon, J. M., Olivier, T. L., and Kousoulas, K. G. (2004b). Herpes
simplex virus type-1 (HSV-1) glycoprotein K (gK) and the UL20 protein are
interdependent for intracellular trafficking and trans-Golgi network localization. J
Virol, (In Press).
Frame, M. C., Marsden, H. S., and McGeoch, D. J. (1986). Novel herpes simplex virus
type 1 glycoproteins identified by antiserum against a synthetic oligopeptide from
the predicted product of gene US4. J Gen Virol 67 ( Pt 4), 745-51.
Frenkel, N., Locker, H., Batterson, W., Hayward, G. S., and Roizman, B. (1976).
Anatomy of herpes simplex virus DNA. VI. Defective DNA originates from the S
component. J Virol 20(2), 527-31.
71
Frenkel, N., Schirmer, E. C., Wyatt, L. S., Katsafanas, G., Roffman, E., Danovich, R. M.,
and June, C. H. (1990). Isolation of a new herpesvirus from human CD4+ T cells.
Proc Natl Acad Sci U S A 87(2), 748-52.
Friedman, H. M., Cohen, G. H., Eisenberg, R. J., Seidel, C. A., and Cines, D. B. (1984).
Glycoprotein C of herpes simplex virus 1 acts as a receptor for the C3b
complement component on infected cells. Nature 309(5969), 633-5.
Friedman, H. M., Wang, L., Fishman, N. O., Lambris, J. D., Eisenberg, R. J., Cohen, G.
H., and Lubinski, J. (1996). Immune evasion properties of herpes simplex virus
type 1 glycoprotein gC. J Virol 70(7), 4253-60.
Frink, R. J., Anderson, K. P., and Wagner, E. K. (1981). Herpes simplex virus type 1
HindIII fragment L encodes spliced and complementary mRNA species. J Virol
39(2), 559-72.
Frink, R. J., Eisenberg, R., Cohen, G., and Wagner, E. K. (1983). Detailed analysis of the
portion of the herpes simplex virus type 1 genome encoding glycoprotein C. J
Virol 45(2), 634-47.
Fuchs, W., Klupp, B. G., Granzow, H., and Mettenleiter, T. C. (1997). The UL20 gene
product of pseudorabies virus functions in virus egress. J Virol 71(7), 5639-46.
Fuchs, W., Klupp, B. G., Granzow, H., Osterrieder, N., and Mettenleiter, T. C. (2002).
The interacting UL31 and UL34 gene products of pseudorabies virus are involved
in egress from the host-cell nucleus and represent components of primary
enveloped but not mature virions. J Virol 76(1), 364-78.
Furlong, D., Swift, H., and Roizman, B. (1972). Arrangement of herpesvirus
deoxyribonucleic acid in the core. J Virol 10(5), 1071-4.
Gage, P. J., Levine, M., and Glorioso, J. C. (1993). Syncytium-inducing mutations
localize to two discrete regions within the cytoplasmic domain of herpes simplex
virus type 1 glycoprotein B. J. Virol. 67(4), 2191-201.
Gelman, I. H., and Silverstein, S. (1987). Herpes simplex virus immediate-early
promoters are responsive to virus and cell trans-acting factors. J Virol 61(7),
2286-96.
Geraghty, R. J., Krummenacher, C., Cohen, G. H., Eisenberg, R. J., and Spear, P. G.
(1998). Entry of alphaherpesviruses mediated by poliovirus receptor-related
protein 1 and poliovirus receptor. Science 280(5369), 1618-20.
Gershon, A. A., Sherman, D. L., Zhu, Z., Gabel, C. A., Ambron, R. T., and Gershon, M.
D. (1994). Intracellular transport of newly synthesized varicella-zoster virus: final
envelopment in the trans-Golgi network. J Virol 68(10), 6372-90.
72
Ghiasi, H., Nesburn, A. B., Cai, S., and Wechsler, S. L. (1998). The US5 open reading
frame of herpes simplex virus type 1 does encode a glycoprotein (gJ).
Intervirology 41(2-3), 91-7.
Gibson, W. (1996). Structure and assembly of the virion. Intervirology 39(5-6), 389-400.
Gibson, W., and Roizman, B. (1972). Proteins specified by herpes simplex virus. VIII.
Characterization and composition of multiple capsid forms of subtypes 1 and 2. J
Virol 10(5), 1044-52.
Godowski, P. J., and Knipe, D. M. (1986). Transcriptional control of herpesvirus gene
expression: gene functions required for positive and negative regulation. Proc
Natl Acad Sci U S A 83(2), 256-60.
Goodman, J. L., and Engel, J. P. (1991). Altered pathogenesis in herpes simplex virus
type 1 infection due to a syncytial mutation mapping to the carboxy terminus of
glycoprotein B. J. Virol. 65(4), 1770-8.
Granzow, H., Klupp, B. G., Fuchs, W., Veits, J., Osterrieder, N., and Mettenleiter, T. C.
(2001). Egress of alphaherpesviruses: comparative ultrastructural study. J Virol
75(8), 3675-84.
Gruter, W. (1924). Das herpesvirus, seine atiologischeund ung klinische bedeutung.
Munch Med Wschr 71, 1058.
Haan, K. M., Lee, S. K., and Longnecker, R. (2001). Different functional domains in the
cytoplasmic tail of glycoprotein B are involved in Epstein-Barr virus-induced
membrane fusion. Virology 290(1), 106-14.
Harms, J. S., Ren, X., Oliveira, S. C., and Splitter, G. A. (2000). Distinctions between
bovine herpesvirus 1 and herpes simplex virus type 1 VP22 tegument protein
subcellular associations. J Virol 74(7), 3301-12.
Harris, S. L., Frank, I., Yee, A., Cohen, G. H., Eisenberg, R. J., and Friedman, H. M.
(1990). Glycoprotein C of herpes simplex virus type 1 prevents complementmediated cell lysis and virus neutralization. J Infect Dis 162(2), 331-7.
Heine, J. W., Honess, R. W., Cassai, E., and Roizman, B. (1974). Proteins specified by
herpes simplex virus. XII. The virion polypeptides of type 1 strains. J Virol 14(3),
640-51.
Herold, B. C., Visalli, R. J., Susmarski, N., Brandt, C. R., and Spear, P. G. (1994).
Glycoprotein C-independent binding of herpes simplex virus to cells requires cell
surface heparan sulphate and glycoprotein B. J. Gen. Virol. 75, 1211-22.
Highlander, S. L., Goins, W. F., Person, S., Holland, T. C., Levine, M., and Glorioso, J.
C. (1991). Oligomer formation of the gB glycoprotein of herpes simplex virus
type 1. J Virol 65(8), 4275-83.
73
Hirokawa, T., Boon-Chieng, S., and Mitaku, S. (1998). SOSUI: classification and
secondary structure prediction system for membrane proteins. Bionformatics 14,
378-379.
Hofmann, K., and Stoffel, W. (1993). TMbase-database of membrane spanning segments.
J. Biol. Chem. 377, 166.
Hoggan, M. D., and Roizman, B. (1959). The isolation and properties of a variant of
Herpes simplex producing multinucleated giant cells in monolayer cultures in the
presence of antibody. Am J Hyg 70, 208-19.
Homa, F. L., Purifoy, D. J., Glorioso, J. C., and Levine, M. (1986). Molecular basis of the
glycoprotein C-negative phenotypes of herpes simplex virus type 1 mutants
selected with a virus-neutralizing monoclonal antibody. J Virol 58(2), 281-9.
Honess, R. W., and Roizman, B. (1974). Regulation of herpesvirus macromolecular
synthesis. I. Cascade regulation of the synthesis of three groups of viral proteins. J
Virol 14(1), 8-19.
Huber, M. T., and Compton, T. (1998). The human cytomegalovirus UL74 gene encodes
the third component of the glycoprotein H-glycoprotein L-containing envelope
complex. J Virol 72(10), 8191-7.
Huemer, H. P., Larcher, C., van Drunen Littel-van den Hurk, S., and Babiuk, L. A.
(1993). Species selective interaction of Alphaherpesvirinae with the "unspecific"
immune system of the host. Archives Of Virology. 130(3-4), 353-64.
Huemer, H. P., Nowotny, N., Crabb, B. S., Meyer, H., and Hubert, P. H. (1995). gp13
(EHV-gC): a complement receptor induced by equine herpesviruses. Virus Res
37(2), 113-26.
Hutchinson, L., Browne, H., Wargent, V., Davis-Poynter, N., Primorac, S., Goldsmith,
K., Minson, A. C., and Johnson, D. C. (1992a). A novel herpes simplex virus
glycoprotein, gL, forms a complex with glycoprotein H (gH) and affects normal
folding and surface expression of gH. J. Virol. 66(4), 2240-50.
Hutchinson, L., Goldsmith, K., Snoddy, D., Ghosh, H., Graham, F. L., and Johnson, D.
C. (1992b). Identification and characterization of a novel herpes simplex virus
glycoprotein, gK, involved in cell fusion. J Virol 66(9), 5603-9.
Hutchinson, L., and Johnson, D. C. (1995). Herpes simplex virus glycoprotein K
promotes egress of virus particles. J. Virol. 69(9), 5401-13.
Hutchinson, L., Roop-Beauchamp, C., and Johnson, D. C. (1995). Herpes simplex virus
glycoprotein K is known to influence fusion of infected cells, yet is not on the cell
surface. J. Virol. 69(7), 4556-63.
74
Igarashi, K., Fawl, R., Roller, R. J., and Roizman, B. (1993). Construction and properties
of a recombinant herpes simplex virus 1 lacking both S-component origins of
DNA synthesis. J Virol 67(4), 2123-32.
Isaacs, S. N., Kotwal, G. J., and Moss, B. (1992). Vaccinia virus complement-control
protein prevents antibody-dependent complement-enhanced neutralization of
infectivity and contributes to virulence. Proc Natl Acad Sci U S A 89(2), 628-32.
Ishov, A. M., and Maul, G. G. (1996). The periphery of nuclear domain 10 (ND10) as site
of DNA virus deposition. J Cell Biol 134(4), 815-26.
Jacob, R. J., Morse, L. S., and Roizman, B. (1979). Anatomy of herpes simplex virus
DNA. XII. Accumulation of head-to-tail concatemers in nuclei of infected cells
and their role in the generation of the four isomeric arrangements of viral DNA. J
Virol 29(2), 448-57.
Jacobs, L., Mulder, W. A., Van Oirschot, J. T., Gielkens, A. L., and Kimman, T. G.
(1993). Deleting two amino acids in glycoprotein gI of pseudorabies virus
decreases virulence and neurotropism for pigs, but does not affect
immunogenicity. J Gen Virol 74 ( Pt 10), 2201-6.
Jacobson, J. G., Chen, S. H., Cook, W. J., Kramer, M. F., and Coen, D. M. (1998).
Importance of the herpes simplex virus UL24 gene for productive ganglionic
infection in mice. Virology 242(1), 161-9.
Jacobson, J. G., Martin, S. L., and Coen, D. M. (1989). A conserved open reading frame
that overlaps the herpes simplex virus thymidine kinase gene is important for viral
growth in cell culture. J. Virol. 63(4), 1839-43.
Jayachandra, S., Baghian, A., and Kousoulas, K. G. (1997). Herpes simplex virus type 1
glycoprotein K is not essential for infectious virus production in actively
replicating cells but is required for efficient envelopment and translocation of
infectious virions from the cytoplasm to the extracellular space. J. Virol. 71(7),
5012-24.
Johnson, D. C., Burke, R. L., and Gregory, T. (1990). Soluble forms of herpes simplex
virus glycoprotein D bind to a limited number of cell surface receptors and inhibit
virus entry into cells. J. Virol. 64(6), 2569-76.
Johnson, D. C., and Feenstra, V. (1987). Identification of a novel herpes simplex virus
type 1-induced glycoprotein which complexes with gE and binds
immunoglobulin. J. Virol. 61(7), 2208-16.
Johnson, D. C., Frame, M. C., Ligas, M. W., Cross, A. M., and Stow, N. D. (1988).
Herpes simplex virus immunoglobulin G Fc receptor activity depends on a
complex of two viral glycoproteins, gE and gI. J Virol 62(4), 1347-54.
75
Johnson, D. C., and Spear, P. G. (1982). Monensin inhibits the processing of herpes
simplex virus glycoproteins, their transport to the cell surface, and the egress of
virions from infected cells. J. Virol. 43(3), 1102-12.
Johnson, D. C., Webb, M., Wisner, T. W., and Brunetti, C. (2001). Herpes simplex virus
gE/gI sorts nascent virions to epithelial cell junctions, promoting virus spread. J
Virol 75(2), 821-33.
Jons, A., Dijkstra, J. M., and Mettenleiter, T. C. (1998). Glycoproteins M and N of
pseudorabies virus form a disulfide-linked complex. J Virol 72(1), 550-7.
Jons, A., Granzow, H., Kuchling, R., and Mettenleiter, T. C. (1996). The UL49.5 gene of
pseudorabies virus codes for an O-glycosylated structural protein of the viral
envelope. J Virol 70(2), 1237-41.
Kalinyak, J. E., Fleagle, G., and Docherty, J. J. (1977). Incidence and distribution of
herpes simplex virus types 1 and 2 from genital lesions in college women. J Med
Virol 1(3), 175-81.
Karr, B. M., and Read, G. S. (1999). The virion host shutoff function of herpes simplex
virus degrades the 5' end of a target mRNA before the 3' end. Virology 264(1),
195-204.
Katan, M., Haigh, A., Verrijzer, C. P., van der Vliet, P. C., and O'Hare, P. (1990).
Characterization of a cellular factor which interacts functionally with Oct-1 in the
assembly of a multicomponent transcription complex. Nucleic Acids Res 18(23),
6871-80.
Kaufman, R. H., Gardner, H. L., Rawls, W. E., Dixon, R. E., and Young, R. L. (1973).
Clinical features of herpes genitalis. Cancer Res 33(6), 1446-51.
Kieff, E. D., Bachenheimer, S. L., and Roizman, B. (1971). Size, composition, and
structure of the deoxyribonucleic acid of herpes simplex virus subtypes 1 and 2. J
Virol 8(2), 125-32.
Kimura, H., Wang, Y., Pesnicak, L., Cohen, J. I., Hooks, J. J., Straus, S. E., and
Williams, R. K. (1998). Recombinant varicella-zoster virus glycoproteins E and I:
immunologic responses and clearance of virus in a guinea pig model of chronic
uveitis. J Infect Dis 178(2), 310-7.
Klupp, B. G., Fuchs, W., Granzow, H., Nixdorf, R., and Mettenleiter, T. C. (2002).
Pseudorabies virus UL36 tegument protein physically interacts with the UL37
protein. J Virol 76(6), 3065-71.
Klupp, B. G., Granzow, H., and Mettenleiter, T. C. (2000). Primary envelopment of
pseudorabies virus at the nuclear membrane requires the UL34 gene product. J
Virol 74(21), 10063-73.
76
Klupp, B. G., Granzow, H., Mundt, E., and Mettenleiter, T. C. (2001). Pseudorabies virus
UL37 gene product is involved in secondary envelopment. J Virol 75(19), 892736.
Klupp, B. G., Nixdorf, R., and Mettenleiter, T. C. (2000). Pseudorabies virus
glycoprotein M inhibits membrane fusion. J Virol 74(15), 6760-8.
Knipe, D. M., Batterson, W., Nosal, C., Roizman, B., and Buchan, A. (1981). Molecular
genetics of herpes simplex virus. VI. Characterization of a temperature-sensitive
mutant defective in the expression of all early viral gene products. J Virol 38(2),
539-47.
Knipe, D. M., Ruyechan, W. T., and Roizman, B. (1979). Molecular genetics of herpes
simplex virus. III. Fine mapping of a genetic locus determining resistance to
phosphonoacetate by two methods of marker transfer. J Virol 29(2), 698-704.
Knipe, D. M., Senechek, D., Rice, S. A., and Smith, J. L. (1987). Stages in the nuclear
association of the herpes simplex virus transcriptional activator protein ICP4. J
Virol 61(2), 276-84.
Knopf, C. W. (1986). Nucleotide sequence of the DNA polymerase gene of herpes
simplex virus type 1 strain Angelotti. Nucleic Acids Res 14(20), 8225-6.
Kopp, M., Granzow, H., Fuchs, W., Klupp, B., and Mettenleiter, T. C. (2004).
Simultaneous deletion of pseudorabies virus tegument protein UL11 and
glycoprotein M severely impairs secondary envelopment. J Virol 78(6), 3024-34.
Korsager, B., Spencer, E. S., Mordhorst, C. H., and Andersen, H. K. (1975). Herpesvirus
hominis infections in renal transplant recipients. Scand J Infect Dis 7(1), 11-9.
Kotwal, G. J., Isaacs, S. N., McKenzie, R., Frank, M. M., and Moss, B. (1990). Inhibition
of the complement cascade by the major secretory protein of vaccinia virus.
Science 250(4982), 827-30.
Koyano, S., Mar, E. C., Stamey, F. R., and Inoue, N. (2003). Glycoproteins M and N of
human herpesvirus 8 form a complex and inhibit cell fusion. J Gen Virol 84(Pt 6),
1485-91.
Kristie, T. M., and Roizman, B. (1984). Separation of sequences defining basal
expression from those conferring alpha gene recognition within the regulatory
domains of herpes simplex virus 1 alpha genes. Proc Natl Acad Sci U S A 81(13),
4065-9.
Kristie, T. M., and Roizman, B. (1986). DNA-binding site of major regulatory protein
alpha 4 specifically associated with promoter-regulatory domains of alpha genes
of herpes simplex virus type 1. Proc Natl Acad Sci U S A 83(13), 4700-4.
77
Kristie, T. M., and Sharp, P. A. (1990). Interactions of the Oct-1 POU subdomains with
specific DNA sequences and with the HSV alpha-trans-activator protein. Genes
Dev 4(12B), 2383-96.
Kritas, S. K., Pensaert, M. B., and Mettenleiter, T. C. (1994). Role of envelope
glycoproteins gI, gp63 and gIII in the invasion and spread of Aujeszky's disease
virus in the olfactory nervous pathway of the pig. J Gen Virol 75 ( Pt 9), 2319-27.
Kudelova, M., Kostal, M., Cervenakova, L., Rajcani, J., and Kaerner, H. C. (1991).
Pathogenicity and latency competence for rabbits of the herpes simplex virus type
1 ANGpath gC and gE defective mutants. Acta Virol 35(5), 438-49.
Kwon, B., Kim, B. S., Cho, H. R., Park, J. E., and Kwon, B. S. (2003). Involvement of
tumor necrosis factor receptor superfamily(TNFRSF) members in the
pathogenesis of inflammatory diseases. Exp Mol Med 35(1), 8-16.
Kwong, A. D., and Frenkel, N. (1987). Herpes simplex virus-infected cells contain a
function(s) that destabilizes both host and viral mRNAs. Proc Natl Acad Sci U S
A 84(7), 1926-30.
Kwong, A. D., Kruper, J. A., and Frenkel, N. (1988). Herpes simplex virus virion host
shutoff function. J Virol 62(3), 912-21.
La Boissiere, S., Hughes, T., and O'Hare, P. (1999). HCF-dependent nuclear import of
VP16. Embo J 18(2), 480-9.
Ladin, B. F., Blankenship, M. L., and Ben-Porat, T. (1980). Replication of herpesvirus
DNA. V. Maturation of concatemeric DNA of pseudorabies virus to genome
length is related to capsid formation. J Virol 33(3), 1151-64.
Ladin, B. F., Ihara, S., Hampl, H., and Ben-Porat, T. (1982). Pathway of assembly of
herpesvirus capsids: an analysis using DNA+ temperature-sensitive mutants of
pseudorabies virus. Virology 116(2), 544-61.
Lake, C. M., Molesworth, S. J., and Hutt-Fletcher, L. M. (1998). The Epstein-Barr virus
(EBV) gN homolog BLRF1 encodes a 15-kilodalton glycoprotein that cannot be
authentically processed unless it is coexpressed with the EBV gM homolog
BBRF3. J Virol 72(7), 5559-64.
Lam, Q., Smibert, C. A., Koop, K. E., Lavery, C., Capone, J. P., Weinheimer, S. P., and
Smiley, J. R. (1996). Herpes simplex virus VP16 rescues viral mRNA from
destruction by the virion host shutoff function. Embo J 15(10), 2575-81.
Laquerre, S., Person, S., and Glorioso, J. C. (1996). Glycoprotein B of herpes simplex
virus type 1 oligomerizes through the intermolecular interaction of a 28-aminoacid domain. J Virol 70(3), 1640-50.
78
Lee, G. T., Para, M. F., and Spear, P. G. (1982). Location of the structural genes for
glycoproteins gD and gE and for other polypeptides in the S component of herpes
simplex virus type 1 DNA. J Virol 43(1), 41-9.
Lee, J. Y., Irmiere, A., and Gibson, W. (1988). Primate cytomegalovirus assembly:
evidence that DNA packaging occurs subsequent to B capsid assembly. Virology
167(1), 87-96.
Leopardi, R., Ward, P. L., Ogle, W. O., and Roizman, B. (1997). Association of herpes
simplex virus regulatory protein ICP22 with transcriptional complexes containing
EAP, ICP4, RNA polymerase II, and viral DNA requires posttranslational
modification by the U(L)13 proteinkinase. J Virol 71(2), 1133-9.
Ligas, M. W., and Johnson, D. C. (1988). A herpes simplex virus mutant in which
glycoprotein D sequences are replaced by beta-galactosidase sequences binds to
but is unable to penetrate into cells. J. Virol. 62(5), 1486-94.
Locker, H., Frenkel, N., and Halliburton, I. (1982). Structure and expression of class II
defective herpes simplex virus genomes encoding infected cell polypeptide
number 8. J Virol 43(2), 574-93.
Lockshon, D., and Galloway, D. A. (1986). Cloning and characterization of oriL2, a large
palindromic DNA replication origin of herpes simplex virus type 2. J Virol 58(2),
513-21.
Long, D., Wilcox, W. C., Abrams, W. R., Cohen, G. H., and Eisenberg, R. J. (1992).
Disulfide bond structure of glycoprotein D of herpes simplex virus types 1 and 2.
J Virol 66(11), 6668-85.
Longnecker, R., Chatterjee, S., Whitley, R. J., and Roizman, B. (1987). Identification of a
herpes simplex virus 1 glycoprotein gene within a gene cluster dispensable for
growth in cell culture. Proc. Natl. Acad. Sci. USA. 84(12), 4303-7.
Lopez, C., Pellett, P., Stewart, J., Goldsmith, C., Sanderlin, K., Black, J., Warfield, D.,
and Feorino, P. (1988). Characteristics of human herpesvirus-6. J Infect Dis
157(6), 1271-3.
Lopez, M., Cocchi, F., Menotti, L., Avitabile, E., Dubreuil, P., and Campadelli-Fiume, G.
(2000). Nectin2alpha (PRR2alpha or HveB) and nectin2delta are low-efficiency
mediators for entry of herpes simplex virus mutants carrying the Leu25Pro
substitution in glycoprotein D. J Virol 74(3), 1267-74.
Lubinski, J., Wang, L., Mastellos, D., Sahu, A., Lambris, J. D., and Friedman, H. M.
(1999). In vivo role of complement-interacting domains of herpes simplex virus
type 1 glycoprotein gC. J Exp Med 190(11), 1637-46.
Lubinski, J. M., Wang, L., Soulika, A. M., Burger, R., Wetsel, R. A., Colten, H., Cohen,
G. H., Eisenberg, R. J., Lambris, J. D., and Friedman, H. M. (1998). Herpes
79
simplex virus type 1 glycoprotein gC mediates immune evasion in vivo. J Virol
72(10), 8257-63.
Mach, M., Kropff, B., Dal Monte, P., and Britt, W. (2000). Complex formation by human
cytomegalovirus glycoproteins M (gpUL100) and N (gpUL73). J Virol 74(24),
11881-92.
Machtiger, N. A., Pancake, B. A., Eberle, R., Courtney, R. J., Tevethia, S. S., and
Schaffer, P. A. (1980). Herpes simplex virus glycoproteins: isolation of mutants
resistant to immune cytolysis. J. Virol. 34, 336-346.
MacLean, C. A., Dolan, A., Jamieson, F. E., and McGeoch, D. J. (1992). The
myristylated virion proteins of herpes simplex virus type 1: investigation of their
role in the virus life cycle. J. Gen. Virol. 73(Pt 3), 539-47.
MacLean, C. A., Efstathiou, S., Elliott, M. L., Jamieson, F. E., and McGeoch, D. J.
(1991). Investigation of herpes simplex virus type 1 genes encoding multiply
inserted membrane proteins. J. Gen. Virol. 72(Pt 4), 897-906.
MacLean, C. A., Robertson, L. M., and Jamieson, F. E. (1993). Characterization of the
UL10 gene product of herpes simplex virus type 1 and investigation of its role in
vivo. J Gen Virol 74 ( Pt 6), 975-83.
Manservigi, R., Spear, P. G., and Buchan, A. (1977). Cell fusion induced by herpes
simplex virus is promoted and suppressed by different viral glycoproteins. Proc.
Natl. Acad. Sci. USA. 74(9), 3913-7.
Markovitz, N. S., Filatov, F., and Roizman, B. (1999). The U(L)3 protein of herpes
simplex virus 1 is translated predominantly from the second in-frame methionine
codon and is subject to at least two posttranslational modifications. J Virol 73(10),
8010-8.
Maul, G. G., Ishov, A. M., and Everett, R. D. (1996). Nuclear domain 10 as preexisting
potential replication start sites of herpes simplex virus type-1. Virology 217(1),
67-75.
Mauri, D. N., Ebner, R., Montgomery, R. I., Kochel, K. D., Cheung, T. C., Yu, G. L.,
Ruben, S., Murphy, M., Eisenberg, R. J., Cohen, G. H., Spear, P. G., and Ware, C.
F. (1998). LIGHT, a new member of the TNF superfamily, and lymphotoxin
alpha are ligands for herpesvirus entry mediator. Immunity 8(1), 21-30.
McGeoch, D. J., Dalrymple, M. A., Davison, A. J., Dolan, A., Frame, M. C., McNab, D.,
Perry, L. J., Scott, J. E., and Taylor, P. (1988). The complete DNA sequence of
the long unique region in the genome of herpes simplex virus type 1. J. Gen.
Virol. 69, 1531-1574.
80
McGeoch, D. J., Dolan, A., Donald, S., and Rixon, F. J. (1985). Sequence determination
and genetic content of the short unique region in the genome of herpes simplex
virus type 1. J Mol Biol 181(1), 1-13.
McLauchlan, J., and Rixon, F. J. (1992). Characterization of enveloped tegument
structures (L particles) produced by alphaherpesviruses: integrity of the tegument
does not depend on the presence of capsid or envelope. J. Gen. Virol. 73(Pt 2),
269-76.
McNabb, D. S., and Courtney, R. J. (1992). Characterization of the large tegument
protein (ICP1/2) of herpes simplex virus type 1. Virology 190(1), 221-32.
Melancon, J. M., Foster, T. P., and Kousoulas, K. G. (2004). Genetic analysis of the
herpes simplex virus type 1 (HSV-1) UL20 protein domains involved in
cytoplasmic virion envelopment and virus-induced cell fusion. J Virol 78(14),
7329-43.
Mettenleiter, T. C. (2000). Aujeszky's disease (pseudorabies) virus: the virus and
molecular pathogenesis--state of the art, June 1999. Vet Res 31(1), 99-115.
Mettenleiter, T. C. (2002a). Brief overview on cellular virus receptors. Virus Res 82(1-2),
3-8.
Mettenleiter, T. C. (2002b). Herpesvirus assembly and egress. J Virol 76(4), 1537-47.
Mettenleiter, T. C., Schreurs, C., Zuckermann, F., and Ben-Porat, T. (1987a). Role of
pseudorabies virus glycoprotein gI in virus release from infected cells. J Virol
61(9), 2764-9.
Mettenleiter, T. C., Zsak, L., Kaplan, A. S., Ben-Porat, T., and Lomniczi, B. (1987b).
Role of a structural glycoprotein of pseudorabies in virus virulence. J Virol
61(12), 4030-2.
Milne, R. S., Connolly, S. A., Krummenacher, C., Eisenberg, R. J., and Cohen, G. H.
(2001). Porcine HveC, a member of the highly conserved HveC/nectin 1 family,
is a functional alphaherpesvirus receptor. Virology 281(2), 315-28.
Minson, A. C., Hodgman, T. C., Digard, P., Hancock, D. C., Bell, S. E., and Buckmaster,
E. A. (1986). An analysis of the biological properties of monoclonal antibodies
against glycoprotein D of herpes simplex virus and identification of amino acid
substitutions that confer resistance to neutralization. J Gen Virol 67 ( Pt 6), 100113.
Mo, C., and Holland, T. C. (1997). Determination of the transmembrane topology of
herpes simplex virus type 1 glycoprotein K. J. Biol. Chem. 272(52), 33305-11.
81
Mocarski, E. S., and Roizman, B. (1982). Structure and role of the herpes simplex virus
DNA termini in inversion, circularization and generation of virion DNA. Cell
31(1), 89-97.
Montgomerie, J. Z., Becroft, D. M., Croxson, M. C., Doak, P. B., and North, J. D. (1969).
Herpes-simplex-virus infection after renal transplantation. Lancet 2(7626), 86771.
Montgomery, R. I., Warner, M. S., Lum, B. J., and Spear, P. G. (1996). Herpes simplex
virus-1 entry into cells mediated by a novel member of the TNF/NGF receptor
family. Cell 87, 427-36.
Morgan, C., Rose, H. M., Holden, M., and Jones, E. P. (1959). Electron microscopic
observations on the development of herpes simplex virus. J Exp Med 110, 643-56.
Morgan, C., Rose, H. M., and Mednis, B. (1968). Electron microscopy of herpes simplex
virus. I. Entry. J Virol 2(5), 507-16.
Mori, Y., Yang, X., Akkapaiboon, P., Okuno, T., and Yamanishi, K. (2003). Human
herpesvirus 6 variant A glycoprotein H-glycoprotein L-glycoprotein Q complex
associates with human CD46. J Virol 77(8), 4992-9.
Mossman, K. L., Sherburne, R., Lavery, C., Duncan, J., and Smiley, J. R. (2000).
Evidence that herpes simplex virus VP16 is required for viral egress downstream
of the initial envelopment event. J Virol 74(14), 6287-99.
Mulder, W. A., Jacobs, L., Priem, J., Kok, G. L., Wagenaar, F., Kimman, T. G., and Pol,
J. M. (1994). Glycoprotein gE-negative pseudorabies virus has a reduced
capability to infect second- and third-order neurons of the olfactory and
trigeminal routes in the porcine central nervous system. J Gen Virol 75 ( Pt 11),
3095-106.
Muller, M. T. (1987). Binding of the herpes simplex virus immediate-early gene product
ICP4 to its own transcription start site. J Virol 61(3), 858-65.
Nahmias, A. J., Josey, W. E., Naib, Z. M., Luce, C. F., and Guest, B. A. (1970).
Antibodies to Herpesvirus hominis types 1 and 2 in humans. II. Women with
cervical cancer. Am J Epidemiol 91(6), 547-52.
Nahmias, A. J., Keyserling, H., and Kerrick, G. M. (1983). Herpes simplex. In
"Infectious diseases of the fetus and newborn infant" (J. S. Remington, and J. O.
Klein, Eds.), pp. 636-678. WB Saunders, Philadelphia.
Nahmias, A. J., Keyserling, H., and Lee, F. K. (1989). Herpes simplex viruses 1 and 2.
3rd ed. In "Viral infections of humans : epidemiology and control" (A. S. Evans,
Ed.), pp. 393-417. Plenum Medical Book Co., New York.
82
Nahmias, A. J., Lee, F. K., and Beckman-Nahmias, S. (1990). Sero-epidemiological and sociological patterns of herpes simplex virus infection in the world. Scand J Infect
Dis Suppl 69, 19-36.
Nazerian, K. (1974). DNA configuration in the core of Marek's disease virus. J Virol
13(5), 1148-50.
Neidhardt, H., Schroder, C. H., and Kaerner, H. C. (1987). Herpes simplex virus type 1
glycoprotein E is not indispensable for viral infectivity. J. Virol. 61(2), 600-3.
Newcomb, W. W., Homa, F. L., Thomsen, D. R., Booy, F. P., Trus, B. L., Steven, A. C.,
Spencer, J. V., and Brown, J. C. (1996). Assembly of the herpes simplex virus
capsid: characterization of intermediates observed during cell-free capsid
formation. J Mol Biol 263(3), 432-46.
Newcomb, W. W., Trus, B. L., Booy, F. P., Steven, A. C., Wall, J. S., and Brown, J. C.
(1993). Structure of the herpes simplex virus capsid. Molecular composition of
the pentons and the triplexes. J Mol Biol 232(2), 499-511.
Nicholson, P., Addison, C., Cross, A. M., Kennard, J., Preston, V. G., and Rixon, F. J.
(1994). Localization of the herpes simplex virus type 1 major capsid protein VP5
to the cell nucleus requires the abundant scaffolding protein VP22a. J Gen Virol
75 ( Pt 5), 1091-9.
Nicola, A. V., McEvoy, A. M., and Straus, S. E. (2003). Roles for endocytosis and low
pH in herpes simplex virus entry into HeLa and Chinese hamster ovary cells. J
Virol 77(9), 5324-32.
Nicola, A. V., and Straus, S. E. (2004). Cellular and viral requirements for rapid
endocytic entry of herpes simplex virus. J Virol 78(14), 7508-17.
Nishioka, Y., and Silverstein, S. (1977). Degradation of cellular mRNA during infection
by herpes simplex virus. Proc Natl Acad Sci U S A 74(6), 2370-4.
Nishioka, Y., and Silverstein, S. (1978a). Alterations in the protein synthetic apparatus of
Friend erythroleukemia cells infected with vesicular stomatitis virus or herpes
simplex virus. J Virol 25(1), 422-6.
Nishioka, Y., and Silverstein, S. (1978b). Requirement of protein synthesis for the
degradation of host mRNA in Friend erythroleukemia cells infected wtih herpes
simplex virus type 1. J Virol 27(3), 619-27.
Novotny, M. J., Parish, M. L., and Spear, P. G. (1996). Variability of herpes simplex
virus 1 gL and anti-gL antibodies that inhibit cell fusion but not viral infectivity.
Virology 221(1), 1-13.
83
Ojala, P. M., Sodeik, B., Ebersold, M. W., Kutay, U., and Helenius, A. (2000). Herpes
simplex virus type 1 entry into host cells: reconstitution of capsid binding and
uncoating at the nuclear pore complex in vitro. Mol Cell Biol 20(13), 4922-31.
Olson, J. K., and Grose, C. (1998). Complex formation facilitates endocytosis of the
varicella-zoster virus gE:gI Fc receptor. J Virol 72(2), 1542-51.
Olson, L. C., Buescher, E. L., Artenstein, M. S., and Parkman, P. D. (1967). Herpesvirus
infections of the human central nervous system. N Engl J Med 277(24), 1271-7.
Oroskar, A. A., and Read, G. S. (1987). A mutant of herpes simplex virus type 1 exhibits
increased stability of immediate-early (alpha) mRNAs. J Virol 61(2), 604-6.
Pellett, P. E., Kousoulas, K. G., Pereira, L., and Roizman, B. (1985). Anatomy of the
herpes simplex virus 1 strain F glycoprotein B gene: primary sequence and
predicted protein structure of the wild type and of monoclonal antibody-resistant
mutants. J. Virol. 53(1), 243-53.
Perdue, M. L., Cohen, J. C., Randall, C. C., and O'Callaghan, D. J. (1976). Biochemical
studies of the maturation of herpesvirus nucleocapsid species. Virology 74(1),
UNKNOWN.
Perdue, M. L., Kemp, M. C., Randall, C. C., and O'Callaghan, D. J. (1974). Studies of the
molecular anatomy of the L-M cell strain of equine herpes virus type 1: proteins
of the nucleocapsid and intact virion. Virology 59(1), 201-16.
Pertel, P. E. (2002). Human herpesvirus 8 glycoprotein B (gB), gH, and gL can mediate
cell fusion. J Virol 76(9), 4390-400.
Pertel, P. E., and Spear, P. G. (1996). Modified entry and syncytium formation by herpes
simpelx virus type 1 mutants selected for resistance to heparin inhibition.
Virology 226, 22-33.
Pogue-Geile, K. L., Lee, G. T., Shapira, S. K., and Spear, P. G. (1984). Fine mapping of
mutations in the fusion-inducing MP strain of herpes simplex virus type 1.
Virology 136(1), 100-9.
Pogue-Geile, K. L., and Spear, P. G. (1987). The single base pair substitution responsible
for the syn phenotype of herpes simplex virus type 1, strain MP. Virology 157,
67-74.
Polvino-Bodnar, M., Orberg, P. K., and Schaffer, P. A. (1987). Herpes simplex virus type
1 oriL is not required for virus replication or for the establishment and
reactivation of latent infection in mice. J Virol 61(11), 3528-35.
Post, L. E., Mackem, S., and Roizman, B. (1981). Regulation of alpha genes of herpes
simplex virus: expression of chimeric genes produced by fusion of thymidine
kinase with alpha gene promoters. Cell 24(2), 555-65.
84
Purifoy, D. J., Lewis, R. B., and Powell, K. L. (1977). Identification of the herpes
simplex virus DNA polymerase gene. Nature 269(5629), 621-3.
Purves, F. C., Longnecker, R. M., Leader, D. P., and Roizman, B. (1987). Herpes simplex
virus 1 protein kinase is encoded by open reading frame US3 which is not
essential for virus growth in cell culture. J Virol 61(9), 2896-901.
Quinlan, M. P., Chen, L. B., and Knipe, D. M. (1984). The intranuclear location of a
herpes simplex virus DNA-binding protein is determined by the status of viral
DNA replication. Cell 36(4), 857-68.
Rafield, L. F., and Knipe, D. M. (1984). Characterization of the major mRNAs
transcribed from the genes for glycoprotein B and DNA-binding protein ICP8 of
herpes simplex virus type 1. J. Virol. 49(3), 960-9.
Ramaswamy, R., and Holland, T. C. (1992). In vitro characterization of the HSV-1 UL53
gene product. Virology 186(2), 579-87.
Read, G. S., and Frenkel, N. (1983). Herpes simplex virus mutants defective in the
virion-associated shutoff of host polypeptide synthesis and exhibiting abnormal
synthesis of alpha (immediate early) viral polypeptides. J Virol 46(2), 498-512.
Read, G. S., Karr, B. M., and Knight, K. (1993). Isolation of a herpes simplex virus type
1 mutant with a deletion in the virion host shutoff gene and identification of
multiple forms of the vhs (UL41) polypeptide. J Virol 67(12), 7149-60.
Reeves, W. C., Corey, L., Adams, H. G., Vontver, L. A., and Holmes, K. K. (1981). Risk
of recurrence after first episodes of genital herpes. Relation to HSV type and
antibody response. N Engl J Med 305(6), 315-9.
Reynolds, A. E., Liang, L., and Baines, J. D. (2004). Conformational changes in the
nuclear lamina induced by herpes simplex virus type 1 require genes U(L)31 and
U(L)34. J Virol 78(11), 5564-75.
Reynolds, A. E., Ryckman, B. J., Baines, J. D., Zhou, Y., Liang, L., and Roller, R. J.
(2001). U(L)31 and U(L)34 proteins of herpes simplex virus type 1 form a
complex that accumulates at the nuclear rim and is required for envelopment of
nucleocapsids. J Virol 75(18), 8803-17.
Reynolds, A. E., Wills, E. G., Roller, R. J., Ryckman, B. J., and Baines, J. D. (2002).
Ultrastructural localization of the herpes simplex virus type 1 UL31, UL34, and
US3 proteins suggests specific roles in primary envelopment and egress of
nucleocapsids. J Virol 76(17), 8939-52.
Rice, S. A., Long, M. C., Lam, V., and Spencer, C. A. (1994). RNA polymerase II is
aberrantly phosphorylated and localized to viral replication compartments
following herpes simplex virus infection. J Virol 68(2), 988-1001.
85
Rixon, F. J., Addison, C., McGregor, A., Macnab, S. J., Nicholson, P., Preston, V. G.,
and Tatman, J. D. (1996). Multiple interactions control the intracellular
localization of the herpes simplex virus type 1 capsid proteins. J Gen Virol 77 ( Pt
9), 2251-60.
Rixon, F. J., Addison, C., and McLauchlan, J. (1992). Assembly of enveloped tegument
structures (L particles) can occur independently of virion maturation in herpes
simplex virus type 1-infected cells. J. Gen. Virol. 73(Pt 2), 277-84.
Roberts, S. R., Ponce de Leon, M., Cohen, G. H., and Eisenberg, R. J. (1991). Analysis of
the intracellular maturation of the herpes simplex virus type 1 glycoprotein gH in
infected and transfected cells. Virology 184(2), 609-24.
Roizman, B. (1962). Polykaryocytosis. Cold Spring Harb Symp Quant Biol 27, 327-42.
Roizman, B., Bartha, A., and Biggs, P. M. (1973). Editorial: Provisional labels for
herpesviruses. J Gen Virol 20(3), 417-9.
Roizman, B., Desrosiers, R. C., Fleckenstein, B., Lopez, C., Minson, A. C., and Studdert,
M. J. (1992). The family Herpesviridae: an update. The Herpesvirus Study Group
of the International Committee on Taxonomy of Viruses. Archives Of Virology.
123(3-4), 425-49.
Roizman, B., and Furlong, D. (1974). The replication of herpesviruses. In
"Comprehensive virology" (H. Fraenkel-Conrat, and R. R. Wagner, Eds.), Vol. 3,
pp. 229-403. Plenum Press, New York,.
Roizman, B., and Knipe, D. M. (2001). Herpes Simplex Viruses and Their Replication.
Third edition ed. In "Fields Virology" (D. M. Knipe, and P. M. Howley, Eds.),
Vol. 2, pp. 2399-2459. 2 vols. Lippincott-Williams & Wilkins, Philadelphia, PA.
Roizman, B., and Pellett, P. (2001). The Family Herpesviridae: A Brief Introduction.
Third edition ed. In "Fields Virology" (D. M. Knipe, and P. M. Howley, Eds.),
Vol. 2, pp. 2381-2397. 2 vols. Lippincott-Williams & Wilkins, Philadelphia, PA.
Roller, R. J., Zhou, Y., Schnetzer, R., Ferguson, J., and DeSalvo, D. (2000). Herpes
simplex virus type 1 U(L)34 gene product is required for viral envelopment. J
Virol 74(1), 117-29.
Roop, C., Hutchinson, L., and Johnson, D. C. (1993). A mutant herpes simplex virus type
1 unable to express glycoprotein L cannot enter cells, and its particles lack
glycoprotein H. J. Virol. 67(4), 2285-97.
Rowe, W. P., Hartley, J. W., Waterman, S., Turner, H. C., and Huebner, R. J. (1956).
Cytopathogenic agent resembling human salivary gland virus recovered from
tissue cultures of human adenoids. Proc Soc Exp Biol Med 92(2), 418-24.
86
Ruyechan, W. T., Morse, L. S., Knipe, D. M., and Roizman, B. (1979). Molecular
genetics of herpes simplex virus. II. Mapping of the major viral glycoproteins
and of the genetic loci
specifying the social behavior of infected cells. J.
Virol. 29, 677-697.
Sahu, A., Isaacs, S. N., Soulika, A. M., and Lambris, J. D. (1998). Interaction of vaccinia
virus complement control protein with human complement proteins: factor Imediated degradation of C3b to iC3b1 inactivates the alternative complement
pathway. J Immunol 160(11), 5596-604.
Salahuddin, S. Z., Ablashi, D. V., Markham, P. D., Josephs, S. F., Sturzenegger, S.,
Kaplan, M., Halligan, G., Biberfeld, P., Wong-Staal, F., Kramarsky, B., and et al.
(1986). Isolation of a new virus, HBLV, in patients with lymphoproliferative
disorders. Science 234(4776), 596-601.
Sanders, P. G., Wilkie, N. M., and Davison, A. J. (1982). Thymidine kinase deletion
mutants of herpes simplex virus type 1. J. Gen. Virol. 63(2), 277-95.
Sarmiento, M., Haffey, M., and Spear, P. G. (1979). Membrane proteins specified by
herpes simplex viruses. III. Role of glycoprotein VP7(B2) in virion infectivity. J
Virol 29(3), 1149-58.
Scott, E. S., and O'Hare, P. (2001). Fate of the inner nuclear membrane protein lamin B
receptor and nuclear lamins in herpes simplex virus type 1 infection. J Virol
75(18), 8818-30.
Segal, A. L., Katcher, A. H., Brightman, V. J., and Miller, M. F. (1974). Recurrent herpes
labialis, recurrent aphthous ulcers, and the menstrual cycle. J Dent Res 53(4),
797-803.
Shieh, M. T., WuDunn, D., Montgomery, R. I., Esko, J. D., and Spear, P. G. (1992). Cell
surface receptors for herpes simplex virus are heparan sulfate proteoglycans. J.
Cell. Biol. 116(5), 1273-81.
Ship, II, Miller, M. F., and Ram, C. (1977). A retrospective study of recurrent herpes
labialis (RHL) in a professional population, 1958-1971. Oral Surg Oral Med Oral
Pathol 44(5), 723-30.
Shukla, D., Dal Canto, M. C., Rowe, C. L., and Spear, P. G. (2000). Striking similarity of
murine nectin-1alpha to human nectin-1alpha (HveC) in sequence and activity as
a glycoprotein D receptor for alphaherpesvirus entry. J Virol 74(24), 11773-81.
Shukla, D., Liu, J., Blaiklock, P., Shworak, N. W., Bai, X., Esko, J. D., Cohen, G. H.,
Eisenberg, R. J., Rosenberg, R. D., and Spear, P. G. (1999). A novel role for 3-Osulfated heparan sulfate in herpes simplex virus 1 entry. Cell 99(1), 13-22.
Sinclair, M. C., McLauchlan, J., Marsden, H., and Brown, S. M. (1994). Characterization
of a herpes simplex virus type 1 deletion variant (1703) which under-produces
87
Vmw63 during immediate early conditions of infection. J Gen Virol 75 ( Pt 5),
1083-9.
Smibert, C. A., Popova, B., Xiao, P., Capone, J. P., and Smiley, J. R. (1994). Herpes
simplex virus VP16 forms a complex with the virion host shutoff protein vhs. J
Virol 68(4), 2339-46.
Smiley, J. R., Duncan, J., and Howes, M. (1990). Sequence requirements for DNA
rearrangements induced by the terminal repeat of herpes simplex virus type 1
KOS DNA. J Virol 64(10), 5036-50.
Smith, M. G. (1956). Propagation in tissue cultures of a cytopathogenic virus from
human salivary gland virus (SGV) disease. Proc Soc Exp Biol Med 92(2), 424-30.
Sodeik, B., Ebersold, M. W., and Helenius, A. (1997). Microtubule-mediated transport of
incoming herpes simplex virus 1 capsids to the nucleus. J Cell Biol 136(5), 100721.
Spear, P. G. (1985a). Antigenic structure of herpes simplex viruses. In
"Immunochemistry of viruses: The basis for serodiagnosis and vaccines" (M. H.
V. Van Regenmortel, and A. R. Neurath, Eds.), pp. 425-446. Elsevier Science,
Amsterdam.
Spear, P. G. (1985b). Glycoproteins specified by herpes simplex viruses. In "The
herpesviruses" (B. Roizman, Ed.), Vol. 3, pp. 315-356. Plenum, New York.
Spear, P. G. (1993). Membrane fusion induced by herpes simplex virus. In "Viral fusion
mechanisms" (J. Bentz, Ed.), pp. 201-232. CRC Press., Boca Raton, Fla.
Spear, P. G. (2004). Herpes simplex virus: receptors and ligands for cell entry. Cell
Microbiol 6(5), 401-10.
Spear, P. G., Eisenberg, R. J., and Cohen, G. H. (2000). Three classes of cell surface
receptors for alphaherpesvirus entry. Virology 275(1), 1-8.
Spear, P. G., and Longnecker, R. (2003). Herpesvirus entry: an update. J Virol 77(19),
10179-85.
Spencer, J. V., Newcomb, W. W., Thomsen, D. R., Homa, F. L., and Brown, J. C. (1998).
Assembly of the herpes simplex virus capsid: preformed triplexes bind to the
nascent capsid. J Virol 72(5), 3944-51.
Spengler, M., Niesen, N., Grose, C., Ruyechan, W. T., and Hay, J. (2001). Interactions
among structural proteins of varicella zoster virus. Arch Virol Suppl(17), 71-9.
Spruance, S. L., and Crumpacker, C. S. (1982). Topical 5 percent acyclovir in
polyethylene glycol for herpes simplex labialis. Antiviral effect without clinical
benefit. Am J Med 73(1A), 315-9.
88
Spruance, S. L., Crumpacker, C. S., Schnipper, L. E., Kern, E. R., Marlowe, S., Arndt, K.
A., and Overall, J. C., Jr. (1984). Early, patient-initiated treatment of herpes
labialis with topical 10% acyclovir. Antimicrob Agents Chemother 25(5), 553-5.
Spruance, S. L., Overall, J. C., Jr., Kern, E. R., Krueger, G. G., Pliam, V., and Miller, W.
(1977). The natural history of recurrent herpes simplex labialis: implications for
antiviral therapy. N Engl J Med 297(2), 69-75.
Steven, A. C., and Spear, P. G. (1997). Herpesvirus capsid assembly and envelopment. In
"Structural Biology of Viruses" (W. Chiu, R. M. Burnett, and R. L. Garcea, Eds.),
pp. 312-351. Oxford University Press, New York.
Stow, N. D. (1982). Localization of an origin of DNA replication within the TRS/IRS
repeated region of the herpes simplex virus type 1 genome. Embo J 1(7), 863-7.
Stow, N. D., and Stow, E. C. (1986). Isolation and characterization of a herpes simplex
virus type 1 mutant containing a deletion within the gene encoding the immediate
early polypeptide Vmw110. J Gen Virol 67 ( Pt 12), 2571-85.
Sullivan, V., and Smith, G. L. (1987). Expression and characterization of herpes simplex
virus type 1 (HSV-1) glycoprotein G (gG) by recombinant vaccinia virus:
neutralization of HSV-1 infectivity with anti-gG antibody. J Gen Virol 68 ( Pt
10), 2587-98.
Takahashi, K., Nakanishi, H., Miyahara, M., Mandai, K., Satoh, K., Satoh, A., Nishioka,
H., Aoki, J., Nomoto, A., Mizoguchi, A., and Takai, Y. (1999). Nectin/PRR: an
immunoglobulin-like cell adhesion molecule recruited to cadherin-based adherens
junctions through interaction with Afadin, a PDZ domain-containing protein. J
Cell Biol 145(3), 539-49.
Takai, Y., and Nakanishi, H. (2003). Nectin and afadin: novel organizers of intercellular
junctions. J Cell Sci 116(Pt 1), 17-27.
Tal-Singer, R., Peng, C., Ponce, D. L. M., Abrams, W. R., Banfield, B. W., Tufaro, F.,
Cohen, G. H., and Eisenberg, R. J. (1995). Interaction of herpes simplex virus
glycoprotein gC with mammalian cell surface molecules. J. Virol. 69(7), 4471-83.
Thomsen, D. R., Newcomb, W. W., Brown, J. C., and Homa, F. L. (1995). Assembly of
the herpes simplex virus capsid: requirement for the carboxyl-terminal twentyfive amino acids of the proteins encoded by the UL26 and UL26.5 genes. J Virol
69(6), 3690-703.
Tirabassi, R. S., and Enquist, L. W. (1998). Role of envelope protein gE endocytosis in
the pseudorabies virus life cycle. J Virol 72(6), 4571-9.
Tirabassi, R. S., Townley, R. A., Eldridge, M. G., and Enquist, L. W. (1997).
Characterization of pseudorabies virus mutants expressing carboxy-terminal
89
truncations of gE: evidence for envelope incorporation, virulence, and
neurotropism domains. J Virol 71(9), 6455-64.
Tognon, M., Manservigi, R., Cavrini, V., and Campadelli-Fiume, G. (1984).
Characterization of a herpes simplex virus type 1 mutant resistant to
benzhydrazone, a selective inhibitor of herpesvirus glycosylation. Proc Natl Acad
Sci U S A 81(8), 2440-3.
Tran, L. C., Kissner, J. M., Westerman, L. E., and Sears, A. E. (2000). A herpes simplex
virus 1 recombinant lacking the glycoprotein G coding sequences is defective in
entry through apical surfaces of polarized epithelial cells in culture and in vivo.
Proc Natl Acad Sci U S A 97(4), 1818-22.
Trus, B. L., Booy, F. P., Newcomb, W. W., Brown, J. C., Homa, F. L., Thomsen, D. R.,
and Steven, A. C. (1996). The herpes simplex virus procapsid: structure,
conformational changes upon maturation, and roles of the triplex proteins VP19c
and VP23 in assembly. J Mol Biol 263(3), 447-62.
Turner, A., Bruun, B., Minson, T., and Browne, H. (1998). Glycoproteins gB, gD, and
gHgL of herpes simplex virus type 1 are necessary and sufficient to mediate
membrane fusion in a Cos cell transfection system. J Virol 72(1), 873-5.
Uprichard, S. L., and Knipe, D. M. (1996). Herpes simplex ICP27 mutant viruses exhibit
reduced expression of specific DNA replication genes. J Virol 70(3), 1969-80.
van Genderen, I. L., Brandimarti, R., Torrisi, M. R., Campadelli, G., and van Meer, G.
(1994). The phospholipid composition of extracellular herpes simplex virions
differs from that of host cell nuclei. Virology 200(2), 831-6.
Van Regenmortel, M. H. V., Fauquet, C. M., Bishop, D. H. L., Carstens, E. B., Estes, M.
K., Lemon, S. M., Maniloff, J., Mayo, M. A., McGeoch, D. J., and Pringle, C. R.
(2000). "Virus taxonomy : classification and nomenclature of viruses : seventh
report of the International Committee on Taxonomy of Viruses." Academic Press,
San Diego.
Varmuza, S. L., and Smiley, J. R. (1985). Signals for site-specific cleavage of HSV
DNA: maturation involves two separate cleavage events at sites distal to the
recognition sequences. Cell 41(3), 793-802.
Vidal, J. B. (1873). Ann Dermatol Syphiligr 1, 350.
Vlazny, D. A., Kwong, A., and Frenkel, N. (1982). Site-specific cleavage/packaging of
herpes simplex virus DNA and the selective maturation of nucleocapsids
containing full-length viral DNA. Proc Natl Acad Sci U S A 79(5), 1423-7.
Wagner, E. K. (1985). Individual HSV transcripts: Characterisation of specific genes. In
"The Herpesviruses" (B. Roizman, Ed.), pp. 45-104. Plenum Press, New York.
90
Ward, P. L., Avitabile, E., Campadelli-Fiume, G., and Roizman, B. (1998). Conservation
of the architecture of the Golgi apparatus related to a differential organization of
microtubules in polykaryocytes induced by syn- mutants of herpes simplex virus
1. Virology 241(2), 189-99.
Ward, P. L., Campadelli-Fiume, G., Avitabile, E., and Roizman, B. (1994). Localization
and putative function of the UL20 membrane protein in cells infected with herpes
simplex virus 1. J. Virol. 68(11), 7406-17.
Warner, M. S., Geraghty, R. J., Martinez, W. M., Montgomery, R. I., Whitbeck, J. C.,
Xu, R., Eisenberg, R. J., Cohen, G. H., and Spear, P. G. (1998). A cell surface
protein with herpesvirus entry activity (HveB) confers susceptibility to infection
by mutants of herpes simplex virus type 1, herpes simplex virus type 2, and
pseudorabies virus. Virology 246(1), 179-89.
Watson, R. J., Weis, J. H., Salstrom, J. S., and Enquist, L. W. (1982). Herpes simplex
virus type-1 glycoprotein D gene: nucleotide sequence and expression in
Escherichia coli. Science 218(4570), 381-4.
Weller, S. K., Spadaro, A., Schaffer, J. E., Murray, A. W., Maxam, A. M., and Schaffer,
P. A. (1985). Cloning, sequencing, and functional analysis of oriL, a herpes
simplex virus type 1 origin of DNA synthesis. Mol Cell Biol 5(5), 930-42.
Weller, T. H., and Stoddard, M. B. (1952). Intranuclear inclusion bodies in cultures of
human tissue inoculated with varicella vesicle fluid. J Immunol 68(3), 311-9.
Whealy, M. E., Card, J. P., Robbins, A. K., Dubin, J. R., Rziha, H. J., and Enquist, L. W.
(1993). Specific pseudorabies virus infection of the rat visual system requires both
gI and gp63 glycoproteins. J Virol 67(7), 3786-97.
Whitley, R., Arvin, A., Prober, C., Corey, L., Burchett, S., Plotkin, S., Starr, S., Jacobs,
R., Powell, D., Nahmias, A., and et al. (1991). Predictors of morbidity and
mortality in neonates with herpes simplex virus infections. The National Institute
of Allergy and Infectious Diseases Collaborative Antiviral Study Group. N Engl J
Med 324(7), 450-4.
Whitley, R. J. (2001). Herpes Simplex Viruses. Third edition ed. In "Fields Virology" (D.
M. Knipe, and P. M. Howley, Eds.), Vol. 2, pp. 2461-2509. 2 vols. LippincottWilliams & Wilkins, Philadelphia, PA.
Whitley, R. J., Levin, M., Barton, N., Hershey, B. J., Davis, G., Keeney, R. E., Whelchel,
J., Diethelm, A. G., Kartus, P., and Soong, S. J. (1984). Infections caused by
herpes simplex virus in the immunocompromised host: natural history and topical
acyclovir therapy. J Infect Dis 150(3), 323-9.
Whitley, R. J., Soong, S. J., Dolin, R., Galasso, G. J., Ch'ien, L. T., and Alford, C. A.
(1977). Adenine arabinoside therapy of biopsy-proved herpes simplex
91
encephalitis. National Institute of Allergy and Infectious Diseases collaborative
antiviral study. N Engl J Med 297(6), 289-94.
Whitley, R. J., Soong, S. J., Hirsch, M. S., Karchmer, A. W., Dolin, R., Galasso, G.,
Dunnick, J. K., and Alford, C. A. (1981). Herpes simplex encephalitis: vidarabine
therapy and diagnostic problems. N Engl J Med 304(6), 313-8.
Wildy, P. (1973). Herpes: history and classification. In "The herpesviruses" (A. S.
Kaplan, Ed.), pp. 1-25. Academic Press, New York.
Wildy, P., and Watson, D. H. (1962). Electron microscopic studies on the architecture of
animal viruses. Cold Spring Harb Symp Quant Biol 27, 25-47.
Wisner, T., Brunetti, C., Dingwell, K., and Johnson, D. C. (2000). The extracellular
domain of herpes simplex virus gE is sufficient for accumulation at cell junctions
but not for cell-to-cell spread. J Virol 74(5), 2278-87.
Wu, C. A., Nelson, N. J., McGeoch, D. J., and Challberg, M. D. (1988). Identification of
herpes simplex virus type 1 genes required for origin-dependent DNA synthesis. J
Virol 62(2), 435-43.
Wu, S. X., Zhu, X. P., and Letchworth, G. J. (1998). Bovine herpesvirus 1 glycoprotein
M forms a disulfide-linked heterodimer with the U(L)49.5 protein. J Virol 72(4),
3029-36.
WuDunn, D., and Spear, P. G. (1989). Initial interaction of herpes simplex virus with
cells is binding to heparan sulfate. J. Virol. 63(1), 52-8.
Yoon, M., Zago, A., Shukla, D., and Spear, P. G. (2003). Mutations in the N termini of
herpes simplex virus type 1 and 2 gDs alter functional interactions with the
entry/fusion receptors HVEM, nectin-2, and 3-O-sulfated heparan sulfate but not
with nectin-1. J Virol 77(17), 9221-31.
Zago, A., and Spear, P. G. (2003). Differences in the N termini of herpes simplex virus
type 1 and 2 gDs that influence functional interactions with the human entry
receptor Nectin-2 and an entry receptor expressed in Chinese hamster ovary cells.
J Virol 77(17), 9695-9.
Zelus, B. D., Stewart, R. S., and Ross, J. (1996). The virion host shutoff protein of herpes
simplex virus type 1: messenger ribonucleolytic activity in vitro. J Virol 70(4),
2411-9.
Zhou, Z. H., Chen, D. H., Jakana, J., Rixon, F. J., and Chiu, W. (1999). Visualization of
tegument-capsid interactions and DNA in intact herpes simplex virus type 1
virions. J Virol 73(4), 3210-8.
92
Zhou, Z. H., He, J., Jakana, J., Tatman, J. D., Rixon, F. J., and Chiu, W. (1995).
Assembly of VP26 in herpes simplex virus-1 inferred from structures of wild-type
and recombinant capsids. Nat Struct Biol 2(11), 1026-30.
Zhu, Q., and Courtney, R. J. (1994). Chemical cross-linking of virion envelope and
tegument proteins of herpes simplex virus type 1. Virology 204(2), 590-9.
Zsak, L., Zuckermann, F., Sugg, N., and Ben-Porat, T. (1992). Glycoprotein gI of
pseudorabies virus promotes cell fusion and virus spread via direct cell-to-cell
transmission. J Virol 66(4), 2316-25.
Zuckermann, F. A., Mettenleiter, T. C., Schreurs, C., Sugg, N., and Ben-Porat, T. (1988).
Complex between glycoproteins gI and gp63 of pseudorabies virus: its effect on
virus replication. J Virol 62(12), 4622-6.
93
CHAPTER II
AN α-HELICAL DOMAIN WITHIN THE CARBOXYL TERMINUS
OF HERPES SIMPLEX VIRUS TYPE 1 (HSV-1) GLYCOPROTEIN B
(gB) IS ASSOCIATED WITH CELL FUSION AND RESISTANCE TO
HEPARIN INHIBITION OF CELL FUSION*
INTRODUCTION
Viral glycoproteins are key determinants in membrane fusion events throughout
the virus life cycle. Herpes simplex viruses specify at least twelve glycoproteins: gB, gC,
gD, gE, gG, gH, gI, gJ, gK, gL gM, and gN, which are expressed in infected cells. These
glycoproteins function in several important roles including pH independent virus entry
via fusion of the viral envelope with cellular membranes, cell-to-cell spread, virusinduced cell fusion, and egress of infectious virion particles (reviewed in Mettenleiter,
2000; Spear, 1993). Recombinant mutant viruses that lack either gB (UL27), gD (US6),
gH (UL22) or gL (UL1) attach to cell surfaces through cell surface glycosaminoglycans,
but fail to penetrate, indicating these glycoproteins are required at least in part for virusto-cell fusion (Cai, Gu, and Person, 1988; Forrester et al., 1992; Ligas and Johnson,
1988; Roop, Hutchinson, and Johnson, 1993). Either of two HSV-1 glycoproteins, gC
(UL44) or gB, can mediate binding of virus to heparan sulfate proteoglycans on cellular
surfaces. Accordingly, attachment is inhibited by soluble heparin, a glycosaminoglycan
closely related to heparan sulfate (Gruenheid et al., 1993; Herold et al., 1994; Herold et
al., 1991; Shieh et al., 1992; WuDunn and Spear, 1989). Fusion between the viral
Reprinted from Virology, Vol 287, T. P. Foster, J. M. Melancon, and K. G. Kousoulas, An α-Helical
Domain within the Carboxyl Terminus of Herpes Simplex Virus Type 1 (HSV-1) glycoprotein B (gB) is
Associated with Cell Fusion and Resistance to Heparin Inhibition of Cell Fusion, Pages 18-29, Copyright
2001, with permission from Elsevier.
*
94
envelope and cell membrane requires gB, gD and gH/gL, and possibly the tegument
protein UL25 (Addison et al., 1984; Cai, Gu, and Person, 1988; Forrester et al., 1992;
Ligas and Johnson, 1988; Roop, Hutchinson, and Johnson, 1993). Penetration of virus
requires the interaction of gD with specific cell surface receptors (Cocchi et al., 1998;
Geraghty et al., 1998; Montgomery et al., 1996; Warner et al., 1998).
Spread of infectious virus occurs either by release of virions to extracellular
spaces or through virus-induced cell-to-cell fusion. Mutations that cause extensive virus
induced cell-to-cell fusion can arise in at least four genes of the HSV genome: the UL53gK gene, the gB gene, the UL20 gene, and the UL24 gene (Baines et al., 1991; Bond and
Person, 1984; Bzik et al., 1984b; Cai, Gu, and Person, 1988; Debroy, Pederson, and
Person, 1985; DeLuca et al., 1982; Dolter, Ramaswamy, and Holland, 1994; Engel,
Boyer, and Goodman, 1993; Gage, Levine, and Glorioso, 1993; Hutchinson et al., 1992;
Jacobson, Martin, and Coen, 1989; Pogue-Geile and Spear, 1987; Ruyechan et al., 1979;
Sanders, Wilkie, and Davison, 1982; Weise et al., 1987). However, like virus-to-cell
fusion, gB, gD, gH, and gL have been shown to be necessary and sufficient to cause
virus-induced cell fusion. The evidence is twofold. First, mutant viruses that do not
express any one of these four glycoproteins fail to produce syncytia (Cai, Gu, and Person,
1988; Davis-Poynter et al., 1994; Ligas and Johnson, 1988; Roop, Hutchinson, and
Johnson, 1993). Similarly, transient expression of HSV-1 or HSV-2 gB, gD, gH and gL
causes polykaryocyte formation in transfected Cos cells (Muggeridge, 2000; Turner et
al., 1998). Glycoproteins gE (US8), gI(US7), and gM (UL10), as well as the UL45
protein, may be able to enhance virus-induced cell fusion, since deletions of these genes
cause inhibition of cell fusion (Davis-Poynter et al., 1994; Haanes et al., 1994; Visalli
95
and Brandt, 1991). Similarly, gC may be able to inhibit virus-induced cell fusion of at
least some cell types, which may explain the lack of gC expression in many syncytial
mutants (Manservigi, Spear, and Buchan, 1977; Novotny, Parish, and Spear, 1996; Pertel
and Spear, 1996). It is apparent there are similarities as well as significant differences
between the virus-induced cell fusion and fusion of cells by transient co-expression of
gB, gD, gH/gL. Both systems require the expression of gD receptors for fusion to occur
(Pertel et al., 2001; Terry-Allison et al., 1998); however, the transient expression system,
in contrast to the virus induced cell fusion, does not require the presence of heparan
sulfate (Pertel et al., 2001; Shieh and Spear, 1994). Additionally, viral proteins that
exhibit the ability to influence virus-induced cell fusion do not produce similar effects in
the transient fusion assay system (Klupp, Nixdorf and Mettenleiter, 2000; Turner et al.,
1998).
Glycoprotein B is highly conserved across all subfamilies of herpesviruses. It
exists within cells as a homodimeric type I integral membrane protein that is Nglycosylated at multiple sites within the polypeptide. Each 904 amino acid (aa) monomer
includes a cleavable signal sequence, an extracellular domain of 696 aa, a 69 aa triple
pass transmembrane region, and a 109 aa carboxyl terminal domain. Single amino acid
substitutions within two regions of the intracellular cytoplasmic domain of gB were
shown to cause syncytium formation and were designated Region I (aa positions 816 and
817) and Region II (aa positions 853, 854, and 857) (Bzik et al., 1984a; Cai et al., 1988;
Gage, Levine, and Glorioso, 1993; Walev et al., 1994). Deletion of the carboxyl terminal
41 amino acids of gB did not adversely affect viral infectivity and replication; whereas,
96
deletion of the carboxyl terminal 28 amino acids caused extensive virus-induced cell
fusion (Baghian et al., 1993; Huff et al., 1988).
We extended our previous findings by examining the role of specific carboxyl
terminal domains of gB in cell fusion in the context of viral infection as well as in a
transfection-fusion system. We show that an α-helical domain within the gB carboxyl
terminus appears to regulate cell fusion. Moreover, we provide evidence that heparin
inhibition of gB-mediated cell fusion is also regulated by the carboxyl terminus of gB for
both virus-induced cell fusion as well as fusion by co-expression of gB, gD, and gH/gL.
In this regard, gB-mediated fusion functions similarly for both fusion systems.
MATERIALS AND METHODS
Cells and Viruses
Vero and COS7 cells were obtained from ATCC (Rockville, MD) and grown in
Dulbecco’s modified Eagle’s Medium (DMEM) supplemented with 10% fetal calf serum
(FCS). KOS/EGFP specifies the enhanced green fluorescence protein (EGFP) from a
CMV-EGFP gene cassette inserted within the intergenic region between the UL53 and
UL54 open reading frames (ORFs) (Foster, Chouljenko, and Kousoulas, 1999). In order
to facilitate visualization of syncytia formation, recombinant viruses amb1511/EGFP,
MP/EGFP, and tsB5/EGFP were constructed by insertion of a CMV-EGFP gene cassette
as described in detail previously for the KOS/EGFP virus (Foster, Chouljenko, and
Kousoulas, 1999). All viruses were propagated on Vero cells.
Construction of Expression Plasmids
Plasmid p9-2080 specifies a wild type (wt) copy of gB; whereas, plasmids p91511, p9-1513, p9-1528, p9-1537 specify carboxyl-terminally truncated gB driven by the
97
Adenovirus Major Late Promoter (Ad MLP) and were as described previously (Baghian
et al ., 1993). The entire open reading frames (ORFs) for gB and gD were amplified by
PCR with specific primers that removed the 3´ stop codon such that the genes were
inserted in frame with a V5 epitope tag when ligated to pcDNA3.1 TA TOPO to generate
plasmids pCMV-gB and pCMV-gD. Truncations in gB were generated by PCR.
Plasmids gB(∆36), gB(∆28), gB(∆16), express C-terminally truncated derivates of gB of
36, 28, and 16 amino acids (aa) respectively. A gB gene specifying a single codon
change from alanine to proline at amino acid 874 was produced by splice-overlap
extension (SOE) PCR utilizing synthetic oligonucleotides containing the point mutation.
The PCR derived gB(A874P) was ligated into pcDNA3.1 TA TOPO to generate plasmid
pCMV-gB(A874P). All plasmid inserts were sequenced by Sanger dideoxy-chain
termination methodology using an automated DNA sequencer (PE Biosystems, 377
Sequencer). Plasmid pcDNA3 gHgL were kindly provided by A. Minson.
Cell Fusion Assay
In vitro cell fusion assays were performed essentially as described previously
(Klupp, Nixdorf, and Mettenleiter, 2000; Muggeridge, 2000; Turner et al., 1998). Briefly,
the day prior to transfection, 35 mm 6 well plates were seeded with 8 x 104 COS7 cells.
A total of 6 µg of plasmid DNA encoding the indicated glycoproteins was transfected
with Superfect (Qiagen, Valencia CA) according to the manufacturer’s directions. Three
hours post-transfection, media was aspirated and cells were overlaid with DMEM 5%
FCS. Where indicated, heparin (10mg/ml) was included in the overlay media. Twentyfour hours post transfection, cells were overlaid with 4 x105 freshly trypsinized COS7
98
cells per well with or without the inclusion of heparin. Cells were subsequently
incubated at 37°C for an additional 12 hours before levels of fusion were determined.
Immunofluorescence Assay and Quantitation of Fusion
Cells were fixed with 100% methanol for 10 minutes, washed twice with TBS,
and incubated at room temperature for 1 hour with the primary antibody anti-V5
(Invitrogen, Carlsbad, CA) diluted 1:200 in TBS-10% goat serum. Cells were then
washed three times with TBS and incubated for one hour at room temperature with FITC
conjugated goat anti-mouse IgG (Sigma, St. Louis, MO) diluted 1:100 in TBS-10%goat
serum. After incubation, excess antibody was removed by washing three times with
TBS. Propidium iodide nuclear counterstain (Molecular Probes, Eugene, OR) was
applied in order to facilitate quantitation of fusion. Fusion was examined on a Nikon
Diaphot inverted fluorescent microscope. The number of nuclei in 100 transfected cells
or polykaryocytes per well was determined by two independent observers. Transfected
cells were defined as those cells that were IFA positive. Average values and standard
deviations were then calculated from the combined data obtained by the two observers.
Percent change in levels of fusion over wild-type (wt) gB was calculated by the formula
[(average nuclei mutated gB)-(average nuclei wt gB)]/ (average nuclei wt gB) x 100%.
Percent inhibition of fusion by heparin was defined as {[(average nuclei of untreated
assay)-1]-[(average nuclei of heparin treated assay)-1]}/ [(average nuclei of untreated
assay)-1] x 100%.
Heparin Inhibition of Viral Polykaryocyte Formation
Subconfluent Vero cells were infected at an MOI of 0.1 for 1 hour and
subsequently overlaid with DMEM-2% FCS, 1% methylcellulose either lacking or
99
containing heparin (10mg/ml). The amount of heparin used in the virus-free fusion
inhibition experiments was chosen on the basis of being the amount required to inhibit
virus-induced cell fusion. Lower amounts of heparin (1 mg/ml instead of 10 mg/ml),
effectively inhibited virus-free fusion caused by wild type gB. Viral plaques were
visualized by phase-contrast and fluorescent microscopy at 24 hours post infection.
RESULTS
The Effect of gB Truncations on Virus-Independent Cell Fusion
Previously, we investigated the effect of several gB truncations on virus-induced
cell fusion. We found that a recombinant virus expressing a 28 amino acid truncated gB
resulted in extensive virus-induced cell fusion (Baghian et al., 1993). A schematic of the
predicted structure of the carboxyl terminus of gB showing the location of relevant
truncations and mutations is shown in figure 2.1. The 28 amino acid deletion of gB
disrupts the predicted α-helical structure H17b composed of 16 amino acids. To assess
whether these gB truncations caused similar effects in a virus-independent cell fusion
assay, we transfected COS7 cells with plasmids constitutively expressing glycoproteins
gD, gH and gL in the presence of different mutated gB (Fig. 2.2). The extent of cell
fusion was quantified by counting nuclei in 100 polykaryocytes per assay by two
independent observers (Fig. 2.3, Table 2.1). As shown previously, transient expression of
full-length, wild-type gB, gD, gH, and gL genes under the human cytomegalovirus
immediate early promoter (CMV-IE) caused limited polykaryocyte formation (Turner et
al., 1998) (Fig. 2.2A). Similar numbers of polykaryocytes were formed when the CMVIE driven gB gene was substituted with plasmid p9-2080 expressing gB under the
adenovirus major late promoter (Ad-MLP). (Fig. 2.2B). Transfection controls that did not
100
L
P
M
A
Y
P
K
L
T
T
K
E
L
K
N
P
T
P
D A
S G
1528 (∆49)
N
E
G
H17a
E G
1513 (∆64)
gB(syn3)
E
R to H
K
A
I
A
A
T
A
D D A L E R M R M L S M R E K K
E
A
Y
E
V
K
E
H
K
E
L
T
K
V
M
L S
R R R V
T K S
A
M
A
D
N
T
Y
N
Q
T
V P
F
G
H17b
gB(∆16)
G
D
D K
N
gB(∆36)
1511/gB(∆28)
gB(A874P)
A D E
D
L D
Figure 2.1: Schematic diagram of the carboxyl terminus of HSV-1 gB. The predicted
secondary structure of the gB carboxyl terminal 109 amino acids is shown as described
previously (Baghian et al., 1993; Pellett et al., 1985). The location of the gB truncations
and mutations specified by plasmids p9-1513, p9-1528, p9-1511, pCMV-gB(∆28),
pCMV-gB(∆36), pCMV-gB(A874P), pCMV-gB(∆16), pCMV-gB(syn3) are shown. The
symbol ∆ followed by a number indicates the number of amino acids deleted from the
carboxyl terminus of gB. The location of the two predicted α−helices H17a and H17b are
shown and marked by dashed lines.
101
Figure 2.2: Truncation of the carboxyl terminal 28 amino acid of gB causes
extensive fusion of transfected Cos cells. Cells were transfected with plasmids
expressing gD, gH/gL, and plasmid A: pCMV-gB, B: p9-2080, C: p9-1537, D: p9-1513,
E: p9-1528, F: p9-1511. Transfected cells were visualized by IFA at 36 hours after
transfection (65X magnification).
102
50
Nuclei per Transfected Cell
40
30
20
10
0
gB, gD
gHgL
2080, gD
gHgL
1537, gD
gHgL
1513, gD
gHgL
1528, gD
gHgL
1511, gD
gHgL
Plasmids Transfected
Figure 2.3: Quantitation of the effect of gB truncation on fusion of transfected Cos
Cells. Nuclei in 100 IFA positive cells per assay were counted by two observers and the
average values and standard deviation were calculated.
Table 2.1: Truncation of the carboxyl terminal 28 amino acid of gB causes extensivc
fusion of transfected Cos cells.
Plasmids Transfected
(all include gD, gHgL)
Total nuclei counted
Average nuclei
Standard Deviation
Standard Error
Median
Smallest # of nuclei
Largest # of nuclei
% change from gB
gB
2080
1537
1528
1513
1511
1838
9.19
6.98
0.49
8
1
51
-
1561
7.81
6.58
0.47
6
1
52
-15.1%
444
2.22
1.92
0.14
2
1
17
-75.8%
888
4.44
4.32
0.31
3
1
34
-51.7%
762
3.81
2.58
0.18
3
1
15
-58.5%
4460
22.30
20.21
1.43
16
1
129
142.7%
103
include gB failed to cause any cell fusion (data not shown). Similarly, inclusion of p91537 in the transfection fusion assay failed to cause any cell fusion (Fig. 2.2C). Plasmid
p9-1537 specifies gB containing an internal, in-frame truncation spanning amino acids
130-697. This deletion causes rapid degradation of the gB product (Baghian et al., 1993).
All gB mutants, which modified H17b used in these experiments expressed high levels of
gB and similar amounts of gB were detected on cell surfaces by immunofluorescence
against paraformaldehyde fixed transfected cells (Baghian et al., 1993; data not shown).
Transfection with p9-1511 caused more than 2-fold increase in cell fusion in comparison
to wild-type gB (Fig. 2.2F, Table 2.1). An identical truncation of 28 amino acids in gB
specified by mutant virus amb1511 caused extensive cell fusion (Baghian et al., 1993).
Interestingly, transfection of the 1511(gB∆28) plasmid mixture resulted in fusion of the
entire cell monolayer (8 x 106 cells) within 48-72 hours after transfection (data not
shown).
Transfections with p9-1513 or p9-1528 specifying gB truncations of 64 and 49
amino acids, respectively, both of which disrupt the predicted α-helical region H17a,
caused substantially less cell fusion (approximately 50% reduction) than that of the wildtype gB gene (Fig. 2.2 D, E, Table 2.1). Previously, we showed that gB specified by p91513 and p9-1528 was not as efficiently processed through the Golgi as the gB specified
by p9-2080 (wild type) or p9-1511 (Baghian et al., 1993). Therefore, the inability of
these two mutant forms of gB to cause cell fusion may be due to their altered processing
and transport to cell surfaces.
104
The Role of α-helical Domain H17b in Cell Fusion
The 1511 gB truncation interrupted the predicted α−helical domain H17b,
allowing only the first half of this domain to be expressed (Fig. 2.1) (Baghian et al.,
1993; Pellett et al., 1985). To further assess the role of this domain in the transient fusion
assay, additional mutated gB genes were constructed and tested for their ability to cause
cell fusion. Plasmid pCMV-gB(∆36) specifies a gB truncated immediately before the
H17b domain, while plasmid pCMV-gB(∆16) specifies gB truncated after the H17b
domain (Fig. 2.1). The gB(∆16) truncation produced fusion similar to wild-type gB (Fig.
2.4A, C). In contrast, gB(∆36) caused similar levels of cell fusion to gB(∆28) (Fig. 2.4:
D, E). To evaluate the role of the predicted α-helical structure of the H17b domain in cell
fusion, a single amino acid replacement was engineered into the H17b domain by
replacing alanine 874 with proline, causing disruption of the predicted α−helical structure
(Fig. 2.1). Expression of gB(A874P) produced cell fusion similar to gB(∆28) and
gB(∆36) (Fig. 2.4: D, E, F). For comparative purposes, we tested the ability of gB
specified by syncytial mutant HFEM(tsB5), referred to as gB(syn3), which contains a
missense syncytial mutation within H17a, to cause cell fusion. Transient expression of
the gB(syn3) gene did not increase cell fusion relative to the wild-type gB, in agreement
with results obtained with other HSV-1 gB syncytial mutations (Turner et al., 1998)
(Fig. 2.4: A, B). Polykaryocyte formation was quantified by counting propidium iodide
stained nuclei as explained earlier. Results are summarized in Table 2.2A.
The Effect of Heparin on gB-Mediated Cell Fusion
Heparin is a potent inhibitor of virus penetration as well as gB-mediated (syn3)
virus induced cell fusion (Gruenheid et al., 1993; Herold et al., 1994; Herold et al., 1991;
105
Figure 2.4: gB-H17b-targeted mutations enhance fusion of transfected Cos cells.
Cells transfected with plasmids expressing gD, gH/gL, and A: gB, B: gB(syn3), C:
gB(∆16), D: gB(∆28), E: gB (∆36), and F: gB (A874P). Transfected cells were visualized
by IFA at 36 hours after transfection at 65X magnification.
106
Figure 2.5: gB-H17b-targeted mutations confer resistance to heparin inhibition of
cell fusion. Cells transfected with plasmids expressing gD, gH/gL, and A: gB, B:
gB(syn3), C: gB(∆16), D: gB(∆28), E: gB (∆36), and F: gB (A874P). Cells were
incubated in the presence of heparin. Transfected cells were visualized by IFA at 36
hours after transfection at 65X magnification.
107
Nuclei per Transfected Cell
gB
gB (syn3)
gB (∆16)
gB (∆28)
gB (∆36)
gB (A874P)
Plasmids Transfected (all include gD, gHgL)
Figure 2.6: Quantitative comparison of fusion of transfected cells in the presence
and absence of heparin. Cell fusion represented in figure 4 (without heparin) and figure
2.5 (with heparin) was quantitated as described in the legend of figure 3.
108
Table 2.2A: gB-H17b-targeted mutations enhance fusion of transfected Cos cells.
Plasmids Transfected
(all include gD, gHgL)
Total nuclei counted
Average nuclei
Standard Deviation
Standard Error
Median
Smallest # of nuclei
Largest # of nuclei
% Increase over wt gB
gB
gB(syn3)
gB(∆16)
gB(∆28)
gB(∆36)
gB (A874P)
2035
10.18
6.93
0.49
9
1
41
-
2340
11.7
8.61
0.61
11
1
44
14.9%
2232
11.16
7.81
0.55
10
1
47
9.6%
4146
20.73
15.62
1.1
17
1
92
103.6%
4332
21.66
17.36
1.23
18
1
99
112.8%
4000
20
14.33
1.01
18
1
80
96.5%
Table 2.2B: gB-H17b-targeted mutations confer resistance to heparin inhibition of
cell fusion.
Plasmids Transfected
(all include gD, gHgL)
Total nuclei counted
Average nuclei
Standard Deviation
Standard Error
Median
Smallest # of nuclei
Largest # of nuclei
% Inhibition by Heparin
gB
gB(syn3)
gB(∆16)
gB(∆28)
gB(∆36)
gB (A874P)
471
2.36
1.93
0.13
2
1
10
85.2%
486
2.43
2.14
0.15
2
1
14
86.6%
1059
5.3
5.07
1.1
3
1
26
57.7%
3414
17.07
10.95
0.77
16
1
55
18.6%
3397
16.99
14.04
0.99
13
1
91
22.6%
2543
12.72
9.31
0.66
11
1
50
38.3%
109
Shieh et al., 1992; WuDunn and Spear, 1989). Transfected cell monolayers were
incubated either in the presence or absence of heparin (10 mg/ml). Cell fusion was
visualized through fluorescence microscopy and the levels of observed cell fusion was
quantified (Figs. 2.5, 2.6). Incubation of transfected cell monolayers with heparin
inhibited both wild-type gB and gB(syn3)-mediated cell fusion (~85% inhibition) (Figs.
2.4, 2.5: A, B; Table 2.2B) as previously reported for tsB5 virus-induced cell fusion
(Seck et al., 1994). In contrast, fusion of cells expressing gB(∆28) or gB(∆36) was
largely resistant to heparin inhibition (Figs. 2.4, 2.5: D, E). Cell fusion caused by
gB(A874P) was inhibited by approximately 40% in heparin treated versus untreated
controls (Fig. 2.4, 2.5:F, Table 2.2B). Heparin inhibited fusion caused by gB(∆16) by
approximately 58% (Figs. 2.4, 2.5: C; Table 2.2B).
The Effect of Heparin on Virus-Induced Cell Fusion
Heparin was unable to fully inhibit cell fusion mediated by truncated or mutated
gB in the transient fusion assay. Therefore, we assayed the effect of heparin on virusinduced cell fusion caused by syncytial mutant viruses. Syncytial viruses mp(MP),
HFEM(tsB5), and amb1511 were engineered to constitutively express EGFP as described
previously for the construction of the KOS/EGFP virus (Foster, Chouljenko, and
Kousoulas, 1999). Expression of EGFP from this locus results in distinct nuclear and
diffuse cytoplasmic fluorescence, facilitating visualization of polykaryocytes (Fig. 2.7).
As previously reported (Seck et al., 1994), tsB5-induced cell fusion was inhibited by
heparin (Fig. 2.7: C, D). In contrast fusion caused by mp(MP), which contains the syn1
mutation in glycoprotein K (gK) was not inhibited by heparin (Fig. 2.7: G, H). Similar to
110
the results obtained in the transient fusion assay, cell fusion caused by mutant virusamb1511/EGFP was resistant to heparin inhibition (Fig. 2.7: E, F).
DISCUSSION
Studies on the role of individual viral glycoproteins in virus entry as well as virusinduced cell fusion are hampered by the fact that multiple glycoproteins and proteins are
involved in both processes. In this regard, the established model system for virusassociated membrane fusion phenomena via transient co-expression of gB, gD, and
gH/gL represents a reasonable model system to investigate individual glycoprotein
functions. However, it is important to compare virus-induced cell fusion and virus-free
fusion systems to ensure that functional aspects of glycoproteins expressed in the
transfection fusion system represent glycoprotein functions manifested in the context of
the viral genome. Our investigations produced the following conclusions. First, we show
that the predicted α−helical domain H17b within the carboxyl-terminus of gB is involved
in both virus-induced and virus-free fusion systems. Second, heparin is a specific
inhibitor of gB-mediated fusion in both systems. Third, resistance to heparin inhibition of
gB-mediated cell fusion is associated with the predicted α−helical structure H17b. Thus,
gB-mediated fusion is functionally similar for both fusion assays.
Previously, our laboratory showed that truncating gB by 28 amino acids produced
wild type levels of mature gB and caused extensive virus-induced cell fusion (Baghian et
al., 1993), while a truncation of 41 amino acids did not affect virus replication and failed
to cause cell fusion (Huff et al., 1988). It was proposed that the α-helical domain H17b,
which was interrupted by the 28 amino acid truncation, must be involved in virus-induced
cell fusion (Baghian et al., 1993). Co-expression of the 28 amino acid truncated gB
111
Figure 2.7: Cell fusion caused by viruses specifying H17b mutated gB is resistant to
heparin inhibition. Vero cells were infected at an MOI of 0.1 with KOS/EGFP (A, B),
tsb5/EGFP (C, D), amb1511/EGFP (E,F) and MP/EGFP (G, H) in the presence (B, D, F,
H) or absence (A, C, E, G) of heparin. Infected cells were visualized by fluorescence
microscopy (65X magnification).
112
specified by plasmid p9-1511 with gD, and gH/gL caused extensive cell fusion as
evidenced by the formation of large-size polykaryocytes. The extent of the fusion
occurring after cotransfection of glycoprotein genes containing the truncated gB was
similar to virus-induced cell fusion caused by mutant viruses expressing the gB(syn3) or
gB(∆28). Ultimately, transfection of Cos cells led to fusion of the entire cell monolayer
of up to 5 x 106 cells into a single syncytium within 48 to 72 hours. Clearly, the 28 amino
acid gB deletion functioned similarly in both the virus-induced cell fusion and virus-free
transfection fusion assay. Three conclusions can be made based on these results. First,
the ability of truncated gB to cause extensive cell fusion in the transfection fusion assay
provides further evidence that certain gB functions can be manifested in the absence of
productive viral replication. Secondly, the truncated gB's ability to extensively fuse
membranes requires only the presence of co-expressed gD and gH/gL glycoproteins.
Thirdly, the domains of gB that are responsible for this extensive cell fusion phenotype,
in both virus-induced and virus-free transfection systems, are located in the carboxylterminus of gB.
In agreement with our previous results, which implicated the H17b α-helical
domain in virus-induced cell fusion, we found that deletion of the entire H17b domain or
substitution of a single proline within the H17b helical domain caused extensive cell
fusion as evidenced by the formation of large-size polykaryocytes. The ability of the two
gB truncations and the single proline residue substitution to cause extensive cell fusion
indicates that the predicted α−helical structure H17b serves as a major determinant for
membrane fusion. All gB mutants, which disrupted the predicted α-helical domain H17 b
either by truncation or the proline substitution did not affect gB synthesis or transport to
113
cell surfaces as evidenced by immunofluorescence against transfected cells fixed with 4%
paraformaldehyde (not shown). Therefore, the ability of these mutant gB forms to cause
cell fusion is largely due to their specific gB mutations. H17b is located immediately
adjacent to the predicted 31 amino acid-long α-helix, H17a, within which multiple
syncytial mutations have been mapped (Bzik et al., 1984a; Gage, Levine, and Glorioso,
1993). However, syncytial mutations within H17a and H17b can be functionally
differentiated with respect to their ability to cause membrane fusion in the transfection
fusion system. The syn3 mutation within H17a failed to enhance fusion in the transfection
fusion assay. In contrast, both the 36 and 28 amino acid truncations as well as the proline
substitution affecting H17b readily caused extensive cell fusion. The most plausible
explanation for these results is that H17b functions as an independent fusion domain
regulator. Partial or full inactivation of this domain confers the ability on gB to cause
extensive cell fusion. Furthermore, fusion associated with this domain requires the
presence of only gD and gH/gL. In contrast, the H17a domain must require additional
viral proteins expressed during viral infection. Truncations and mutations in the gH
carboxyl terminus have been shown to inhibit cell fusion implying that the carboxylterminus of gH is also involved in membrane fusion (Browne, Bruun, and Minson, 1996;
Wilson, Davis-Poynter, and Minson, 1994). An alternative explanation of the role of the
H17b domain in cell fusion is that the carboxyl-termini of gB and gH functionally
interact and this interaction affects the fusogenic abilities of both gB and gH. It is worth
noting that there is a discrepancy between trasfection fusion results obtained with HSV-1
and HSV-2 glycoproteins, inasmuch as a syn mutation within HSV-2 gB was reported to
enhance fusion in the transfection fusion system (Muggeridge, 2000). HSV-2 enters
114
faster into cells than HSV-1 implying the presence of an enhanced mechanism for virusto-cell fusion (DeLuca, 1981, Kousoulas, unpublished observations). Therefore, the
different dynamics of HSV-1 and HSV-2 glycoprotein-mediated membrane fusion
phenomena may account for the observed differences in fusion caused by syncytial
mutations within HSV -1 and HSV-2 gB.
Heparin is a specific inhibitor of gB-
mediated virus-induced cell fusion (Seck et al., 1994). In agreement to previous reports,
we confirmed that heparin specifically inhibited fusion caused by syncytial mutant
HFEM (tsB5), while it did not inhibit fusion caused by mutants with syncytial mutations
in gK. Heparin is known to predominantly bind to gC and secondarily to gB (Herold et
al., 1994; Herold et al., 1991; Laquerre et al., 1998; Spear et al., 1992). The binding sites
of other proteins exhibiting heparin-binding activity have been mapped to dense regions
of positively charged residues containing variable lengths of arginine-rich and/or lysine
rich regions (Cardin et al., 1991a; Cardin et al., 1991b; Marino et al., 1999; Sendak and
Bensadoun, 1998; Sendak et al., 2000; Wong et al., 1995). The primary sequence of gB
contains a heparin binding site at amino acids 68 to 76 (KPKKNKKPK) in the amino
terminus of gB. Deletion of this amino acid domain results in substantial reduction in
heparin binding of gB (Laquerre et al., 1998). There are additional potential heparin
binding sites within gB at amino acid locations 158, 318, 494, 862 and 882 containing the
consensus binding motif R/K-X-R/K. The simplest explanation for the inhibitory effect of
heparin on gB-mediated cell fusion is that heparin physically blocks cell fusion. Heparin
inhibited fusion in the transfection fusion system when full-copy gB, gD, and gH/gL
were co-expressed or when transfections contained the gB(syn3)gene. However, heparin
was unable to substantially inhibit fusion caused by truncations of gB that delete all or
115
part of the H17b α−helix. Heparin did partially inhibit fusion caused by the single Ala to
Pro substitution, which inactivates the predicted α−helical structure of the H17b domain.
These differences between the truncated gB and the proline substituted gB could be due
to the more severe nature of the truncations versus the single amino acid replacement in
that certain amino acids upstream or downstream of the proline substitution may confer
partial susceptibility to heparin inhibition of membrane fusion.
We offer the following hypotheses to explain the effect of heparin on gBmediated cell fusion. First, truncation of gB may affect the conformation of the gB amino
terminus resulting in the partial loss of heparin binding. Secondly, gB truncations may
cause changes in the interaction of gB with cellular receptors, which render gB-mediated
fusion resistant to heparin. In support of the latter hypothesis, it was shown that bovine
herpesvirus gB interacted with two types of receptors on Madin Darby bovine kidney
cells (MDBK), a heparan sulfate proteoglycan and an unknown high affinity receptor,
respectively. Truncated gB retained the ability to bind heparin, but not the ability to block
the interaction of gB with a high affinity binding receptor on MDBK cells (Li et al.,
1996). Furthermore, deletion of the carboxyl terminus of gB resulted in the loss of one
monoclonal antibody epitope located in the amino terminal of gB, implying that the
truncated gB had assumed an altered conformation, which was responsible for the loss of
the binding to the high-affinity receptor (Li et al., 1996). Additional evidence that
perturbations of the carboxyl terminus of gB may affect the conformation of the gB
extracellular portion was provided by an Ala to Val change at position 851 within the
syncytial region II of HSV-1 gB, which caused delayed viral entry kinetics (Gage,
Levine, and Glorioso, 1993).
116
Substantial differences between the virus-induced and transfection-induced fusion
systems exist with respect to the role of heparan sulfate moieties on cell surfaces.
Heparan sulfate enhanced virus-induced cell fusion, inasmuch as viruses with syncytial
mutations failed to readily fuse heparan sulfate deficient cells. In contrast, fusion caused
by transient transfection of viral glycoproteins did not require the presence of heparan
sulfate, although it did require the presence of cell surface entry receptors specific for gD
(Pertel et al., 2001; Shieh and Spear, 1994). Therefore, the ability of heparin to inhibit
both virus-induced cell fusion and virus-free fusion in the transfection fusion assay must
reflect direct inhibition of gB-associated functions and not heparin competitive inhibition
of gB binding to heparan sulfate proteoglycans on apposed cell surfaces. Such
interactions presumably, would strengthen interactions between plasma membranes and
facilitate membrane fusion.
Heparin is known to bind to a variety of important proteins on cell surfaces and
modulate the functions of these proteins predominantly through signal transduction
effects to intracellular targets. However, heparin can also internalize into cells via
specific or nonspecific receptors (Barzu et al., 1996; Heinzelmann et al., 1999;
Letourneur, Caleb, and Castellot, 1995a; Letourneur, Caleb, and Castellot, 1995b;
Watanabe, Haba, and Yuasa, 1995). Therefore, it is possible that endocytosed heparin
may directly bind to the carboxyl terminus of gB at the predicted sites located at amino
acids 862 and 882. In this scenario, resistance to heparin inhibition of cell fusion by
truncated gB may be due to the lack of heparin binding sites, particularly the site at
amino acid 882, which is deleted in both gB truncations. The reduced ability of heparin to
inhibit cell fusion caused by gB(A874P) and gB(∆16) may be the result of local
117
conformational changes, which prevent binding of heparin to upstream binding sites.
Additional experiments are needed to ascertain whether heparin readily enters into gBtransfected cells and whether the carboxyl terminus of gB is able to bind heparin.
A carboxyl terminal truncation of the PRV gB enhanced both virus-induced and
virus-free cell fusion (Klupp, Nixdorf, and Mettenleiter, 2000; Nixdorf et al., 2000).
However, in the virus-free transfection assay cell fusion can occur in the absence of gD.
Furthermore, a hybrid gD/gH protein composed of the extracellular portion of gD fused
in-frame to a portion of gH, which contains the carboxyl terminus of gH including its
membrane spanning region, functionally substituted the gH/gL heterodimer (Klupp and
Mettenleiter, 1999; Klupp, Nixdorf, and Mettenleiter, 2000). We constructed and tested a
similar HSV-1 gD/gH fusion protein and found that it failed to substitute for the presence
of gD or gH/gL in the virus-free fusion system (not shown). Thus, it is clear that there are
similarities between PRV and HSV-1 with respect to the role of the gB carboxyl terminus
in cell fusion, but also substantial differences with respect to the functions of gD and
gH/gL. In this regard, the apparent disparity between the two fusion systems may reflect
fundamental differences in the functions of these proteins as well as differences in their
interactions with cellular receptors (Nixdorf et al., 1999).
REFERENCES
Addison, C., Rixon, F. J., Palfreyman, J. W., O'Hara, M., and Preston, V. G. (1984).
Characterisation of a herpes simplex virus type 1 mutant which has a temperaturesensitive defect in penetration of cells and assembly of capsids. Virology 138,
246-59.
Baghian, A., Huang, L., Newman, S., Jayachandra, S., and Kousoulas, K. G. (1993).
Truncation of the carboxy-terminal 28 amino acids of glycoprotein B specified by
herpes simplex virus type 1 mutant amb1511-7 causes extensive cell fusion. J
Virol 67, 2396-401.
118
Baines, J. D., Ward, P. L., Campadelli-Fiume, G., and Roizman, B. (1991). The UL20
gene of herpes simplex virus 1 encodes a function necessary for viral egress. J
Virol 65, 6414-24.
Barzu, T., Pascal, M., Maman, M., Roque, C., Lafont, F., and Rousselet, A. (1996). Entry
and distribution of fluorescent antiproliferative heparin derivatives into rat
vascular smooth muscle cells: comparison between heparin-sensitive and heparinresistant cultures. J Cell Physiol 167, 8-21.
Bond, V. C., and Person, S. (1984). Fine structure physical map locations of alterations
that affect cell fusion in herpes simplex virus type 1. Virology 132, 368-76.
Browne, H. M., Bruun, B. C., and Minson, A. C. (1996). Characterization of herpes
simplex virus type 1 recombinants with mutations in the cytoplasmic tail of
glycoprotein H. J Gen Virol 77, 2569-73.
Bzik, D. J., Fox, B. A., DeLuca, N. A., and Person, S. (1984a). Nucleotide sequence of a
region of the herpes simplex virus type 1 gB glycoprotein gene: mutations
affecting rate of virus entry and cell fusion. Virology 137, 185-90.
Bzik, D. J., Fox, B. A., DeLuca, N. A., and Person, S. (1984b). Nucleotide sequence
specifying the glycoprotein gene, gB, of herpes simplex virus type 1. Virology
133, 301-14.
Cai, W. H., Gu, B., and Person, S. (1988). Role of glycoprotein B of herpes simplex virus
type 1 in viral entry and cell fusion. J Virol 62, 2596-604.
Cai, W. Z., Person, S., DebRoy, C., and Gu, B. H. (1988). Functional regions and
structural features of the gB glycoprotein of herpes simplex virus type 1. An
analysis of linker insertion mutants. J Mol Biol 201, 575-88.
Cardin, A. D., Demeter, D. A., Weintraub, H. J., and Jackson, R. L. (1991a). Molecular
design and modeling of protein-heparin interactions. Methods Enzymol 203, 55683.
Cardin, A. D., Demeter, D. A., Weintraub, H. J., and Jackson, R. L. (1991b). Molecular
design and modeling of protein-heparin interactions. Methods Enzymol 203, 55683.
Cocchi, F., Menotti, L., Mirandola, P., Lopez, M., and Campadelli-Fiume, G. (1998). The
ectodomain of a novel member of the immunoglobulin subfamily related to the
poliovirus receptor has the attributes of a bona fide receptor for herpes simplex
virus types 1 and 2 in human cells. J Virol 72, 9992-10002.
119
Davis-Poynter, N., Bell, S., Minson, T., and Browne, H. (1994). Analysis of the
contributions of herpes simplex virus type 1 membrane proteins to the induction
of cell-cell fusion. J Virol 68, 7586-90.
Debroy, C., Pederson, N., and Person, S. (1985). Nucleotide sequence of a herpes
simplex virus type 1 gene that causes cell fusion. Virology 145, 36-48.
DeLuca, N., Bzik, D. J., Bond, V. C., Person, S., and Snipes, W. (1982). Nucleotide
sequences of herpes simplex virus type 1 (HSV-1) affecting virus entry, cell
fusion, and production of glycoprotein gb (VP7). Virology 122, 411-23.
Dolter, K. E., Ramaswamy, R., and Holland, T. C. (1994). Syncytial mutations in the
herpes simplex virus type 1 gK (UL53) gene occur in two distinct domains. J
Virol 68, 8277-81.
Engel, J. P., Boyer, E. P., and Goodman, J. L. (1993). Two novel single amino acid
syncytial mutations in the carboxy terminus of glycoprotein B of herpes simplex
virus type 1 confer a unique pathogenic phenotype. Virology 192, 112-20.
Forrester, A., Farrell, H., Wilkinson, G., Kaye, J., Davis-Poynter, N., and Minson, T.
(1992). Construction and properties of a mutant of herpes simplex virus type 1
with glycoprotein H coding sequences deleted. J Virol 66, 341-8.
Foster, T. P., Chouljenko, V. N., and Kousoulas, K. G. (1999). Functional
characterization of the HveA homolog specified by African green monkey kidney
cells with a herpes simplex virus expressing the green fluorescence protein.
Virology 258, 365-74.
Gage, P. J., Levine, M., and Glorioso, J. C. (1993). Syncytium-inducing mutations
localize to two discrete regions within the cytoplasmic domain of herpes simplex
virus type 1 glycoprotein B. J Virol 67, 2191-201.
Geraghty, R. J., Krummenacher, C., Cohen, G. H., Eisenberg, R. J., and Spear, P. G.
(1998). Entry of alphaherpesviruses mediated by poliovirus receptor-related
protein 1 and poliovirus receptor. Science 280, 1618-20.
Gruenheid, S., Gatzke, L., Meadows, H., and Tufaro, F. (1993). Herpes simplex virus
infection and propagation in a mouse L cell mutant lacking heparan sulfate
proteoglycans. J Virol 67, 93-100.
Haanes, E. J., Nelson, C. M., Soule, C. L., and Goodman, J. L. (1994). The UL45 gene
product is required for herpes simplex virus type 1 glycoprotein B-induced fusion.
J Virol 68, 5825-34.
Heinzelmann, M., Platz, A., Flodgaard, H., Polk, H. C., Jr., and Miller, F. N. (1999).
Endocytosis of heparin-binding protein (CAP37) is essential for the enhancement
120
of lipopolysaccharide-induced TNF-alpha production in human monocytes. J
Immunol 162, 4240-5.
Herold, B. C., Visalli, R. J., Susmarski, N., Brandt, C. R., and Spear, P. G. (1994).
Glycoprotein C-independent binding of herpes simplex virus to cells requires cell
surface heparan sulphate and glycoprotein B. J Gen Virol 75, 1211-22.
Herold, B. C., WuDunn, D., Soltys, N., and Spear, P. G. (1991). Glycoprotein C of
herpes simplex virus type 1 plays a principal role in the adsorption of virus to
cells and in infectivity. J Virol 65, 1090-8.
Huff, V., Cai, W., Glorioso, J. C., and Levine, M. (1988). The carboxy-terminal 41 amino
acids of herpes simplex virus type 1 glycoprotein B are not essential for
production of infectious virus particles. J Virol 62, 4403-6.
Hutchinson, L., Goldsmith, K., Snoddy, D., Ghosh, H., Graham, F. L., and Johnson, D.
C. (1992). Identification and characterization of a novel herpes simplex virus
glycoprotein, gK, involved in cell fusion. J Virol 66, 5603-9.
Jacobson, J. G., Martin, S. L., and Coen, D. M. (1989). A conserved open reading frame
that overlaps the herpes simplex virus thymidine kinase gene is important for viral
growth in cell culture. J Virol 63, 1839-43.
Klupp, B. G., and Mettenleiter, T. C. (1999). Glycoprotein gL-independent infectivity of
pseudorabies virus is mediated by a gD-gH fusion protein. J Virol 73, 3014-22.
Klupp, B. G., Nixdorf, R., and Mettenleiter, T. C. (2000). Pseudorabies virus
glycoprotein M inhibits membrane fusion. J Virol 74, 6760-8.
Laquerre, S., Argnani, R., Anderson, D. B., Zucchini, S., Manservigi, R., and Glorioso, J.
C. (1998). Heparan sulfate proteoglycan binding by herpes simplex virus type 1
glycoproteins B and C, which differ in their contributions to virus attachment,
penetration, and cell-to-cell spread. J Virol 72, 6119-30.
Letourneur, D., Caleb, B. L., and Castellot, J. J., Jr. (1995a). Heparin binding,
internalization, and metabolism in vascular smooth muscle cells: I. Upregulation
of heparin binding correlates with antiproliferative activity. J Cell Physiol 165,
676-86.
Letourneur, D., Caleb, B. L., and Castellot, J. J., Jr. (1995b). Heparin binding,
internalization, and metabolism in vascular smooth muscle cells: II. Degradation
and secretion in sensitive and resistant cells. J Cell Physiol 165, 687-95.
Li, Y., van Drunen Littel-van den Hurk, S., Liang, X., and Babiuk, L. A. (1996). The
cytoplasmic domain of bovine herpesvirus 1 glycoprotein B is important for
121
maintaining conformation and the high-affinity binding site of gB. Virology 222,
262-8.
Ligas, M. W., and Johnson, D. C. (1988). A herpes simplex virus mutant in which
glycoprotein D sequences are replaced by beta-galactosidase sequences binds to
but is unable to penetrate into cells. J Virol 62, 1486-94.
Manservigi, R., Spear, P. G., and Buchan, A. (1977). Cell fusion induced by herpes
simplex virus is promoted and suppressed by different viral glycoproteins. Proc
Natl Acad Sci U S A 74, 3913-7.
Marino, M., Friedlander, J. A., McCluskey, R. T., and Andrews, D. (1999). Identification
of a heparin-binding region of rat thyroglobulin involved in megalin binding. J
Biol Chem 274, 30377-86.
Mettenleiter, T. C. (2000). Aujeszky's disease (pseudorabies) virus: the virus and
molecular pathogenesis--state of the art, June 1999. Vet Res 31, 99-115.
Montgomery, R. I., Warner, M. S., Lum, B. J., and Spear, P. G. (1996). Herpes simplex
virus-1 entry into cells mediated by a novel member of the TNF/NGF receptor
family. Cell 87, 427-36.
Muggeridge, M. I. (2000). Characterization of cell-cell fusion mediated by herpes
simplex virus 2 glycoproteins gB, gD, gH and gL in transfected cells. J Gen Virol
81, 2017-27.
Nixdorf, R., Klupp, B. G., Karger, A., and Mettenleiter, T. C. (2000). Effects of
truncation of the carboxy terminus of pseudorabies virus glycoprotein B on
infectivity. J Virol 74, 7137-45.
Nixdorf, R., Schmidt, J., Karger, A., and Mettenleiter, T. C. (1999). Infection of Chinese
hamster ovary cells by pseudorabies virus. J Virol 73, 8019-26.
Novotny, M. J., Parish, M. L., and Spear, P. G. (1996). Variability of herpes simplex
virus 1 gL and anti-gL antibodies that inhibit cell fusion but not viral infectivity.
Virology 221, 1-13.
Pellett, P. E., Kousoulas, K. G., Pereira, L., and Roizman, B. (1985). Anatomy of the
herpes simplex virus 1 strain F glycoprotein B gene: primary sequence and
predicted protein structure of the wild type and of monoclonal antibody-resistant
mutants. J Virol 53, 243-53.
Pertel, P. E., Fridberg, A., Parish, M. L., and Spear, P. G. (2001). Cell Fusion Induced by
Herpes Simplex Virus Glycoproteins gB, gD, and gH-gL Requires a gD Receptor
but Not Necessarily Heparan Sulfate. Virology 279, 313-324.
122
Pertel, P. E., and Spear, P. G. (1996). Modified entry and syncytium formation by herpes
simplex virus type 1 mutants selected for resistance to heparin inhibition.
Virology 226, 22-33.
Pogue-Geile, K. L., and Spear, P. G. (1987). The single base pair substitution responsible
for the Syn phenotype of herpes simplex virus type 1, strain MP. Virology 157,
67-74.
Roop, C., Hutchinson, L., and Johnson, D. C. (1993). A mutant herpes simplex virus type
1 unable to express glycoprotein L cannot enter cells, and its particles lack
glycoprotein H. J Virol 67, 2285-97.
Ruyechan, W. T., Morse, L. S., Knipe, D. M., and Roizman, B. (1979). Molecular
genetics of herpes simplex virus. II. Mapping of the major viral glycoproteins and
of the genetic loci specifying the social behavior of infected cells. J Virol 29, 67797.
Sanders, P. G., Wilkie, N. M., and Davison, A. J. (1982). Thymidine kinase deletion
mutants of herpes simplex virus type 1. J Gen Virol 63, 277-95.
Seck, T., Lingen, M., Weise, K., and Falke, D. (1994). Evidence for a multistep
mechanism for cell-cell fusion by herpes simplex virus with mutations in the syn
3 locus using heparin derivatives during fusion from within. Arch Virol 136, 17381.
Sendak, R. A., and Bensadoun, A. (1998). Identification of a heparin-binding domain in
the distal carboxyl- terminal region of lipoprotein lipase by site-directed
mutagenesis. J Lipid Res 39, 1310-5.
Sendak, R. A., Berryman, D. E., Gellman, G., Melford, K., and Bensadoun, A. (2000).
Binding of hepatic lipase to heparin. Identification of specific heparin-binding
residues in two distinct positive charge clusters. J Lipid Res 41, 260-8.
Shieh, M. T., and Spear, P. G. (1994). Herpesvirus-induced cell fusion that is dependent
on cell surface heparan sulfate or soluble heparin. J Virol 68, 1224-8.
Shieh, M. T., WuDunn, D., Montgomery, R. I., Esko, J. D., and Spear, P. G. (1992). Cell
surface receptors for herpes simplex virus are heparan sulfate proteoglycans. J
Cell Biol 116, 1273-81.
Spear, P. G. (1993). Entry of alphaherpesviruses into cells. Seminars in Virology 4, 167180.
Spear, P. G., Shieh, M. T., Herold, B. C., WuDunn, D., and Koshy, T. I. (1992). Heparan
sulfate glycosaminoglycans as primary cell surface receptors for herpes simplex
virus. Adv Exp Med Biol 313, 341-53.
123
Terry-Allison, T., Montgomery, R. I., Whitbeck, J. C., Xu, R., Cohen, G. H., Eisenberg,
R. J., and Spear, P. G. (1998). HveA (herpesvirus entry mediator A), a coreceptor
for herpes simplex virus entry, also participates in virus-induced cell fusion. J
Virol 72, 5802-10.
Turner, A., Bruun, B., Minson, T., and Browne, H. (1998). Glycoproteins gB, gD, and
gHgL of herpes simplex virus type 1 are necessary and sufficient to mediate
membrane fusion in a Cos cell transfection system. J Virol 72, 873-5.
Visalli, R. J., and Brandt, C. R. (1991). The HSV-1 UL45 gene product is not required for
growth in Vero cells. Virology 185, 419-23.
Walev, I., Lingen, M., Lazzaro, M., Weise, K., and Falke, D. (1994). Cyclosporin A
resistance of herpes simplex virus-induced "fusion from within" as a phenotypical
marker of mutations in the Syn 3 locus of the glycoprotein B gene. Virus Genes 8,
83-6.
Warner, M. S., Geraghty, R. J., Martinez, W. M., Montgomery, R. I., Whitbeck, J. C.,
Xu, R., Eisenberg, R. J., Cohen, G. H., and Spear, P. G. (1998). A cell surface
protein with herpesvirus entry activity (HveB) confers susceptibility to infection
by mutants of herpes simplex virus type 1, herpes simplex virus type 2, and
pseudorabies virus. Virology 246, 179-89.
Watanabe, J., Haba, M., and Yuasa, H. (1995). Uptake of fractionated 3H-heparin by
isolated rat Kupffer cells. Pharm Res 12, 1092-5.
Weise, K., Kaerner, H. C., Glorioso, J., and Schroder, C. H. (1987). Replacement of
glycoprotein B gene sequences in herpes simplex virus type 1 strain ANG by
corresponding sequences of the strain KOS causes changes of plaque morphology
and neuropathogenicity. J Gen Virol 68, 1909-19.
Wilson, D. W., Davis-Poynter, N., and Minson, A. C. (1994). Mutations in the
cytoplasmic tail of herpes simplex virus glycoprotein H suppress cell fusion by a
syncytial strain. J Virol 68, 6985-93.
Wong, P., Hampton, B., Szylobryt, E., Gallagher, A. M., Jaye, M., and Burgess, W. H.
(1995). Analysis of putative heparin-binding domains of fibroblast growth factor1. Using site-directed mutagenesis and peptide analogues. J Biol Chem 270,
25805-11.
WuDunn, D., and Spear, P. G. (1989). Initial interaction of herpes simplex virus with
cells is binding to heparan sulfate. J Virol 63, 52-8.
124
CHAPTER III
HERPES SIMPLEX VIRUS TYPE 1 (HSV-1) GLYCOPROTEIN K
(gK) IS REQUIRED FOR GLYCOPROTEIN B (gB)-MEDIATED
VIRUS-INDUCED CELL FUSION, WHILE NEITHER gB AND gK
NOR gB AND UL20p FUNCTION REDUNDANTLY IN VIRION DEENVELOPMENT*
INTRODUCTION
Herpes simplex viruses specify at least eleven virally encoded glycoproteins, as
well as several non-glycosylated membrane associated proteins that function in several
important roles including virus entry via fusion of the viral envelope with cellular
membranes, intracellular virion morphogenesis and egress, cell-to-cell spread, and virusinduced cell fusion (Mettenleiter, 2002a; Mettenleiter, 2002b; Roizman and Sears, 2001;
Spear, 2004; Spear and Longnecker, 2003). Mutations that cause extensive virus-induced
cell-to-cell fusion have been mapped to at least four regions of the viral genome: the
UL20 gene (Baines et al., 1991; MacLean et al., 1991; Melancon, Foster, and Kousoulas,
2004), the UL24 gene (Jacobson et al., 1998; Sanders, Wilkie, and Davison, 1982), the
UL27 gene encoding glycoprotein B (gB) (Bzik et al., 1984; Pellett et al., 1985), and the
UL53 gene coding for glycoprotein K (gK) (Bond and Person, 1984; Debroy, Pederson,
and Person, 1985; Hutchinson et al., 1992; Pogue-Geile et al., 1984; Pogue-Geile and
Spear, 1987; Ryechan et al., 1979).
*
Accepted for publication in the Journal of Virology, MS # JVI01347-04 - Version 2, J. M. Melancon, R.
E. Luna, T. P. Foster, and K. G. Kousoulas, Herpes Simplex Virus Type 1 (HSV-1) Glycoprotein K (gK) is
Required for Glycoprotein B (gB)-mediated Virus-Induced Cell Fusion, while neither gB and gK nor gB
and UL20p Function Redundantly in Virion De-Envelopment, with permission from the American Society
for Microbiology.
125
The UL20 and UL53 (gK) genes encode multipass transmembrane proteins of 222
and 338 amino acids, respectively, which are conserved in all alphaherpesviruses
(Debroy, Pederson, and Person, 1985; MacLean et al., 1991; Ramaswamy and Holland,
1992). UL20p assumes a membrane topology that constitutes a mirror image of gK with
both amino and carboxyl termini of UL20p located intracellularly, while both amino and
carboxyl termini of gK are located extracellularly (Foster, Alvarez, and Kousoulas, 2003;
Foster et al., 2004b; Melancon, Foster, and Kousoulas, 2004). gK is known to be posttranslationally modified by N-linked carbohydrate addition, while UL20p is not
glycosylated (Debroy, Pederson, and Person, 1985; Hutchinson et al., 1992; Ramaswamy
and Holland, 1992). UL20p and gK localize to TGN membranes after endocytosis from
cell surfaces (Foster et al., 2004b). UL20p and gK are essential for cytoplasmic virion
morphogenesis, since mutant viruses lacking either gK or UL20p accumulate capsids
within the cytoplasm that are unable to acquire envelopes by budding into TGNassociated membranes (Baines et al., 1991; Foster and Kousoulas, 1999; Foster et al.,
2004a; Fuchs et al., 1997; Hutchinson and Johnson, 1995; Jayachandra, Baghian, and
Kousoulas, 1997; Melancon, Foster, and Kousoulas, 2004). Furthermore, UL20p is
essential for virus-induced cell fusion caused by either gB or gK syncytial mutations
(Foster et al., 2004a), and it is necessary for gK cell-surface expression (Foster, Alvarez,
and Kousoulas, 2003; Foster et al., 2004a; Foster et al., 2004b), strongly suggesting that
direct or indirect interactions between gK and UL20 are necessary for intracellular
protein trafficking, virus egress, and virus mediated cell-to-cell fusion (Dietz et al., 2000;
Foster, Alvarez, and Kousoulas, 2003; Foster et al., 2004a; Fuchs et al., 1997; Melancon,
Foster, and Kousoulas, 2004).
126
Glycoprotein B (gB) is highly conserved across all subfamilies of herpesviruses. It
exists within cells as a homodimeric type I integral membrane protein that is Nglycosylated at multiple sites (Claesson-Welsh and Spear, 1986; Claesson-Welsh and
Spear, 1987; Highlander et al., 1991; Laquerre, Person, and Glorioso, 1996; Roizman and
Sears, 2001; Spear, 1993). A variety of evidence indicates that gB plays important roles
in membrane fusion phenomena during virus entry and virus-induced cell fusion: HSV-1
mutant viruses lacking gB are not able to enter into cells (Cai et al., 1987) due to a postattachment defect that can be resolved by polyethelene glycol mediated fusion of viral
envelopes with cellular membranes (Cai, Gu, and Person, 1988); Single amino acid
substitutions and truncations of the carboxyl terminus of gB cause extensive virusinduced cell fusion (Baghian et al., 1993; Bzik et al., 1984; Cai et al., 1988; Gage,
Levine, and Glorioso, 1993); Transient co-expression of gB with gD, gH and gL causes
cell-to-cell fusion, which is substantially increased by carboxyl terminal truncations of
gB (Foster, Melancon, and Kousoulas, 2001; Haan, Lee, and Longnecker, 2001; Klupp,
Nixdorf, and Mettenleiter, 2000; Pertel, 2002). These results suggest a direct role for gB
in membrane fusion and suggest that perturbations of the carboxyl terminal domains of
gB facilitate gB-mediated cell-to-cell fusion.
It has been shown that syncytial mutations in gK failed to cause virus-induced cell
fusion in the absence of gB indicating that gB was essential for gK-associated cell-to-cell
fusion (Cai, Gu, and Person, 1988). In this paper, we utilized the HSV-1 (F) BAC system
to generate multiple mutant viruses lacking either gK, UL20, or gB, or combinations of
gK and gB, or UL20 and gB. Comparison of the BAC-generated insertion-deletion
mutants in the HSV-1(F) genetic background revealed subtle but important differences in
127
individual virus phenotypes in comparison to the laboratory adapted virus strain KOS,
most likely due to the limited passage of these viruses in cell culture. In addition, mutant
viruses were generated carrying syncytial mutations in gB in the absence of either gK or
UL20. Characterization of these viruses revealed that gK is necessary for gB-mediated
virus-induced cell fusion. Concurrent deletion of either gB and gK or gB and UL20p did
not prevent perinuclear virion de-envelopment indicating that gB and gK or gB and
UL20p do not function redundantly in mediating fusion between the viral envelopes and
the outer nuclear membrane.
MATERIALS AND METHODS
Cells and Viruses
African green monkey kidney (Vero) cells, CV-1 cells and 293 cells were
obtained from ATCC (Rockville, MD). Cells were maintained in Dulbecco’s modified
Eagle medium (GIBCO-BRL; Grand Island, N.Y.), supplemented with 10% fetal calf
serum (FCS) and antibiotics. Fd20-1 cells and D6 cells were maintained as described
previously (Cai et al., 1987; Melancon, Foster, and Kousoulas, 2004). The UL19 and
UL20 complementing cell line, G5, was a gift of P. Desai, (John Hopkins Medical
Center) (Desai et al., 1993). gKDIV5, gKsyn20DIV5, and tsB5 were as described
previously (Foster, Alvarez, and Kousoulas, 2003; Foster, Rybachuk, and Kousoulas,
2001). Flp-In-CV-1 cells were maintained as directed by the manufacturer (Invitrogen;
Carlsbad, CA).
Plasmids
Plasmid pcDNA5/FRT/gK was constructed by inserting the gK gene, PCR
amplified from KOS viral DNA, into pcDNA5/FRT/V5-His-TOPO (Invitrogen).
128
Plasmid p20F2 contains a 7,185 bp HSV-1 (KOS) genomic fragment spanning the region
comprising UL19, UL20, UL21, and UL22 and was described previously (Melancon,
Foster, and Kousoulas, 2004). Primers 5’CMVEGFP_20.5_BclI (5’TTCTTCTTCTTC
TGATCATGCTGACGTAAGTAGTTATTAATAGTAATCAATTACGGGG3’) and
3’CMVEGFP_BclI (5’CTCTTCCTCTCCTGATCAATGAGTTTGGACAAACCACA
ACTAGA3’) were used to PCR amplify a CMV-EGFP gene cassette from pTF9200
(Foster, Rybachuk, and Kousoulas, 1998). The resultant PCR product was digested with
BclI and ligated into p20F2 that was BamHI restricted and CIP treated. The resultant
plasmid, designated p∆1920-EGFP, contained a CMV-EGFP gene cassette, which
replaced the HSV-1 region in p20F2 contained within the BamHI sites at genomic
positions 39234 and 41549 (Genbank accession number NC_001806), removing the
entire UL20 ORF as well as the first 1293 bp of the UL19 ORF.
Construction of Transformed Flp-In-CV-1 Cell Lines
Generation of stable Flp-In-CV-1 expression cell lines was performed essentially
as directed by the manufacturer (Invitrogen) and as described previously (Melancon,
Foster, and Kousoulas, 2004). Confluent Flp-In-CV-1 monolayers in six well plates were
transfected with 0.3 µg of pcDNA5/FRT/gK and 2.7 µg of pOG44 (Invitrogen), which
expresses Flp recombinase, using Lipofectamine 2000. After 2 weeks, hygromycin B
(125µg/ml) resistant colonies were tested for the ability to complement the growth of
YEbac∆gK virus. All colonies tested complemented YEbac∆gK virus. Isolate FcgK-1,
derived from pcDNA5/FRT/gK transfected cells, was chosen for further use. FcgK-1
cells were used in generating YEbac∆gK virus stocks.
129
Construction of a Replication-Deficient Adenovirus Expressing gB
pAd5-Blue is a replication-defective, E1/E3 deleted Ad5-containing plasmid
(Moraes, Mayr, and Grubman, 2001). gB was PCR amplified from HSV-1 KOS viral
DNA using primers gB5’ClaI (5’-TCCTCCATCGATATGCGCCAGGGCGCCCCCG3’) and gB3’XbaI (5’-CTTCTTTCTAGATCACAGGTCGTCCTCGTCGGC-3’) and
transferred into the unique ClaI and XbaI sites located in the E1-deleted region of pAd5Blue, immediately downstream of the CMV promoter, to generate pAd5-gB. pAd5-gB
was transfected into 293 cells in a T-25 flask with Lipofectamine 2000, according to the
manufacturer’s directions (Invitrogen). 72 hours post-transfection, cell lysates were
prepared by multiple freeze/thaw cycles. A small portion of the cell lysate was then used
to infect a subsequent flask of 293 cells. After four passages, a high titer stock of Ad5gB was obtained, which was deficient for replication on Vero cells. Ad5-gB virus at
different MOIs was tested for its ability to complement YEbac∆gB on Vero cells, and an
MOI of 10 was found to be optimal.
Construction of HSV-1 Mutants with Deletions of the UL20, gB, and/or gK Genes
Insertion-deletion mutagenesis of pYEbac102 DNA was accomplished in
Escherichia coli using the λ gam, recE recT (GET) recombination system (Narayanan et
al., 1999; Orford et al., 2000), as described previously for mutagenesis of the KSHV
genome (Luna et al., 2004). Electrocompetent YEbac102 DH10B E. coli cells were
transformed with the plasmid pGETrec, which contains the genes encoding recE, recT
and bacteriophage λ gam, grown on plates containing chloramphenicol (12.5µg/ml) and
ampicillin (100µg/ml). Individual colonies were picked and grown overnight in LuriaBertani (LB) medium containing chloramphenicol and ampicillin. The next day, the
130
culture was inoculated into 250 ml of LB containing chloramphenicol and ampicillin until
a 0.4 optical density was reached at 600 nm. Addition of L-arabinose to a final
concentration of 0.2% (w/v) and further incubation of 40 minutes induced the expression
of the recE, recT and λ gam genes from the plasmid pGETrec. The cells were then
harvested and made electrocompetent. For the ∆UL20-GFPZeo mutation, a PCR
fragment containing a GFP-ZeocinR gene cassette flanked by ~50bp of viral sequences on
both sides was used for recombination to construct pYEbac∆UL20, containing the GFPZeocinR gene cassette within the targeted UL20 genomic region. For the ∆gB-Kan
mutation, a PCR fragment containing a kanamycinR gene cassette flanked by ~50bp of
viral sequences on both sides was used for recombination to construct pYEbac∆gB,
which contained the kanamycinR gene cassette within the targeted UL27 genomic region.
For the ∆gK-GFPZeo, a PCR fragment containing a GFP-ZeocinR gene cassette flanked
by ~50bp of viral sequences on both sides was used for recombination to construct
pYEbac∆gK, containing the GFP-ZeocinR gene cassette within the targeted UL53
genomic region. Briefly, 40µl of electrocompetent DH10B cells, harboring both
pYEbac102 and pGETrec, were electroporated with 200ng of each PCR product to delete
the target gene (gB, UL20, or gK) using standard electroporation parameters (1.8kV/cm,
200Ω and 25µF). Following electroporation, cells were grown in 1ml of LB for 60 min
and subsequently streaked onto LB agar plates containing chloramphenicol (12.5 µg/ml)
as well as either kanamycin (50µg/ml) or Zeocin (25µg/ml). Mutant pYEbac102 DNA
containing a deletion in the gB, UL20 or gK gene was isolated from bacterial colonies
and a second round of electroporation was performed to remove the plasmid pGETrec.
Following electroporation, cells were grown on agar plates containing chloramphenicol
131
as well as either kanamycin or Zeocin. Similar procedures were followed, starting with
electrocompetent DH10B E. coli cells harboring pYEbac∆gB, in order to generate
pYEbac∆gB∆gK and pYEbac∆gB∆UL20.
Confirmation of the Targeted Mutations in pYEbac102 DNA
HSV-1 BAC DNAs (pYEbac102, pYEbac∆gB, pYEbac∆UL20, pYEbac∆gK,
pYEbac∆gB∆UL20, and pYEbac∆gB∆gK) were purified from 500 ml of BAC cultures
using the Qiagen Large-Construct Kit (Qiagen; Valencia, CA). BAC DNA was digested
with KpnI, run on a 0.7% agarose gel, and transferred to charged nylon membranes
(BioRAD; Richmond, CA). Southern-blot hybridization was performed using either a 1.1
kb kanamycin or 800 bp GFP-Zeocin PCR product labeled with biotin (New England
Biolabs; Boston, MA). Chemiluminescence detection of the DNA was performed using
the North2South Chemiluminescent Hybridization and Detection Kit as described by the
manufacturer (Pierce Inc.; Rockford, IL). All mutations were sequenced to verify the
presence of the desired mutations in the BACs. To verify that no spurious mutations
were introduced into the viral genome of the individual BAC mutants during mutagenesis
or during the transfection procedure, each null mutation was rescued by homologous
recombination with DNA fragments spanning each gene. Each rescue virus was verified
to have similar phenotypic and replication characteristics as the wild-type BAC,
YEbac102.
Transfection of HSV-1 BAC DNAs
Transient transfection of cells with BAC DNAs was performed using
Lipofectamine 2000 (Invitrogen). Complementing cell lines were grown to 95%
confluency in 6 well plates. Cells were transfected with BAC DNA mixed with
132
Lipofectamine 2000 in Opti-MEM media as recommended by the manufacturer
(Invitrogen). After 6 hours of incubation at 37°C, the media was removed from the
transfected cells, cells were washed with PBS, and fresh DMEM media with 10% fetal
calf serum was added. At 72 hours post-transfection, virus stocks were collected.
Plaque Morphology of YEbac102 Mutants
For plaque morphology, Vero, Fd20-1, FcgK-1 and D6 cells were infected at
MOIs of 0.01 and 0.001 with the YEbac102, YEbac∆gB, YEbac∆UL20, YEbac∆gK,
YEbac∆gB∆gK and YEbac∆gB∆UL20 viruses and visualized by immunohistochemistry
at 48 hpi utilizing HRP conjugated anti-HSV antibodies (DAKO) and Novared
(VectorLabs) substrate development. For complementation of YEbac∆gB∆gK and
YEbac∆gB∆UL20 viruses, Fd20-1 and FcgK-1 were infected with Ad5-gB at an MOI of
10, 1 hour prior to HSV-1 infection. Due to the constitutive expression of gB in all cells,
plaques were visualized by immunohistochemistry at 48 hpi utilizing an anti-gD mAb
(RG-1103; Rumbaugh-Goodwin Institute; Plantation, FL) with the Vectastain Elite ABC
kit and Novared substrate development kit (VectorLabs; Burlingame, CA) according to
the manufacturer’s directions and as described previously (Foster, Alvarez, and
Kousoulas, 2003; Foster et al., 2004a).
One-step Growth Kinetics of YEbac102 Mutants
Analysis of one-step growth kinetics was as described previously (Foster,
Alvarez, and Kousoulas, 2003; Foster, Rybachuk, and Kousoulas, 2001). Briefly, each
virus at an MOI of 5 was adsorbed to approximately 6x105 Vero cells at 4°C for 1 h.
Thereafter, pre-warmed media was added and virus was allowed to penetrate for 2 hours
at 37°C. Any remaining extracellular virus was inactivated by low-pH treatment (0.1M
133
glycine, pH 3.0). Cells and supernatants were harvested immediately thereafter (0 h) or
after 4, 9, 16, 20 or 24 h incubations at 37° C. Virus titers were determined by end point
titration of virus stocks on a complementing cell line.
Generation of Recombinant UL20-null and gK-null Viruses Specifying Mutant gB
In order to generate recombinant viruses specifying mutations in gB, PCRamplification of the UL27 region was performed from purified viral DNA of YEbac102
(wt gB), tsb5 (gBsyn3), or amb1511 (gBamb1511). Each PCR product was used for
homologous recombination with both the Yebac∆gB∆UL20 and YEbac∆gB∆gK viruses.
Virus plaques that formed on Fd20-1 (UL20 complementing) or FcgK-1 (gK
complementing) cells were picked and sequentially plaque-purified at least five times.
Replacement of the gB-null resident kanamycin cassette and the concomitant insertion of
the specific mutant gB gene were confirmed by DNA sequencing.
Generation of UL19 and UL20 Double-Null Viruses
Viruses that contained a CMV-EGFP gene cassette in place of a UL19/ UL20
gene deletion were generated by homologous recombination with plasmid p∆1920-EGFP
and the parental viruses gKDIV5, gKsyn20DIV5, or tsB5 (gBsyn3). G5 cells were
transfected with p∆1920-EGFP and 6h post transfection (hpt) were infected at an MOI of
0.1 with either gKDIV5, gKsyn20DIV5, or tsB5 (gBsyn3) to generate UL19 and UL20
double-null viruses that were designated ∆1920, ∆1920-syn20 and ∆1920-syn3,
respectively. Viruses were subsequently plaque isolated on G5 cells by fluorescence
microscopy and sequentially plaque purified at least five times. Insertion of the CMVEGFP gene cassette and the concomitant deletion of the UL19 and UL20 genes were
confirmed by diagnostic PCR and DNA sequencing.
134
Electron Microscopy
Cell monolayers were infected with the indicated virus at an MOI of 3. All cells
were prepared for TEM examination 24 hpi. Infected cells were fixed in a mixture of 2%
paraformaldehyde and 1.5% glutaraldehyde in 0.1M NaCaC buffer, pH 7.3. Following
osmication (1% OsO4) and dehydration in an ethanol series, the samples were embedded
in Epon-Araldyte resin and polymerized at 70ºC. Thin sections were made on an MTXL
Ultratone (RMC Products), stained with 5% uranyl acetate and CNA lead, and observed
with a Zeiss 10 transmission electron microscope as described previously (Foster et al.,
2004a; Melancon, Foster, and Kousoulas, 2004).
RESULTS
Construction of the HSV-1 BACs pYEbac∆gB, pYEbac∆UL20, pYEbac∆gK,
pYEbac∆gB∆UL20, and pYEbac∆gB∆gK
The complete HSV-1(F) genome was recently cloned into a bacterial artificial
chromosome (pYEbac102) enabling the genetic manipulation of the HSV-1 genome in E.
coli. (Tanaka et al., 2003). The BAC-based GET homologous recombination system was
utilized to construct deletions within the gB, UL20 and gK genes in E. coli. Deletion of
each gene was accomplished by targeting the gB, UL20 or gK open reading frames
(ORFs) using specific primers as detailed in the Materials and Methods section. For
construction of the ∆UL20-GFPZeo mutation, primers A and B were used for
homologous recombination and positioned to remove a 353 bp region of the UL20 gene
upon insertion of a GFP-Zeocin gene cassette (Table 3.1; Fig. 3.1 B, C), extending from
the UL20 ATG to a HpaI site inside the UL20 ORF. Inactivation of the gB gene was
achieved by insertion of a kanamycin gene cassette within gB (∆gB-Kan mutation).
Specifically, primers C and D were designed to remove a 970 bp fragment starting with
135
gB ORF nucleotide 1655 upon homologous recombination with pYEbac102 (Table 3.1;
Fig 3.1 B, C). This gB deletion does not disrupt the UL27.5 ORF overlapping the 5’ end
of gB (Chang et al., 1998) and is similar to that of the K∆T gB-null virus (Cai et al.,
1987), with the exception that the ∆gB-Kan mutation was constructed to include a stop
codon at the 5’ end of the kanamycin cassette ensuring that no aberrant proteins could be
made via a translational read-through of the kanamycin gene cassette. For construction
of the ∆gK-GFPZeo mutation, primers E and F were positioned to delete almost the
entire gK gene (941 bp) without affecting the proximal UL52 gene, which partially
overlaps with the gK ORF (Table 3.1; Fig 3.1 B, C). Each pYEbac102 mutant construct
contains one or more of the insertion-deletion mutations mentioned above: pYEbac∆gB
contains the ∆gB-Kan mutation; pYEbac∆UL20 contains the ∆UL20-GFPZeo mutation;
pYEbac∆gK contains the ∆gK-GFPZeo mutation; pYEbac∆gB∆UL20 contains both the
∆gB-Kan and the ∆UL20-GFPZeo mutations; and pYEbac∆gB∆gK contains both the
∆gB-Kan and the ∆gK-GFPZeo mutations.
The YEbac102 viral constructs were tested for the presence of the engineered
insertion-deletion mutation via diagnostic PCR. Primers a and b (Table 3.1; Fig 3.1A),
located outside of the UL20 gene, amplified the predicted 909 bp diagnostic UL20 DNA
fragments from BACs that contain wt UL20, while a predicted 2,469 bp product was
produced from BACs that contain the ∆UL20-GFPZeo mutation (Fig. 3.2A, panel i).
Primers c and d (Table 3.1; Fig 3.1A) located within gB but outside the deleted region
amplified the predicted 2,127 bp diagnostic gB DNA fragment from BACs that contain
wt gB, while a predicted 2,261 bp product was produced from BACs that contain the
136
Table 3.1: Synthetic Oligonucleotide Primers
Primer
Primer sequence*
Designation
Name
A
3' UL20-GFPZeo
5'-CACGGACATCCCCCAAAACACGGGCGCCGACAACGGCA
GACGATCCCTCTTGATGTTagacatgataagatacattgatgagtttgg-3'
B
5' UL20-GFPZeo
5'-CTGACGTAAGCGACCCTTTGCGGTTTCGGTCTCCCCACCT
CCACCGCACACCCCgtccgttacataacttacggtaaatgg-3'
C
3' gB-Kan
5'-GTTGGGAACTTGGGTGTAGTTGGTGTTGCGGCGCTTGCGC
ATGACCATGTCcggttgatgagagctttgttgtaggtggac-3'
D
5' gB-Kan
5'-CTGACCCTGTGGAACGAGGCCCGCAAGCTGAACCCCAAC
GCCATCGCCTCGTGAccacgttgtgtctcaaaatctctgatgtta-3'
E
5' gK-GFPZeo
5'-AGCACCTCTCAACCGTCGTCTTGATAACGGCGTACGGCCT
CGTGCTCGTGTGGTGAgtccgttacataacttacggtaaatgg-3'
F
3' gK-GFPZeo
5'-CAGTTTCCAATTTGCATATGCCGTTACGGTTTCCGCCGGCC
TGGATGTGACGagacatgataagatacattgatgagtttgg-3'
a
UL20-Rev
5'-GAAGGCTGCGGCCTCGTTCCAG-3'
b
UL20-For
5'-ACGGCCTTATCAAAACACTCGCCTC-3'
c
gB-Rev
5'-TCACAGGTCGTCCTCGTCGGC-3'
d
gB-For
5'-GTGATCGACAAGATCAACGCCAAGG-3'
e
gK-For
5'-GGCACGCCATGGTCGCCGTCCGTTCCCTGC-3'
f
gK-Rev
5'-ATCACATACCCCGTTCCGCTTCCG-3'
Purpose and product size
UL20 GET recombination (2025 bp)
gB GET recombination
(1209 bp)
gK GET recombination
(2022 bp)
Diagnostic PCR for wt UL20 (909 bp)
or ∆UL20-GFPZeo (2469 bp)
Diagnostic PCR for wt gB (2127 bp)
or ∆gB-Kan (2261 bp)
Diagnostic PCR for wt gK (1265 bp)
or ∆gK-GFPZeo (2238 bp)
* For GET recombination primers, the region homologous to HSV-1 DNA is upper case, and the region which binds to the marker gene is lower case
137
UL
US
IRL IRS
TR
TR
A
0.0
B
0.1
UL19 (VP5)
0.2
UL20
0.3
UL20.5
a
GFP-Zeocin
C
A
0.4
b
0.5
0.6
UL27 (gB)
0.7
0.8
UL52
c
d
CMV
∆UL20-GFPZeo mutation
C
UL53 (gK)
CMV
D
∆gB-Kan mutation
1.0
UL54
e
Kanamycin
B
0.9
E
f
GFP-Zeocin
F
∆gK-GFPZeo mutation
Figure 3.1: Schematic of the strategy for the construction of pYEbac102 mutant
BACs. (A) The top line represents the prototypic arrangement of the HSV-1 genome with
the unique long (UL) and unique short (US) regions flanked by the terminal repeat (TR)
and internal repeat (IR) regions. (B) Shown below are the expanded genomic regions of
the UL20, UL27, and UL53 ORFs as well as the approximate locations of the sites where
insertion of the marker genes was targeted and the primers used in diagnostic PCR to
confirm the presence of each mutation. (C) PCR fragments containing the kanamycin or
GFP-Zeocin gene cassette flanked by ~50 bp of viral sequences on both sides were used
for targeted GET recombination in E.coli to construct pYEbac102 mutant BACs with
insertion-deletion mutations in the UL20, UL27, and/or UL53 ORFs. The approximate
location of the primers used in amplification of each PCR fragment are also shown.
138
∆gB-Kan mutation (Fig. 3.2A, panel iii). Primers e and f (Table 3.1; Fig 3.1A) located
outside of the gK deletion region amplified a 1,265 bp diagnostic gK DNA fragment
from BACs that contain wt UL20, while a predicted 2,238 bp product was produced from
BACs that contain the ∆gK-GFPZeo mutation (Fig. 3.2A, panel ii).
Further confirmation of the genetic content of the pYEbac102 mutant constructs
was obtained via restriction endonuclease fragment analysis (Fig. 3.2B) and Southernblotting (Fig. 3.2C, D). Restriction enzyme analysis of pYebac102, pYEbac∆gB,
pYEbac∆UL20, pYEbac∆gK, pYEbac∆gB∆UL20 and pYEbac∆gB∆gK DNA with KpnI
produced a similar genomic DNA restriction pattern with a few exceptions caused by the
specified alterations of gB, UL20 and gK containing KpnI DNA fragments. First, the
4,701 bp band containing the UL27 (gB) ORF increased in size by 134 bp to 4,835 bp for
pYEbac∆gB, pYEbac∆gB∆UL20 and pYEbac∆gB∆gK as a result of the ∆gB-Kan
mutation. This 4,835 bp KpnI DNA fragment (demarcated by a black arrow) was visible
in the lanes labeled with the above mentioned constructs appearing between the predicted
4,680 bp and 4959 bp KpnI DNA fragments (Fig. 3.2B). In addition, Southern blot
analysis revealed that a kanamycin gene cassette probe hybridized to a single 4,835 bp
KpnI DNA fragment contained only in pYEbac∆gB, pYEbac∆gB∆UL20 and
pYEbac∆gB∆gK restriction digests (indicated by a black arrow) (Fig. 3.2C), indicating
that the ∆gB-Kan mutation had been successfully transferred into a single genomic site.
Second, the wild-type 9,432 bp KpnI DNA fragment containing the UL20 ORF increased
in size by 1,560 bp to 10,992 bp for pYEbac∆UL20 and pYEbac∆gB∆UL20. It is
apparent that the 9,432 bp band is missing in pYEbac∆UL20 and pYEbac∆gB∆UL20, as
indicated by a gray arrow (Fig. 3.2B). However, the 10,992 bp KpnI DNA fragment is
139
Figure 3.2: Genomic analysis of pYEbac102 mutants. (A) Diagnostic PCR to confirm
the insertion of the relevant gene cassettes in place of the deleted genomic regions in
UL20 (A, panel i) UL27 (gB) (A, panel ii), and UL53 (gK) (A, panel iii). (B) KpnI
restriction fragment analysis of pYEbac102 mutant BACs in comparison to the wild-type
pYEbac102. The black arrow demarcates the position of the KpnI DNA fragment altered
by the ∆gB-Kan mutation, while the gray arrow points to the missing 9,432 bp fragment
resulting from the ∆UL20-GFPZeo mutation. (C, D) Southern-blot analysis of
pYEbac102 mutant BACs. The KpnI restricted BACs from Figure 2B were hybridized
with either a kanamycin (C) or GFP-Zeocin (D) biotinylated probe. The biotinylated
kanamycin probe hybridized to an estimated 4,835 bp DNA fragment in pYEbac∆gB,
pYEbac∆gB∆gUL20 and pYEbac∆gB∆gK DNA, which was absent in the wild-type
pYEbac102 DNA (indicated by a black arrow in panel C). The biotinylated GFP-Zeocin
probe identified a predicted 10,992 bp DNA fragment (gray arrow) in pYEbac∆UL20 and
pYEbac∆gB∆gUL20 DNA corresponding to the ∆UL20-GFPZeo mutation, and a
predicted 8,956 bp fragment (white arrow) in pYEbac∆gK and pYEbac∆gB∆gK DNA
corresponding to the ∆gK-GFPZeo mutation (D).
140
141
not easily identifiable due to the presence of a KpnI genomic fragments with predicted
sizes of 10,837 and 11,024 bp. The 10,992 bp DNA fragment contains the ∆UL20GFPZeo mutation, which was confirmed by probing the corresponding blot with a GFPZeocin probe (Fig. 3.2D, gray arrow). The GFP-Zeocin probe hybridized to unique DNA
fragments of the predicted sizes for both pYEbac∆UL20 and pYEbac∆gB∆UL20.
Finally, the ∆gK-GFPZeo mutation is predicted to increase in size of the wild-type 7,983
bp DNA fragment containing the UL53 (gK) ORF by 973 bp to 8,956 bp. These DNA
fragments were not readily discernible in the ethidium bromide stained restriction profile
(Fig. 3.2A). However, the GFP-Zeocin probe hybridized to the 8,956 bp DNA fragment
(white arrow) in pYEbac∆gK and pYEbac∆gB∆gK confirming the creation of the ∆gKGFPZeo mutation (Fig. 3.2C). Again, the presence of a single DNA fragment detected by
the GFP-Zeocin probe indicates the absence of any spurious insertions.
Generation of Infectious Virus from pYEbac102-Based Constructs
To generate virus stocks from the mutant pYEbac102 constructs, transient
transfection of individual BAC DNAs was performed into specific cell lines transformed
with either the gB, UL20 or gK gene and capable of complementing the resultant viruses.
Specifically, pYEbac102 was transfected into Vero cells, pYEbac∆gB into gB
complementing D6 cells (Cai et al., 1987), pYEbac∆UL20 into UL20 complementing
Fd20-1 cells (Melancon, Foster, and Kousoulas, 2004), and pYEbac∆gK into FcgK-1
cells. The FcgK-1 cell line constitutively expresses gK gene under the CMV IE promoter
and was constructed to efficiently complement the ∆gK-GFPZeo mutation and minimize
the chance for generation of wild-type viruses via recombination with the endogenous gK
gene (see Materials and Methods). For all transfection experiments, virus plaques
142
became visible 72 hours post transfection and virus stocks were collected at appropriate
points exhibiting maximum cytopathic effects.
For complementation of the double-null viruses produced by transfection of
pYEbac∆gB∆UL20 and pYEbac∆gB∆gK, the replication-deficient adenovirus Ad5-gB
was constructed, which constitutively expresses gB under the CMV-IE promoter (see
Materials and Methods). A series of experiments were performed to assess the optimum
MOI of this virus that could maximize complementation of gB-null viruses. These
experiments showed that Ad5-gB at an MOI of 10 efficiently complemented gB-deficient
viruses (not shown). To complement double-null viruses having the gB gene deletion, the
UL20 or gK complementing cell lines were infected with Ad5-gB one hour prior to
transfection with pYEbac∆gB∆UL20 or pYEbac∆gB∆gK. The same strategy was used
to amplify virus stocks of the double-null viruses.
Plaque Morphology and Growth Kinetics of HSV-1 YEbac102 Mutants
HSV-1 viruses containing deletions in UL20, gB, and gK have been described
previously (Baines et al., 1991; Cai et al., 1987; Foster et al., 2004a; Hutchinson et al.,
1993; Jayachandra, Baghian, and Kousoulas, 1997). These viruses were produced by
homologous recombination of plasmid constructs with viral DNA in cell culture
experiments. Typically, isolation of these viruses required multiple plaque purifications
to ensure that the mutant virus stocks did not contain contaminating wild-type virus. In
contrast, the YEbac102 UL20, gB and gK null viruses were constructed in E. coli using
the GET recombination system (Narayanan et al., 1999; Orford et al., 2000). In this case,
production of mutant viruses did not require any plaque purification, since the progeny
viruses did not contain any contaminating wild-type genomes.
143
Figure 3.3: Plaque phenotypes of gB, gK and UL20-null viruses. Plaque phenotypes
of YEbac102, YEbac∆UL20, YEbac∆gB and YEbac∆gK were observed on Vero (A1,
B1, C1, D1), Fd20-1 (A2, B2, C2, D2), D6 (A3, B3, C3, D3) and FcgK-1 (A4, B4, C4,
D4) cells. Confluent cell monolayers were infected with the wild-type YEbac102 (A1,
A2, A3, A4), YEbac∆UL20 (B1, B2, B3, B4), YEbac∆gB (C1, C2, C3, C4) or
YEbac∆gK (D1, D2, D3, D4) at an MOI of 0.001, and viral plaques were visualized at 48
hpi by immunohistochemistry.
144
To assess the effect of the deletion of the various genes on cell-to-cell spread, the
plaque morphologies of the YEbac102, YEbac∆gB, YEbac∆UL20 and YEbac∆gK
viruses were examined in Vero, Fd20-1, D6 and FcgK-1 cells at 48 hpi (Fig 3.3). The
wild-type YEbac102 produced large plaques of a similar size on all infected cells (Fig
3.3: A). Infection of non-complementing cells with either YEbac∆UL20 or YEbac∆gK
resulted in small plaques containing on average 3 to 5 cells (Fig 3: B1, B3, B4 and D1,
D3, D3, respectively), while complementation of the UL20 and gK gene deletions
resulted in large plaques comparable in size to the YEbac102 plaques (Fig 3.3: B2, D4,
respectively). Similarly, infection of non-complementing cells with the YEbac∆gB virus
produced only single infected cells (Fig. 3.3: C1, C2, C4), indicating that the virus was
unable to spread beyond the primary infected cell, while infection of complementing D6
cells resulted in large plaques (Fig. 3.3: C3).
The YEbac∆gB∆UL20 and YEbac∆gB∆gK double-null viruses formed large
plaques when plated onto Ad5-gB infected Fd20-1 or FcgK-1 cells, respectively (Fig.
3.4: A5, B6), single infected cells on Vero cells or cells able to complement for only gK
or UL20 (Fig. 3.4: A1, A2, A4, B1, B2, B4), or very small plaques in cells able to
complement gB (Fig. 3.4: A3, A6, B3, B5).
Rescue of the individual mutations from each BAC-derived virus was performed
by homologous recombination with wild-type DNA fragments spanning each gene.
Double-null mutations were rescued sequentially with the gB-null mutation rescued first
followed by rescue of either the gK-null or UL20-null mutations. Importantly, all rescued
viruses formed large plaques similar to the wild type YEbac102 virus when plated on
non-complementing Vero cells, indicating that there were no spurious mutations in the
145
Figure 3.4: Plaque phenotypes of double-null viruses and rescued viruses. Plaque
phenotypes of YEbac∆gB∆gUL20 and YEbac∆gB∆gK on Vero (A1, B1), Fd20-1 (A2,
B2), D6 (A3, B3), and FcgK-1 (A4, B4) cells, or Ad5-gB infected Fd20-1 (A5, B5) and
FcgK-1 (A6, B6) cells are shown. Confluent cell monolayers were infected with
YEbac∆gB∆gUL20 (A1, A2, A3, A4, A5, A6) or YEbac∆gB∆gK (B1, B2, B3, B4, B5,
B6) at an MOI of 0.001. The plaque phenotypes of the rescued gB, gK and UL20-null
(D, E, F) and double-null (G, H) viruses in comparison to the wild-type YEbac102 (C)
are shown on Vero cells. Viral plaques were visualized at 48 hpi by
immunohistochemistry.
146
Figure 3.5: Viral replication kinetics. Comparison of virus replication characteristics
of YEbac102 (●),YEbac∆UL20 (■),YEbac∆gK (▲),YEbac∆gB (○),YEbac∆gB∆UL20
(□), and YEbac∆gB∆gK (∆) on Vero cells. One step kinetics of infectious virus
production were calculated after infection at an MOI of 5 followed by incubation at 37°C.
147
parental mutant BAC viruses affecting virus replication and egress (Fig. 3.4: C, D, E, F,
G, H).
To examine the effect of the various mutations on virus replication, Vero cells
were infected at an MOI of 5 with either the wild-type or each mutant virus. Virus stocks
were prepared at 0, 4, 9, 16, 20, or 24 hpi and titrated in triplicate onto complementing
cells (Fig. 3.5). The kinetics of both YEbac∆UL20 and YEbac∆gK viral replication was
substantially slower than that of the YEbac102 virus. Maximum titers for the
YEbac∆UL20 and YEbac∆gK mutants viruses were 105 pfu/ml in comparison to 108
pfu/ml for the YEbac102 virus at 24 hpi (Fig. 3.5). In contrast, the YEbac∆gB,
YEbac∆gB∆UL20 and YEbac∆gB∆gK viruses failed to produce infectious virus in Vero
cells (Fig. 3.5).
Ultrastructural Characterization of the YEbac∆gB∆UL20 and YEbac∆gB∆gK
Double-Null Mutant Viruses
Previous reports indicated that gB was not essential for virion egress (Cai et al.,
1987; Cai, Gu, and Person, 1988), while both gK and UL20p were found to be essential
for cytoplasmic virion morphogenesis and egress (Foster et al., 2004a; Jayachandra,
Baghian, and Kousoulas, 1997). To ascertain whether gB functioned redundantly with
either gK or UL20p in virion de-envelopment from perinuclear spaces, the ultrastructural
phenotypes of each YEbac102-based mutant virus carrying UL20, gB, or gK deletions
relative to the wild-type YEbac102 virus were determined by transmission electron
microscopy at 24 hpi. As expected, the YEbac102 and YEbac∆gB viruses exhibited no
apparent defects in virion egress exemplified by the presence of fully enveloped virions
extracellularly (Fig 3.6 A, B), as well as the presence of fully enveloped virions
intracellularly (not shown). Unlike the wild-type virus, ultrastructural visualization
148
Figure 3.6: Ultrastructural morphology of gB, gK and UL20-null viruses. Electron
micrographs of Vero cells infected with YEbac102 (A), YEbac∆gB (B), YEbac∆UL20
(C) or YEbac∆gK (D). Confluent cell monolayers were infected at an MOI of 5,
incubated at 37°C for 24 hours, and prepared for transmission electron microscopy.
Nuclear (n), cytoplasmic (c) and extracellular (e) spaces are marked.
149
Figure 3.7: Ultrastructural morphology of gB/gK and gB/UL20 double-null viruses.
Electron micrographs of Vero cells infected with YEbac∆gB∆gUL20 (A1, A2, A3, A4,
A5) or YEbac∆gB∆gK (B1, B2, B3, B4). Confluent cell monolayers were infected at an
MOI of 5, incubated at 37°C for 24 hours, and prepared for transmission electron
microscopy. Panels A4 and B3 show partially-enveloped capsids. Panels A5 and B4 show
a tegument-like accumulation on membranes that are folded irregularly. Nuclear (n),
cytoplasmic (c) and extracellular (e) spaces are marked.
150
151
of YEbac∆UL20 and YEbac∆gK infected Vero cells revealed a cytoplasmic defect in
virion egress, characterized by the presence of unenveloped capsids in the cytoplasm as
well as aberrantly enveloped virions (Fig. 3.6 C, D). These results are consistent with
previous findings (Foster et al., 2004a; Jayachandra, Baghian, and Kousoulas, 1997). In
general, ultrastructural examination of Vero cells infected with the YEbac∆gB∆gK (Fig.
3.7A) and YEbac∆gB∆UL20 (Fig. 3.7B) double-null viruses revealed similar virion
morphogenetic defects to those of the gK and UL20-null viruses exhibiting high numbers
of unenveloped capsids in the cytoplasm. In addition, the observed ultrastructural
phenotypes revealed specific defects in cytoplasmic envelopment: i) multiple capsids
appeared proximal to membranes apparently unable to complete the budding process into
cytoplasmic vesicles (Fig. 3.7: A4, B3); ii) cytoplasmic vesicular membranes appeared to
contain electron dense material consistent with the accumulation of tegument proteins in
the presence or absence of proximal capsids (Fig. 3.7: A5, B4).
Generation and Characterization of Recombinant gK-null and UL20-null viruses
Containing the gBsyn3 or gBamb1511 Mutation
It was previously determined that gB was required for the expression of the syn20
(gK) syncytial phenotype (Cai, Gu, and Person, 1988). To determine whether gK was
required for virus-induced cell fusion due to syncytial mutations in gB, YEbac102-based
recombinant viruses were constructed carrying either the gBsyn3 or the gBamb1511
syncytial mutations in the absence of the gK gene. These viruses were constructed by
rescuing the YEbac∆gB∆gK virus with either wild-type gB, gBsyn3 or gBamb1511 (see
Materials and Methods). The same strategy was used to isolate recombinant viruses
carrying the gBsyn3 or gBamb1511 mutations in the absence of the UL20 gene by
rescuing the gB deletion in the YEbac∆gB∆UL20 virus.
152
Figure 3.8: Plaque morphology of gK and UL20-null viruses carrying syncytial
mutations in gB. Plaque phenotypes of gK-null (A, B, C, D, E, F) and UL20-null (G, H,
I, J, K, L) recombinant viruses containing specific gB mutations on Vero (A, C, E, G, I,
K) and FcgK-1 (B, D, F) or Fd20-1 (H, J, L) cells. Confluent cell monolayers were
infected with the gK-null viruses containing: wild-type gB (A, B), gBsyn3 (C, D), or
gBamb1511 (E, F); or UL20-null viruses containing wild-type gB (G, H), gBsyn3 (I, J),
or gBamb1511 (K, L) at an MOI of 0.001, and viral plaques were visualized at 48 hpi by
immunohistochemistry. Insets are shown at 2.4x magnification to facilitate visualization
of the non-syncytial phenotype of the small plaques.
153
To assess the effect of each gB mutation on virus-induced cell fusion, Vero cells,
as well as either gK or UL20 complementing cells, were infected with each mutant virus
at an MOI of 0.001. Virus plaques were visualized at 48 hpi by immunohistochemistry
as described previously (Foster et al., 2004a; Melancon, Foster, and Kousoulas, 2004).
Viruses specifying the wild type, syn3, or amb1511 gB in the gK-null background
formed small, non-syncytial plaques on Vero cells (Fig. 3.8: A, C, E). As expected, the
virus containing the wild-type gB formed large, non-syncytial plaques on FcgK-1 cells
(Fig. 3.8: B), while gK-null viruses specifying either the gBsyn3 or gBamb1511 mutation
formed large, syncytial plaques on FcgK-1 cells (Fig 3.8: D, F). Similarly, UL20-null
viruses specifying the wild-type, syn3, or amb1511 gB formed small, non-syncytial
plaques on Vero cells (Fig. 3.8: G, I, K). The UL20-null virus containing the wild-type
gB formed large, non-syncytial plaques on Fd20-1 cells (Fig. 3.8: H), and the UL20-null
viruses specifying either the gBsyn3 or gBamb1511 mutation formed large, syncytial
plaques on Fd20-1 cells (Fig. 3.8: J, L).
Virion Assembly and Egress Is Not Required for Either gB or gK-Mediated VirusInduced Cell-to-Cell Fusion
To investigate the potential requirement for virion egress in virus-induced cell
fusion, recombinant viruses were constructed carrying a UL19 and UL20 deletion in an
otherwise wild-type, syn20, or tsB5 genetic background. UL19 encodes the major capsid
protein (VP5), and capsid formation can not occur in its absence (Desai et al., 1993).
Briefly, a CMV-EGFP gene cassette was inserted into the viral genome effectively
deleting a sizeable portion of the UL19 ORF and all of the UL20 ORF. Recombinant
viruses were isolated via fluorescence microscopy and plaque purified on G5 cells, which
complement the UL19/UL20-null defect (Desai et al., 1993), and characterized as
154
Figure 3.9: Plaque phenotypes of ∆1920, ∆1920-syn20 and ∆1920-syn3 viruses on
Vero (A, C, E) and FcgK-1 (B, D, F) cells. Confluent cell monolayers were infected
with the ∆1920 (A, B), ∆1920-syn20 (C, D) or ∆1920-syn3 (E, F) at an MOI of 0.001,
and viral plaques were visualized at 48 hpi by immunohistochemistry.
155
described in Materials and Methods. CV-1 or Fd20-1 cells were infected with ∆1920,
∆1920-syn20 and ∆1920-syn3 viruses at an MOI of 0.005. As expected, these infections
did not cause virus-induced cell fusion in non-complementing CV-1 cells (Fig. 3.9: A, C,
E). However, the ∆1920-syn20 and ∆1920-syn3 viruses formed syncytial plaques
containing a range of 3 to 100 nuclei per syncytium in Fd20-1 cells, which complement
only UL20 (Fig. 3.9: D, F), while the ∆1920 virus failed to cause virus-induced cell
fusion (Fig. 3.9: B).
DISCUSSION
HSV-1 BAC Generated Recombinant Viruses
Investigations of the structure and function of individual viral proteins has been
greatly facilitated through the use of recombinant viruses specifying mutated proteins.
Typically, recombinant viruses have been produced by homologous recombination
between a mutated DNA fragment and viral genomes. In most instances, the resultant
recombinant viruses have to be extensively purified due to the presence of a large amount
of contaminating wild-type viruses. Recently, the HSV-1 (F) genome was cloned into a
bacterial artificial chromosome (BAC) by inserting the BAC into the intergenic region
between UL3 and UL4 without deletion of any viral genes (Tanaka et al., 2003), unlike
previously constructed HSV-1 BACs that contained deletions of one or more viral genes
(Horsburgh et al., 1999; Saeki et al., 1998; Stavropoulos and Strathdee, 1998).
Transfection of the resultant pYEbac102 plasmid into cells resulted in production of the
YK304 infectious virus, which was indistinguishable from wild-type HSV-1 in vitro and
in vivo, exhibiting wild-type virulence in mice after intracerebral inoculation (Tanaka et
al., 2003).
156
Taking advantage of the HSV-1 pYEbac102 system, we generated recombinant
viruses lacking either the gK, gB or UL20 genes as well as viruses having deletions in
both gK and gB, or gB and UL20. As expected, the gB/gK and gB/UL20 double-null
viruses failed to produce infectious virus or spread beyond the primary infected Vero
cells. Generally, the gK, gB or UL20-null viruses exhibited similar defects in virus
replication and plaque morphologies to corresponding viruses constructed in the KOS
genetic background. However, it was noted that the HSV-1(F) gK and UL20-null viruses
exhibited up to 10-fold greater defect in virus replication, while virus plaques were
typically smaller than those produced by the KOS mutant viruses. These results may be
attributed to genetic differences between the F and KOS virus strains, although no
apparent differences were noted between the wild-type viruses with regard to their
replication kinetics and plaque morphology. Alternatively, it is possible that the
requirement for extensive passage of mutant viruses in the KOS background during
plaque purification may have resulted in improved adaptation of these viruses for growth
in Vero cells. In this regard, generation of mutant viruses via the pYEbac102 system
provides a unique advantage in assessment of the structure and function of individual
viral proteins without the potential complications arising from extensive cell culture
passage of virus stocks.
gB, UL20p and gK are not Required for Virion De-Envelopment
HSV-1 is thought to acquire an initial envelope when capsids formed in the
nucleus of cells bud into the perinuclear space. Subsequently, a putative fusion event
between viral envelopes and the outer nuclear membranes results in capsid release into
the cytoplasm. Final envelopment is thought to occur by budding into cytoplasmic
157
vesicles derived from the TGN or endosomes that facilitate egress of enveloped virions to
extracellular spaces (Fig. 3.10). Currently, it is not known whether viral glycoproteins
involved in virus entry, or virus-induced cell fusion also function in this de-envelopment
step. Results obtained in this study clearly show that gK, UL20 or gB are not essential for
virion de-envelopment. In the case of gB, previous studies suggested that lack of gB did
not adversely influence egress of virion particles to supernates of infected cells, although
these virions lacked gB and were rendered non-infectious (Cai et al., 1987; Cai, Gu, and
Person, 1988). Similar results have been reported for gH, gL and gD, which are thought
to be required for virus-induced and virus-free (transient expression) membrane fusion
(Forrester et al., 1992; Ligas and Johnson, 1988; Roop, Hutchinson, and Johnson, 1993).
Ultrastructural examination of the gB-null virus confirmed initial findings and clearly
showed that lack of gB did not adversely affect the production of enveloped virions in
extracellular spaces. Investigations with recombinant viruses specifying single gene
deletions in either gB, gK or UL20p did not exclude the possibility that gB and gK may
function in a redundant manner to facilitate virion de-envelopment. This possibility can
be discounted on the basis that the gB/gK double null virus efficiently produced
unenveloped capsids into the cytoplasm of infected cells. Similar results were obtained
for the gB/UL20 double null virus. Furthermore, it is improbable that UL20p or gK alone
could mediate virion de-envelopment due to the fact that these two proteins act in a
cooperative manner for intracellular transport and virus-induced cell fusion. Construction
of a triple-null virus lacking gB, gK and UL20p could resolve this possibility.
Alternatively, other viral glycoproteins such as gH/gL may function redundantly with gK,
UL20p or gB in the de-envelopment process. Virions found in perinuclear spaces are
158
Figure 3.10: Schematic of HSV-1 virion morphogenesis and egress. (I) Mature
capsids budding through the inner nuclear membrane into the perinuclear space; (II) Deenvelopment of perinuclear virions at the outer nuclear membrane; (III) Re-envelopment
of cytoplasmic capsids by budding into cytoplasmic vesicles; (IV) Final egress to the
extracellular space.
159
known to contain the UL34 and UL31 tegument proteins, which are absent from
extracellular enveloped virions (Reynolds et al., 2002). Thus, it is possible that the deenvelopment process involves different tegument-membrane protein interactions from
those occurring during virus entry. In this regard, virion de-envelopment from perinuclear
spaces seems to bear little similarity to either virus entry or virus-induced cell fusion
suggesting that this membrane fusion event may be regulated by a fundamentally
different mechanism.
Role of gK on gB-Mediated Cell-to-Cell Fusion
Previous reports showed that gK syncytial mutations could not cause virusinduced cell fusion in the absence of gB, indicating that gB served essential functions for
cell-to-cell fusion (Cai, Gu, and Person, 1988). Both gBsyn3 and gBamb1511 mutations
cause extensive virus-induced cell fusion (Baghian et al., 1993; Bzik et al., 1984).
However, unlike the gBsyn3 single amino acid mutation, which does not enhance gBmediated fusion in the transient coexpression system, the gBamb1511 truncation causes
greatly enhanced fusion in the transient expression system (Foster, Melancon, and
Kousoulas, 2001). These findings suggest that the gBamb1511 truncation may disengage
gB from other viral or cellular functions responsible for down-regulation of the fusogenic
potential of gB. However, recombinant viruses containing the gB syncytial mutations
gBsyn3 or gBamb1511, but lacking the gK or the UL20 gene, failed to cause virusinduced cell fusion indicating that expression of both gK and UL20p is required for gBmediated membrane fusion. In addition, deletion of the carboxyl terminal 28 amino acids
of gB specified by the amb1511 mutation does not abrogate the requirement for
expression of both gK and UL20p in gB-mediated virus-induced cell fusion, suggesting
160
that gB amino acid sequences other than the carboxyl terminal 28 amino acids are
required for gK or UL20p regulatory interactions with gB. Recently, gK expression was
reported to inhibit membrane fusion caused by coexpression of gB, gD, gH and gL
(Avitabile, Lombardi, and Campadelli-Fiume, 2003). These findings seem to contradict
results obtained herein with the recombinant viruses lacking gK. However, a similar
result was reported for gM, which inhibited membrane fusion in the transient
coexpression system (Klupp, Nixdorf, and Mettenleiter, 2000; Koyano et al., 2003), but
was required for virus-induced cell fusion caused by syncytial mutations in gB (DavisPoynter et al., 1994). These results suggest that gM and gK may regulate virus-induced
cell fusion both positively as well as negatively depending on the presence of different
sets of viral proteins.
Virion Egress is not Required in Virus-Induced Cell Fusion
Virus-induced cell fusion starts at approximately 5 hpi at 37° C at which time
infectious virions are produced and egress out of infected cells. Therefore, it is possible
that egressing virions may contribute directly or indirectly to the virus-induced
membrane fusion phenomena mediated by cell-surface expressed gB and gK carrying
syncytial mutations. It has been shown that cell surface expression of gB is not affected
in a gK-null or UL20-null virus (Foster et al., 2004a; Jayachandra, Baghian, and
Kousoulas, 1997). Therefore, the requirement for the presence of UL20p and gK for gBmediated virus-induced cell fusion may be due to defects in virion egress. Earlier reports
indicated that production of virion particles may not be necessary for virus-induced cell
fusion, inasmuch as, syncytial mutants caused fusion in the presence of the replication
inhibitor phosphonoacetic acid (Kousoulas, Person, and Holland, 1978). These early
161
findings were confirmed herein by the phenotype of the UL19/UL20 double-null viruses
containing either the gBsyn3 or gBamb1511 mutation. These viruses are unable to form
capsids because of the deletion in the UL19 gene, coding for the major capsid protein
VP5. Both viruses caused virus-induced cell fusion in the UL20 complementing cell line
indicating that virion formation and egress are not necessary for virus-induced cell
fusion.
The rapid generation of recombinant viruses using the YEbac102 system greatly
facilitates the characterization of the function of individual viral proteins, while helping
to discern potentially complex interactions among multiple proteins through the
introduction of multiple mutations into an identical genetic background. In this report, we
have demonstrated the utility of this system in investigating the role of gB, gK and
UL20p in virus induced cell fusion and morphogenesis. The existence of an intimate
relationship between gK/UL20p and gB in virus-induced cell fusion suggests that both
gK and UL20p act to regulate the fusogenic properties of gB. Future studies will be
targeted to investigate direct physical interactions among gB, gK and UL20p and their
role in the regulation of membrane fusion phenomena in the HSV-1 life cycle.
REFERENCES
Avitabile, E., Lombardi, G., and Campadelli-Fiume, G. (2003). Herpes simplex virus
glycoprotein K, but not its syncytial allele, inhibits cell-cell fusion mediated by
the four fusogenic glycoproteins, gD, gB, gH, and gL. J Virol 77(12), 6836-44.
Baghian, A., Huang, L., Newman, S., Jayachandra, S., and Kousoulas, K. G. (1993).
Truncation of the carboxy-terminal 28 amino acids of glycoprotein B specified by
herpes simplex virus type 1 mutant amb1511-7 causes extensive cell fusion. J.
Virol. 67(4), 2396-401.
Baines, J. D., Ward, P. L., Campadelli-Fiume, G., and Roizman, B. (1991). The UL20
gene of herpes simplex virus 1 encodes a function necessary for viral egress. J.
Virol. 65(12), 6414-24.
162
Bond, V. C., and Person, S. (1984). Fine structure physical map locations of alterations
that affect cell fusion in herpes simplex virus type 1. Virology 132(2), 368-76.
Bzik, D. J., Fox, B. A., DeLuca, N. A., and Person, S. (1984). Nucleotide sequence of a
region of the herpes simplex virus type 1 gB glycoprotein gene: mutations
affecting rate of virus entry and cell fusion. Virology 137(1), 185-90.
Cai, W., Person, S., Warner, S. C., Zhou, J., and DeLuca, N. A. (1987). Linker-insertion
nonsense and restriction-site deletion mutations of the gB glycoprotein gene of
herpes simplex virus type 1. J. Virol. 61, 714-721.
Cai, W. H., Gu, B., and Person, S. (1988). Role of glycoprotein B of herpes simplex virus
type 1 in viral entry and cell fusion. J. Virol. 62(8), 2596-604.
Cai, W. Z., Person, S., DebRoy, C., and Gu, B. H. (1988). Functional regions and
structural features of the gB glycoprotein of herpes simplex virus type 1. An
analysis of linker insertion mutants. J. Mol. Biol. 201(3), 575-88.
Chang, Y. E., Menotti, L., Filatov, F., Campadelli-Fiume, G., and Roizman, B. (1998).
UL27.5 is a novel gamma2 gene antisense to the herpes simplex virus 1 gene
encoding glycoprotein B. J Virol 72(7), 6056-64.
Claesson-Welsh, L., and Spear, P. G. (1986). Oligomerization of herpes simplex virus
glycoprotein B. J. Virol. 60, 803-806.
Claesson-Welsh, L., and Spear, P. G. (1987). Amino-terminal sequence, synthesis, and
membrane insertion of glycoprotein B of herpes simplex virus type 1. J. Virol. 61,
1-7.
Davis-Poynter, N., Bell, S., Minson, T., and Browne, H. (1994). Analysis of the
contributions of herpes simplex virus type 1 membrane proteins to the induction
of cell-cell fusion. J. Virol. 68(11), 7586-90.
Debroy, C., Pederson, N., and Person, S. (1985). Nucleotide sequence of a herpes
simplex virus type 1 gene that causes cell fusion. Virology 145(1), 36-48.
Desai, P., DeLuca, N. A., Glorioso, J. C., and Person, S. (1993). Mutations in herpes
simplex virus type 1 genes encoding VP5 and VP23 abrogate capsid formation
and cleavage of replicated DNA. J. Virol. 67(3), 1357-64.
Dietz, P., Klupp, B. G., Fuchs, W., Kollner, B., Weiland, E., and Mettenleiter, T. C.
(2000). Pseudorabies virus glycoprotein K requires the UL20 gene product for
processing. J Virol 74(11), 5083-90.
Forrester, A., Farrell, H., Wilkinson, G., Kaye, J., Davis-Poynter, N., and Minson, T.
(1992). Construction and properties of a mutant of herpes simplex virus type 1
with glycoprotein H coding sequences deleted. J Virol 66(1), 341-8.
163
Foster, T. P., Alvarez, X., and Kousoulas, K. G. (2003). Plasma membrane topology of
syncytial domains of herpes simplex virus type 1 glycoprotein K (gK): the UL20
protein enables cell surface localization of gK but not gK-mediated cell-to-cell
fusion. J Virol 77(1), 499-510.
Foster, T. P., and Kousoulas, K. G. (1999). Genetic analysis of the role of herpes simplex
virus type 1 glycoprotein K in infectious virus production and egress. J Virol
73(10), 8457-68.
Foster, T. P., Melancon, J. M., Baines, J. D., and Kousoulas, K. G. (2004a). The Herpes
Simplex Virus Type 1 UL20 Protein Modulates Membrane Fusion Events during
Cytoplasmic Virion Morphogenesis and Virus-Induced Cell Fusion. J Virol
78(10), 5347-57.
Foster, T. P., Melancon, J. M., and Kousoulas, K. G. (2001). An alpha-helical domain
within the carboxyl terminus of herpes simplex virus type 1 (HSV-1) glycoprotein
B (gB) is associated with cell fusion and resistance to heparin inhibition of cell
fusion. Virology 287(1), 18-29.
Foster, T. P., Melancon, J. M., Olivier, T. L., and Kousoulas, K. G. (2004b). Herpes
simplex virus type-1 (HSV-1) glycoprotein K (gK) and the UL20 protein are
interdependent for intracellular trafficking and trans-Golgi network localization. J
Virol, (In Press).
Foster, T. P., Rybachuk, G. V., and Kousoulas, K. G. (1998). Expression of the enhanced
green fluorescent protein by herpes simplex virus type 1 (HSV-1) as an in vitro or
in vivo marker for virus entry and replication. J Virol Methods 75(2), 151-60.
Foster, T. P., Rybachuk, G. V., and Kousoulas, K. G. (2001). Glycoprotein K specified
by herpes simplex virus type 1 is expressed on virions as a Golgi complexdependent glycosylated species and functions in virion entry. J Virol 75(24),
12431-8.
Fuchs, W., Klupp, B. G., Granzow, H., and Mettenleiter, T. C. (1997). The UL20 gene
product of pseudorabies virus functions in virus egress. J Virol 71(7), 5639-46.
Gage, P. J., Levine, M., and Glorioso, J. C. (1993). Syncytium-inducing mutations
localize to two discrete regions within the cytoplasmic domain of herpes simplex
virus type 1 glycoprotein B. J. Virol. 67(4), 2191-201.
Haan, K. M., Lee, S. K., and Longnecker, R. (2001). Different functional domains in the
cytoplasmic tail of glycoprotein B are involved in Epstein-Barr virus-induced
membrane fusion. Virology 290(1), 106-14.
Highlander, S. L., Goins, W. F., Person, S., Holland, T. C., Levine, M., and Glorioso, J.
C. (1991). Oligomer formation of the gB glycoprotein of herpes simplex virus
type 1. J Virol 65(8), 4275-83.
164
Horsburgh, B. C., Hubinette, M. M., Qiang, D., MacDonald, M. L., and Tufaro, F.
(1999). Allele replacement: an application that permits rapid manipulation of
herpes simplex virus type 1 genomes. Gene Ther 6(5), 922-30.
Hutchinson, L., Goldsmith, K., Snoddy, D., Ghosh, H., Graham, F. L., and Johnson, D.
C. (1992). Identification and characterization of a novel herpes simplex virus
glycoprotein, gK, involved in cell fusion. J Virol 66(9), 5603-9.
Hutchinson, L., Graham, F. L., Cai, W., Debroy, C., Person, S., and Johnson, D. C.
(1993). Herpes simplex virus (HSV) glycoproteins B and K inhibit cell fusion
induced by HSV syncytial mutants. Virology 196(2), 514-31.
Hutchinson, L., and Johnson, D. C. (1995). Herpes simplex virus glycoprotein K
promotes egress of virus particles. J. Virol. 69(9), 5401-13.
Jacobson, J. G., Chen, S. H., Cook, W. J., Kramer, M. F., and Coen, D. M. (1998).
Importance of the herpes simplex virus UL24 gene for productive ganglionic
infection in mice. Virology 242(1), 161-9.
Jayachandra, S., Baghian, A., and Kousoulas, K. G. (1997). Herpes simplex virus type 1
glycoprotein K is not essential for infectious virus production in actively
replicating cells but is required for efficient envelopment and translocation of
infectious virions from the cytoplasm to the extracellular space. J. Virol. 71(7),
5012-24.
Klupp, B. G., Nixdorf, R., and Mettenleiter, T. C. (2000). Pseudorabies virus
glycoprotein M inhibits membrane fusion. J Virol 74(15), 6760-8.
Kousoulas, K. G., Person, S., and Holland, T. C. (1978). Timing of some of the molecular
events required for cell fusion induced by herpes simplex virus type 1. J. Virol.
27(3), 505-12.
Koyano, S., Mar, E. C., Stamey, F. R., and Inoue, N. (2003). Glycoproteins M and N of
human herpesvirus 8 form a complex and inhibit cell fusion. J Gen Virol 84(Pt 6),
1485-91.
Laquerre, S., Person, S., and Glorioso, J. C. (1996). Glycoprotein B of herpes simplex
virus type 1 oligomerizes through the intermolecular interaction of a 28-aminoacid domain. J Virol 70(3), 1640-50.
Ligas, M. W., and Johnson, D. C. (1988). A herpes simplex virus mutant in which
glycoprotein D sequences are replaced by beta-galactosidase sequences binds to
but is unable to penetrate into cells. J. Virol. 62(5), 1486-94.
Luna, R. E., Zhou, F., Baghian, A., Chouljenko, V., Forghani, B., Gao, S. J., and
Kousoulas, K. G. (2004). Kaposi's Sarcoma-Associated Herpesvirus Glycoprotein
K8.1 Is Dispensable for Virus Entry. J Virol 78(12), 6389-98.
165
MacLean, C. A., Efstathiou, S., Elliott, M. L., Jamieson, F. E., and McGeoch, D. J.
(1991). Investigation of herpes simplex virus type 1 genes encoding multiply
inserted membrane proteins. J. Gen. Virol. 72(Pt 4), 897-906.
Melancon, J. M., Foster, T. P., and Kousoulas, K. G. (2004). Genetic analysis of the
herpes simplex virus type 1 (HSV-1) UL20 protein domains involved in
cytoplasmic virion envelopment and virus-induced cell fusion. J Virol 78(14),
7329-43.
Mettenleiter, T. C. (2002a). Brief overview on cellular virus receptors. Virus Res 82(1-2),
3-8.
Mettenleiter, T. C. (2002b). Herpesvirus assembly and egress. J Virol 76(4), 1537-47.
Moraes, M. P., Mayr, G. A., and Grubman, M. J. (2001). pAd5-Blue: direct ligation
system for engineering recombinant adenovirus constructs. Biotechniques 31(5),
1050, 1052, 1054-6.
Narayanan, K., Williamson, R., Zhang, Y., Stewart, A. F., and Ioannou, P. A. (1999).
Efficient and precise engineering of a 200 kb beta-globin human/bacterial
artificial chromosome in E. coli DH10B using an inducible homologous
recombination system. Gene Ther 6(3), 442-7.
Orford, M., Nefedov, M., Vadolas, J., Zaibak, F., Williamson, R., and Ioannou, P. A.
(2000). Engineering EGFP reporter constructs into a 200 kb human beta-globin
BAC clone using GET Recombination. Nucleic Acids Res 28(18), E84.
Pellett, P. E., Kousoulas, K. G., Pereira, L., and Roizman, B. (1985). Anatomy of the
herpes simplex virus 1 strain F glycoprotein B gene: primary sequence and
predicted protein structure of the wild type and of monoclonal antibody-resistant
mutants. J. Virol. 53(1), 243-53.
Pertel, P. E. (2002). Human herpesvirus 8 glycoprotein B (gB), gH, and gL can mediate
cell fusion. J Virol 76(9), 4390-400.
Pogue-Geile, K. L., Lee, G. T., Shapira, S. K., and Spear, P. G. (1984). Fine mapping of
mutations in the fusion-inducing MP strain of herpes simplex virus type 1.
Virology 136(1), 100-9.
Pogue-Geile, K. L., and Spear, P. G. (1987). The single base pair substitution responsible
for the syn phenotype of herpes simplex virus type 1, strain MP. Virology 157,
67-74.
Ramaswamy, R., and Holland, T. C. (1992). In vitro characterization of the HSV-1 UL53
gene product. Virology 186(2), 579-87.
Reynolds, A. E., Wills, E. G., Roller, R. J., Ryckman, B. J., and Baines, J. D. (2002).
Ultrastructural localization of the herpes simplex virus type 1 UL31, UL34, and
166
US3 proteins suggests specific roles in primary envelopment and egress of
nucleocapsids. J Virol 76(17), 8939-52.
Roizman, B., and Sears, A. E. (2001). Herpes Simplex Viruses and Their Replication.
Third edition ed. In "Fields Virology" (D. M. Knipe, and P. M. Howley, Eds.),
Vol. 2, pp. 2399-2459. 2 vols. Lippincott-Williams & Wilkins, Philadelphia, PA.
Roop, C., Hutchinson, L., and Johnson, D. C. (1993). A mutant herpes simplex virus type
1 unable to express glycoprotein L cannot enter cells, and its particles lack
glycoprotein H. J. Virol. 67(4), 2285-97.
Ryechan, W. T., Morse, L. S., Knipe, D. M., and Roizman, B. (1979). Molecular genetics
of herpes simplex virus. II. Mapping of the major viral glycoproteins and of the
genetic loci specifying the social behavior of infected cells. J Virol. 29, 677697.
Saeki, Y., Ichikawa, T., Saeki, A., Chiocca, E. A., Tobler, K., Ackermann, M.,
Breakefield, X. O., and Fraefel, C. (1998). Herpes simplex virus type 1 DNA
amplified as bacterial artificial chromosome in Escherichia coli: rescue of
replication-competent virus progeny and packaging of amplicon vectors. Hum
Gene Ther 9(18), 2787-94.
Sanders, P. G., Wilkie, N. M., and Davison, A. J. (1982). Thymidine kinase deletion
mutants of herpes simplex virus type 1. J. Gen. Virol. 63(2), 277-95.
Spear, P. G. (1993). Membrane fusion induced by herpes simplex virus. In "Viral fusion
mechanisms" (J. Bentz, Ed.), pp. 201-232. CRC Press., Boca Raton, Fla.
Spear, P. G. (2004). Herpes simplex virus: receptors and ligands for cell entry. Cell
Microbiol 6(5), 401-10.
Spear, P. G., and Longnecker, R. (2003). Herpesvirus entry: an update. J Virol 77(19),
10179-85.
Stavropoulos, T. A., and Strathdee, C. A. (1998). An enhanced packaging system for
helper-dependent herpes simplex virus vectors. J Virol 72(9), 7137-43.
Tanaka, M., Kagawa, H., Yamanashi, Y., Sata, T., and Kawaguchi, Y. (2003).
Construction of an excisable bacterial artificial chromosome containing a fulllength infectious clone of herpes simplex virus type 1: viruses reconstituted from
the clone exhibit wild-type properties in vitro and in vivo. J Virol 77(2), 1382-91.
167
CHAPTER IV
GENETIC ANALYSIS OF THE HERPES SIMPLEX VIRUS TYPE 1
(HSV-1) UL20 PROTEIN DOMAINS INVOLVED IN CYTOPLASMIC
VIRION ENVELOPMENT AND VIRUS-INDUCED CELL FUSION*
INTRODUCTION
Herpes simplex viruses (HSV) specify at least eleven virally encoded
glycoproteins, as well as several non-glycosylated membrane associated proteins, most of
which play important roles in multiple membrane fusion events during virus entry and
intracellular virion morphogenesis and egress (Roizman and Sears, 1996; Spear, 1993a;
Spear, 1993b; Spear, Eisenberg, and Cohen, 2000). Spread of infectious virus occurs
either by release of virions to extracellular spaces or through virus-induced cell-to-cell
fusion. Mutations that cause extensive virus-induced cell fusion predominantly arise in
four genes of the HSV genome: the UL20 gene (Baines et al., 1991; MacLean et al.,
1991), the UL24 gene (Jacobson et al., 1998; Sanders, Wilkie, and Davison, 1982), the
UL27 gene encoding glycoprotein B (gB) (Bzik et al., 1984; Pellett et al., 1985), and the
UL53 gene coding for glycoprotein K (gK) (Bond and Person, 1984; Debroy, Pederson,
and Person, 1985; Hutchinson et al., 1992; Pogue-Geile et al., 1984; Ryechan et al.,
1979). Of these four membrane associated proteins, only UL20 and gK are essential for
the intracellular transport of virions to extracellular spaces in all cell types (Baines et al.,
*
Reprinted from the Journal of Virology, Vol 78, J. M. Melancon, T. P. Foster, and K. G. Kousoulas,
Genetic analysis of the herpes simplex virus type 1 UL20 protein domains involved in cytoplasmic virion
envelopment and virus-induced cell fusion, Pages 7329-7343, Copyright 2004, with permission from the
American Society for Microbiology.
168
1991; Foster and Kousoulas, 1999; Fuchs et al., 1997; Hutchinson and Johnson, 1995;
Jayachandra, Baghian, and Kousoulas, 1997).
Virus morphogenesis and egress of infectious herpes virions is thought to involve
sequential de-envelopment and re-envelopment steps in transit to extracellular spaces: a)
primary envelopment by budding of capsids assembled in the nuclei through the inner
nuclear leaflet leading to the production of enveloped virions within perinuclear spaces; b)
de-envelopment by fusion of viral envelopes with the outer nuclear leaflet leading to the
accumulation of unenveloped capsids in the cytoplasm; c) re-envelopment of cytoplasmic
capsids into TGN-derived vesicles. This final event in cytoplasmic virion envelopment is
thought to be mediated by interactions between tegument proteins and cytoplasmic
portions of viral glycoproteins embedded within the TGN-derived membranes.
Cytoplasmically enveloped viruses are thought to be transported to extracellular spaces
within Golgi or TGN-derived vesicles (reviewed in: Johnson and Huber, 2002;
Mettenleiter, 2002; Tomishima, Smith, and Enquist, 2001).
The UL20 gene encodes a 222 amino acid non-glycosylated transmembrane
protein that is conserved in all alphaherpesviruses. The UL20p is a structural component
of extracellular enveloped virions and it is expressed in infected cells assuming a
predominantly perinuclear and cytoplasmic distribution (Ward et al., 1994). A partial
deletion of the UL20 gene was reported to form weakly syncytial viral plaques in certain
cell types and severe defects in cytoplasmic virion envelopment implying that UL20
functioned in virus-induced cell fusion and virion morphogenesis. Furthermore,
replication of the UL20-null virus could be partially complemented in certain cell types
indicating a cellular specificity of UL20 functions (Baines et al., 1991). This UL20p
169
cellular specificity was phenomenologically associated with disruption of the Golgi
apparatus during viral infections, inasmuch as, cells that complemented the UL20-null
virus retained intact Golgi, while the Golgi of cells failing to complement the UL20-null
virus were disrupted and dispersed throughout the cytoplasm (Avitabile et al., 1995;
Avitabile et al., 1994).
Recently, we showed that a precise deletion of the UL20 gene caused
accumulation of unenveloped capsids into the cytoplasm indicating that the HSV-1
UL20p functioned in cytoplasmic stages of virion envelopment and that the previously
reported partial deletion of the UL20 gene caused a predicted fusion of the UL20 gene to
the adjacent UL20.5 gene. In addition, syncytial mutations in either gB or gK failed to
cause fusion in the absence of the UL20 gene, suggesting that the UL20 protein was
essential for virus-induced cell fusion (Foster et al., 2004). Furthermore, we showed that
UL20 is required for cell-surface expression of gK, suggesting a functional
interdependence between gK and UL20 for virus egress and cell-to-cell fusion (Dietz et
al., 2000; Foster, Alvarez, and Kousoulas, 2003). In this manuscript we have delineated
via site-directed mutagenesis the functional domains of UL20p involved in infectious
virus production and virus-induced cell fusion. Importantly, we show that both amino and
carboxyl terminal portions of UL20p, which are predicted to lie within the cytoplasmic
side of cellular membranes, function both in cytoplasmic virion envelopment and virusinduced cell fusion. Overall, the data suggest that UL20p’s role in virus-induced cell
fusion can be functionally separated from its role in cytoplasmic virion morphogenesis,
and that UL20p domains that function in gB-syn3 virus-induced cell fusion are distinct
from those functioning in gKsyn1 virus-induced cell fusion.
170
MATERIALS AND METHODS
Cells and Viruses
African green monkey kidney (Vero) cells were obtained from ATCC (Rockville,
MD). The Vero-based UL20 complementing cell line, G5, was a gift of Dr. P. Desai,
(John Hopkins Medical Center) (Desai et al., 1993). Cells were maintained as previously
described (Desai et al., 1993; Foster, Alvarez, and Kousoulas, 2003; Foster and
Kousoulas, 1999). The parental wild-type strain used in this study HSV-1 (KOS) was
originally obtained from P. A. Schaffer (Harvard Medical School). Flp-In-CV-1 cells
were maintained as directed by the manufacturer (Invitrogen, Inc.). ∆20DIV5,
∆20gBsyn3 and ∆20syn20DIV5 viruses were as described previously (Foster et al., 2004),
and virus stocks were grown on Fd20-1 cells, described below. In this paper, for
simplification purposes, the ∆20DIV5 virus is referred to as ∆20 virus and the
∆20syn20DIV5 virus is referred to as ∆20gKsyn1 virus (the syn20 and syn1 mutation are
the same, Ala-to-Val at gK amino acid 40).
Construction of Transformed Flp-In-CV-1 Cell Lines
Generation of stable Flp-In-CV-1 expression cell lines was performed essentially
as directed by the manufacturer (Invitrogen). Briefly, confluent Flp-In-CV-1 monolayers
in six well plates were transfected using Lipofectamine 2000 with 0.3 µg of either
pcDNA5/FRT/UL20 or pFRT/gDpro/UL20 and 2.7 µg of pOG44 (Invitrogen), which
expresses Flp recombinase. After 2 weeks, hygromycin B (125 µg/ml) resistant colonies
were tested for the ability to complement the growth of ∆20, ∆20gBsyn3 and ∆20gKsyn1
viruses. All colonies derived from each plasmid complemented UL20 null virus. Isolates
Fc20-1 and Fd20-1, derived from pcDNA5/FRT/UL20 and pFRT/gDpro/UL20
171
transfected cells, respectively, were chosen for further use. Fd20-1 cells were used in
generating UL20-null virus stocks.
Plasmids
pCR2.1-UL20, which was used as the parental vector for UL20 mutagenesis, was
generated by cloning a 773bp DNA fragment containing the UL20 gene, obtained by PCR
amplification of HSV-1(KOS) viral DNA, into pCR2.1/TOPO (Invitrogen). A 7,185bp
fragment spanning the region comprising UL19, UL20, UL21, and UL22 was PCR
amplified from KOS viral DNA using primers p20F2 5’ (TTTCTTGAATTCACGACGG
CGGTGTAGCCCACG) and p20F2 3’ (CTTCTTGAATTCCCCATCACCCACAACGC
CAGC), EcoRI restricted, and ligated into puc19BX (Foster et al., 2004) to generate
p20F2. p20F2 was modified by silencing the BamHI site at genomic nucleotide position
39235 (Genbank accession number NC_001806), conserving the UL19 amino acid
sequence, and removing a 767 bp BamHI DNA fragment containing the UL20 gene,
resulting in plasmid p20F2BX (see Fig 4.2: D).
UL20 cluster to alanine mutants, single point mutants, and truncation mutants
were generated by two methods. Mutants CL11, CL23 , CL34, CL38, CL121, CL209, 66t,
and 149t were generated using the GeneTailor™ Site-Directed Mutagenesis Kit as
directed by the manufacturer (Invitrogen). Mutants CL5, CL16, CL30, CL41, CL46,
CL49, CL153, CL173, CL177, 181t, 204t, 211t, 216t, Y49A, S50A, R51A, D123A,
R209A, T212A, and R213A were generated by splice-overlap extension (SOE) PCR
(Aiyar, Xiang, and Leis, 1996). Mutant UL20 genes were transferred into the BamHI site
of the vector p20F2BX. All p20F2BX mutant constructs were sequenced to verify error
free mutagenesis. Plasmid p20R contains a wild type UL20 gene cloned into p20F2BX.
172
An additional plasmid was constructed from p20F2BX, designated p∆20NE, to include
the same UL20 deletion as p∆20-EGFP (Foster et al., 2004) without the EGFP gene
cassette. Plasmid pcDNA5/FRT/UL20 was constructed by inserting the UL20 gene, PCR
amplified from KOS viral DNA, into pcDNA5/FRT/V5-His-TOPO (Invitrogen). An
additional plasmid, pFRT/gDpro/UL20, was constructed by replacing the CMV promoter
in pcDNA5/FRT/UL20 with the HSV-1 KOS gD promoter region.
UL20 Complementation Assay for Infectious Virion Production
The complementation assay was modified from a similar assay described
previously (Bjerke et al., 2003). Confluent Vero monolayers in six well plates were
transfected with 1.5 µg of pcDNA3.1/V5-His-TOPO/lacZ (Invitrogen), expressing βgalactosidase under control of the CMV promoter, and 1.5 µg of wild-type or mutant
UL20 plasmid with Lipofectamine 2000 as described by the manufacturer (Invitrogen).
Six hours post-transfection, the monolayers were infected with a UL20-null virus at an
MOI of 1. Infections were placed on a rocker for 1 hour at 4°C, and then transferred to
37°C for 2 hours. Residual virus was inactivated using an acid wash (PBS containing .5M
glycine, pH3) for 2 min, and monolayers were subsequently washed 3 times with DMEM
to restore the pH to a normal level. Infections were incubated at 37°C for 24 hours. After
repeated freeze/thaw cycles, virus stocks were titered in triplicate on G5 cells, which
effectively complement the UL20-null defect (Foster et al., 2004). An aliquot of each
virus stock was saved to assay for β-galactosidase expression as described below.
β-galactosidase expression levels were quantified essentially as described
previously (Bjerke et al., 2003). 100 µl of Z buffer containing 0.27% β−mercaptoethanol
and 0.2% Triton X-100 was added to 100 µl of virus stock and vortexed for 30 s. Cell
173
debris was pelleted by centrifugation at 16,100 g in a microcentrifuge for 30 s. 160 µl of
the supernatant was transferred to a 96-well plate and reacted with 40 ul of 4 mg/ml οnitrophenyl-β−D-galactopyranoside (ONPG) at room temperature. The reactions were
allowed to proceed for 90 minutes before determining absorbance at 405 nm.
Calculation of Complementation Ratios
The complementation ratio for each mutant was calculated with the formula [(virus
titer of mutant / virus titer of negative control)*(experimental β-galactosidase value / βgalactosidase value of negative control)]. Using this formula, values obtained from the βgalactosidase assay were used to correct for minor differences in transfection efficiency;
and the complementation ratio for the negative control (plasmid p∆20NE) was set at 1.
UL20 Complementation Assay for Virus-Induced Cell-to-Cell Fusion
Confluent Vero monolayers in six-well plates were transfected with 3 µg of wildtype or mutant UL20 plasmid with Lipofectamine 2000 as described by the manufacturer
(Invitrogen). 18 hours post transfection, the monolayers were infected at an MOI of 0.1
with either ∆20gKsyn1 or ∆20gBsyn3 viruses. Infections were placed on a rocker at room
temperature for 1 hour, then transferred to 37° C for 30 minutes. Cells were overlaid with
DMEM containing 1% methylcellulose. 24 hours post-infection, cell fusion was
determined by visualization of syncytia formation by fluorescence microscopy. Cell
fusion was blindly scored by two independent observers and scores were linked to
individual mutants after the cell fusion scoring was completed. Cell fusion was scored as
completely absent (--), or present at very low (+), moderate (++), near (+++) or at levels
equivalent (++++) to those observed when the wild-type UL20 gene was used. The score
of (-) indicates no syncytial formation as determined by the absence of cells having more
174
than one nucleus. The score of (+) indicates the presence of syncytia containing on the
average 5 nuclei or less. The score of (++) reflects syncytia containing 6-10 nuclei per
cell. The score of (+++) is assigned to 11-20 nuclei per cell. The score of (++++) is for
syncytia having greater than 20 nuclei per cell.
Electron Microscopy
Cell monolayers were infected with the indicated virus at an MOI of 5. All cells
were prepared for TEM examination 24 hpi. Infected cells were fixed in a mixture of 2 %
paraformaldehyde and 1.5% glutaraldehyde in 0.1M NaCaC buffer, pH 7.3. Following
osmication (1% OsO4) and dehydration in an ethanol series, the samples were embedded
in Epon-Araldyte resin and polymerized at 70º C. Thin sections were made on an MTXL
Ultratone (RMC Products), stained with 5% uranyl acetate and CNA lead, and observed
with a Zeiss 10 transmission electron microscope as described previously (Foster,
Alvarez, and Kousoulas, 2003).
Generation of Recombinant Viruses Specifying Mutant UL20p
Viruses that contained the UL20 mutations were generated by rescuing the UL20null viruses (∆20, ∆20gBsyn3, ∆20gKsyn1) with plasmids containing the desired UL20
mutant genes bracketed by adjacent viral sequences to facilitate homologous
recombination with the viral genome (Fig. 4.3). Briefly, confluent Vero cell monolayers
were transfected with each plasmid and transfected cells were infected with the ∆20 virus
six hours after transfection. Virus stocks were prepared at 48 hpi and recombinant viruses
were plaque isolated on Vero cells and sequentially plaque-purified at least five times.
Replacement of the UL20-null resident CMV-EGFP gene cassette and the concomitant
insertion of the mutated UL20 gene were confirmed by DNA sequencing. Visualization of
175
viral plaques was achieved by either phase contrast microscopy or immunohistochemistry
essentially as described previously (Foster et al., 2004).
RESULTS
Mutagenesis of HSV-1 UL20
To delineate the functional domains of UL20p required for infectious virus
production, virus egress and virus-induced cell fusion, we constructed a panel of 31
mutations within the UL20 gene by site-directed mutagenesis. These mutations included:
1) cluster-to-alanine mutants in which a cluster of proximal amino acids were changed to
alanine residues; 2) single amino acid replacement mutants within alanine cluster regions;
3) carboxyl terminal truncations of UL20p. The mutated amino acids for each type of
mutation are shown in Table 4.1. The carboxyl terminal truncations are identified with the
number of the last remaining amino acid (i.e. 66t retains UL20p amino acids 1-66). To
facilitate placement of each mutation on the predicted luminar or cytoplasmic portions of
UL20p, the hydrophobic and membrane-spanning domains of UL20p were calculated
using the TMPred and SOSUI computer algorithms (Hirokawa, Boon-Chieng, and
Mitaku, 1998; Hofmann and Stoffel, 1993), and used to derive a predicted membrane
spanning model of UL20p (Fig. 4.1). This UL20p model features four membrane
spanning regions placing both amino and carboxyl termini of UL20p within the cytoplasm
(Domains I and V). In addition, a third small domain is located intracellularly (domain
III), while the two other domains are located extracellularly (domains II and IV). The
predicted membrane topology of UL20p was used as the guide for selecting alanine
scanning mutations within predicted intracellular and extracellular domains of UL20p.
176
Table 4.1: Amino acid sequences of mutations
Domain
I
I
I
I
I
I
I
I
I
I
I
I
I
III
IV
IV
IV
IV
IV
IV
IV
V
V
V
V
Mutation Name Wildtype aa sequence Mutant aa sequence
CL5
CL11
CL16
CL23
CL30
CL34
CL38
CL41
CL46
CL49
Y49A
S50A
R51A
CL121
CL153
E153A
T154A
S156A
D158A
CL173
CL177
CL209
R209A
T212A
R213A
DD
DRD
DE
EEGE
EE
SLSS
YGT
SD
SS
YSR
KRDR
ETFSPD
TD
RTR
RFWTR
AA
AAA
AA
AAGA
AA
ALAA
AGA
AA
AA
AAA
ASR
YAR
YSA
AADA
AAFAPA
ATFSPD
EAFSPD
ETFAPD
ETFSPA
AA
AAA
AFWAA
AFWTR
RFWAR
RFWTA
Truncations
66t
149t
181t
204t
211t
216t
VILFL
LFAAA
RALGI
SANFF
RFWTR
AILNA
* indicates stop codon
177
VILG*
LFAG*
RALG*
SANG*
RFWG*
AILG*
Domain IV
D
Extracellular/Luminal
P
CL153
L
90
T G R
G
Y
M
G
A
V
80 M
E
L
P
TM1
K
L
V
L
F
S
70 W
L
F
L
I
V
CL5
D
CL34
20
CL46
F CL30
S
CL16
CL49
A
F
L
S
Y
Q
S
S
M
L
E
E
E
S
V
V 10 D
E
E 30 Y
F
S 50 F 60
R
V
L
P
D
G
L
F
P
E
L
T
G
Y
C
A
A
G
V
L
Y
V
A
V
A
A 150
L
A
G
I
I
Y
T
W
A
R
L
A
CL177
R
A 180
P
L
A
A
L
A
100
G
V
140 L
S
V
110
L
TM2
W
A
A
T
F
M
G
F
S
V
C
A
F
C
L
A
G
W
V
V
TM3
A
F
A
P 130
A
L
Q
I K R D
120
CL121
R
Domain III
P 40 T
D
P
T
CL38 S
H
CL41
210
D
A
A
S
I
A
A
G
M 190
A
TM4
A
S
200
S
N
F
G
CL23
N
I
F
G
L
F
L
A
D
E
H
R
T
V
N
A
R
L
Q
K
S
P
CL11
M
H
P
L
NH2
F
T
Y
R
V
F
D
L 160 L 170 P
R
D
T
1 M
P
S
Domain II
CL173
P T
G
220
V
L
P
A
A
A
F 222
R
F
N
W
I
L
T
COOH
A
R
CL209
Domain V
Domain I
Intracellular/Cytoplasmic
Figure 4.1: Predicted membrane topology of UL20p and location of the 15 clusterto-alanine mutations. Membrane topology was predicted using the TMPred and SOSUI
algorithms (19, 20). UL20p domains where cluster-to-alanine mutations are located are
indicated by a shaded oval. Naming of cluster mutations is based on the first amino acid
mutated in each cluster. Transmembrane region (TM), Cluster mutant (CL).
178
Regions containing clusters of charged amino acids as well as potential phosphorylation
sites were specifically targeted. The location of each cluster mutation is marked on the
UL20p model starting with the first mutated amino acid of the cluster (Fig. 4.1, Table
4.1).
Complementation Assay for Infectious Virus Production
Previous studies showed that deletion of the HSV-1 UL20 and PRV UL20 genes
resulted in up to two logs reduction in infectious virus production relative to their parental
wild type strains (Baines et al., 1991; Foster et al., 2004; Fuchs et al., 1997). To delineate
functional domains of the UL20 protein involved in infectious virus production, each of
the 31 mutated UL20 genes was tested for its ability to complement the HSV-1(KOS)
UL20-null virus. The mutated genes were cloned into plasmids under their own promoter
control bracketed by upstream and downstream DNA sequences to facilitate
complementation experiments as well as isolation of recombinant viruses (Fig. 4.2: D, E;
see Materials and Methods). Complementation experiments involved transfection of Vero
cells with plasmids encoding wild-type or mutant UL20 genes, followed by infection with
the UL20-null virus (Foster et al., 2004). A complementation ratio was calculated for each
mutant UL20 plasmid, p20R (wild-type UL20, positive control), and p∆20NE (UL20-null
mutation, negative control). The complementation ratio of p∆20NE was set to 1 for each
experiment. The complementation ratios for the UL20 cluster mutants and truncation
mutants are shown in Figure 3A. Mutants CL5, CL11, CL16, CL23, CL30, CL34, CL38,
CL41 CL46, CL121, CL173, CL177, and CL209 complemented the UL20-null virus to a
variable extent in comparison to the positive plasmid control p20R and the negative
plasmid control p∆20NE (Fig. 4.3A). Mutants CL49, CL153, 66t, 149t, 181t, 204t, 211t,
179
UL
TR
IRL IRS
US
TR
A
0.0
0.2
0.1
0.3
0.4
0.5
0.6
0.8
0.7
0.9
1.0
∆UL20 virus
UL19 (VP5)
B
C
UL19 (VP5)
p20F2BX-UL20mut
EcoRI
D
EGFP
CMV
UL20Mutant
GGATCA BamHI
(BamHI/BclI)
UL19 (VP5)
p20F2BX
EcoRI
UL20.5
UL20.5
UL21
UL22 (gH)
UL21
EcoRI
BamHI
UL20.5
UL21
EcoRI
GGATCA BamHI
(BamHI/BclI)
UL20Mutant
E
BamHI
BamHI
Figure 4.2: Schematic of the strategy for the construction of UL20-mutant genes
used for complementation and isolation of recombinant viruses. (A) The top line
represents the prototypic arrangement of the HSV-1 genome with the unique long (UL)
and unique short (US) regions flanked by the terminal repeat (TR) and internal repeat (IR)
regions. (B) Shown below is and expanded genomic region of the UL20-null virus
between map units 0.239 and 0.305 containing the UL19, CMV-EGFP, UL20.5, UL21,
and UL22 genes. (C) The p20F2BX-UL20mut plasmid denotes the expression and
recombination plasmids used for complementation assays and construction of the
recombinant viruses carrying each UL20 mutation. (D, E) Mutated UL20 genes were
inserted into the BamHI restriction site of p20F2BX.
180
Figure 4.3: Complementation ratios of mutant UL20p genes. Vero cells were
transfected with plasmids encoding wild-type or mutant UL20 genes under the UL20
promoter and then infected with the HSV-1(KOS) UL20-null (∆20) virus. (A) Bar
graph showing complementation ratios for UL20 cluster-to-alanine mutants and carboxyl
terminal truncations. (B) Complementation ratios of the CL49, Y49A, S50A, and R51A
mutants. (C) Complementation ratios of the CL153, E153A, T154A, S156A, and D158A
mutants. (D) Complementation ratios of the CL209, R209A, T212A, and R213A mutants.
The error bars shown represent the maximum and minimum complementation ratios
obtained from three independent experiments, and the bar height represents the average
complementation ratio.
181
and 216t failed to efficiently complement the UL20-null defect in infectious virion
production producing complementation ratios similar to those of plasmid p∆20NE
(negative control) indicating that the mutated UL20 protein was not able to function
properly in virus egress (Fig. 4.3A). To delineate the specific amino acids responsible for
the UL20-associated defects identified by the cluster mutations, we examined the
complementation ratios of several point mutants within cluster regions. The CL49 (YSRto-AAA; Table 4.1) mutant UL20 gene failed to efficiently complement the ∆20 virus.
The Y49A mutant gene, which involves the Y residue within the CL49 target sequence,
produced a complementation ratio similar to CL49, while the adjacent S50A and R51A
mutant genes complemented infectious virus production to nearly wild-type levels (Fig.
4.3B). These results indicate that the mutation of the Y residue is responsible for
complementation defect exhibited by the UL20 gene carrying the CL49 mutation. In
contrast, the UL20 genes carrying the E153A, T154A, S156A, and D158A mutations
complemented the UL20-null virus at a greater level than the CL153 mutation indicating
that these amino acids were not individually responsible for the lack of complementation
by the UL20 gene carrying the CL153 mutation (Fig. 4.3C). The CL209 UL20 mutant
gene as well as mutant genes carrying individual amino acid changes within the CL209
target sequence efficiently complemented ∆20 infectious virus production (Fig. 4.3D).
Complementation for Virus-Induced Cell-to-Cell Fusion
Recently, we showed that syncytial mutations in either gB or gK failed to cause
virus-induced cell fusion in the absence of the UL20 gene (Foster et al., 2004). Therefore,
the panel of 31 UL20 mutants was tested for the ability to complement UL20-null viruses
containing syncytial mutations in either gB (syn3) or gK (syn1) for virus-induced cell
182
fusion. Confluent Vero monolayers were transfected with plasmids encoding either wild
type or mutant UL20p, and subsequently infected with either ∆20gKsyn1 or ∆20gBsyn3
viruses. These viruses contain a CMV-EGFP gene cassette in place of the UL20 gene,
which facilitates visualization of syncytia formation by fluorescence microscopy (17).
Cell fusion was scored by two observers as completely absent (--), or present at very low
(+), moderate (++), near (+++) or at levels equivalent (++++) to those observed when the
wild-type UL20 gene was used. Complementation results are summarized in Table 4.2,
and select mutant phenotypes are displayed in Figure 4.4.
UL20 genes carrying the CL5, CL11, CL16, CL23, CL30, CL34, CL173, CL177,
and CL209 mutations produced syncytia at levels similar to those produced by the
positive control plasmid p20R containing the wild-type UL20 gene. Syncytia formation
was completely absent in complementation assays with UL20 genes specifying the CL153
mutation and all UL20p carboxyl terminal truncations (66t, 149t, 181t, 204t, and 211t)
except the 216t truncation, which complemented virus-induced cell fusion (Fig. 4.4:
panels E, G, I; Table 4.2). Other UL20 mutations had varying phenotypes with regard to
the extent of syncytia formation (Table 4.2). As with the infectious virion production
assay, the phenotype produced by the CL49 mutation was similar to that of Y49A, but not
S50A or R51A (Table 4.2, Fig. 4.4: panels D, F, H, J). Most mutant UL20 genes were
either able or unable to complement both ∆20gBsyn3 and ∆20gKsyn1 viruses, although
the level of complementation varied among the different UL20 mutant genes. However,
the CL41 and CL46 mutant UL20 genes efficiently complemented gBsyn3, but not
gKsyn1 virus-induced cell fusion indicating that these mutations had a differential effect
183
Table 4.2: Complementation results for cell fusion.
Domain
I
I
I
I
I
I
I
I
I
I
I
I
I
III
IV
IV
IV
IV
IV
IV
IV
V
V
V
V
Mutation Name
UL20-null complementation for cell fusion
gK (syn1)
gB (syn3)
++++
++++
++++
++++
+++
+++
+
++
++
++
++
++++
++++
++
-+++
++++
++++
++++
+++
++++
+++
++++
++++
+++
++++
++++
++++
++++
++++
+++
+
++++
++++
++
++
++++
++++
++
-++++
++++
++++
++++
++++
++++
++++
++++
++++
++++
-----+
-----+++
CL5
CL11
CL16
CL23
CL30
CL34
CL38
CL41
CL46
CL49
Y49A
S50A
R51A
CL121
CL153
E153A
T154A
S156A
D158A
CL173
CL177
CL209
R209A
T212A
R213A
Truncations
66t
149t
181t
204t
211t
216t
The extent of cell fusion was scored by two observers as completely absent (--), or
present at very low levels (+), moderate levels (++), near (+++) or at (++++) levels
similar to those obtained with the wild-type UL20 gene.
184
Figure 4.4: Complementation for virus-induced cell-to-cell fusion of UL20-null
viruses containing syncytial mutations in gK(syn1) or gB(syn3). Vero cells were
transfected with plasmids encoding wild-type or mutant UL20 genes and then infected
with either the ∆20gKsyn1 or ∆20gBsyn3 viruses. Twenty-four hpi, cell fusion was
determined by visualization of syncytia formation by fluorescence microscopy. The extent
of syncytial formation (complementation) obtained with the negative control plasmid,
p∆20NE (A) and positive control plasmid, p20R (B) are shown for reference purposes.
Representative images for the CL38, 204t, 211t, and 216t mutants (C, E, G, I,
respectively), and for the CL49, Y49A, S50A, and R51A (D, H, F, J, respectively) are
shown.
185
186
of gBsyn3 versus gKsyn1-induced cell fusion (Table 4.2). The CL38 mutation, which is
immediately proximal to the CL41 mutation, had an obvious defect in complementation
for both gBsyn3 and gKsyn1-induced cell fusion (Table 4.2, Figure 4C).
Plaque Phenotypes of ∆20gBsyn3 and ∆20gKsyn1 Recombinants Containing Selected
UL20 Mutations
To further investigate the ability of UL20 mutant genes to complement for virusinduced cell fusion caused by the gBsyn3 or gKsyn1 virally encoded mutations, specific
recombinant viruses expressing UL20p mutant genes were isolated by rescue of the UL20
gene deletion of the ∆20gBsyn3 and ∆20gKsyn1 viruses. Recombinant viruses were
identified by a lack of EGFP expression on Vero cells. Viral plaques produced by each
recombinant virus were characterized as small plaques, if they contained on the average
no more than 75 cells, or large plaques, if they contained more than 250 cells per plaque.
As expected, we were unable to isolate recombinants expressing the CL153, 66t, 149t,
181t, 204t, or 211t mutations on Vero cells, while rescue with the wild-type UL20 gene
resulted in viruses that formed large syncytial plaques similar to the viruses carrying the
gBsyn3 or gKsyn1 mutations alone (Fig. 4.5: panels A, G). Recombinant viruses carrying
the gKsyn1 mutation and either the CL41 or CL46 UL20 mutation produced small
syncytial plaques on Vero cells (Fig. 4.5: panels C, D), while the recombinant viruses
carrying the gBsyn3 mutation and either the CL41 or CL46 mutation produced large
syncytial plaques on Vero cells (Fig. 4.5: panels I, J). Recombinant viruses containing the
CL38 mutation (Fig. 4.5, panels B, H), or the CL49 mutation (Fig. 4.5, panels E, K) in the
gBsyn3 or gKsyn1 genetic background produced significantly smaller syncytial plaques
than the gBsyn3 or gKsyn1 viruses specifying the wild-type UL20 gene. The gBsyn3 or
187
Figure 4.5: Plaque phenotypes of ∆20gKsyn1 (A, B, C, D, E, F) and ∆20gBsyn3 (G,
H, I, J, K, L) derived recombinant viruses containing selected UL20 mutations on
Vero cells. Confluent cell monolayers were infected with the recombinant viruses
containing wild-type UL20 (A, G), or the UL20 mutants CL38 (B, H), CL41 (C, I), CL46
(D, J), CL49 (E, K), or 216t (F, L), and viral plaques were visualized by
immunohistochemistry at 24 hpi.
188
gKsyn1 recombinant viruses specifying the 216t mutation produced syncytial plaques
similar in size to the gBsyn3 or gKsyn1 viruses (Fig. 4.5: panels F, L).
Plaque Phenotypes of Selected UL20 Mutations in the Context of Wild-Type gB and
gK Genes
Attempts were made to isolate recombinant viruses specifying each of the 31
engineered UL20 mutation in the ∆20 genetic background specifying wild-type gB and
gK genes. As expected, the CL153 mutation and all of the UL20p truncations except 216t
failed to yield recombinant viruses on Vero cells. Most other UL20 mutations produced
non-syncytial virus plaques on Vero cells of similar size to recombinant viruses produced
by rescuing the ∆20 virus with the wild-type UL20 allele in agreement with initial
complementation results. Interestingly, recombinant viruses were isolated containing the
CL49, Y49A, and 216t mutations, which allowed complementation for virus-induced cell
fusion, but exhibited deficiencies in infectious virus production (Fig. 4.3: panels A, B;
Table 4.2; Fig. 4.4: panels D, F, I). Recombinant viruses containing the CL49, Y49A,
216t, CL209, and R209A mutations produced partially syncytial plaques on Vero cells
(Fig. 4.6: panels B1, C1, F1, G1, H1) suggesting that specific UL20 domains were
involved in virus-induced cell fusion. In contrast, these viruses produced non-syncytial
plaques when plated on the G5 cells (Fig. 4.6: panels B2, C2, F2, G2, H2) indicating that
the wild-type UL20 gene expressed by G5 cells was dominant over UL20 syncytial
mutations. In the amino terminus of UL20p, recombinants expressing the S50A and R51A
mutations, which are immediately proximal to the Y49A and contained within the CL49
mutation, did not cause syncytial plaque formation suggesting that mutation of the Y
residue is responsible for the resultant syncytial phenotype (Fig. 4.6: compare panels B1,
189
Figure 4.6: Domains of UL20p associated with UL20p-induced cell fusion. Plaque
phenotypes of recombinant viruses containing selected UL20 mutations are shown
on Vero (A1, B1, C1, D1, E1, F1, G1, H1, I1, J1) and G5 (A2, B2, C2, D2, E2, F2, G2,
H2, I2, J2) cells. Confluent cell monolayers were infected with recombinant viruses
containing wild-type UL20 (A1, A2), or the UL20 mutants CL49 (B1, B2), Y49A (C1,
C2), S50A (D1, D2), R51A (E1, E2), 216t (F1, F2), CL209 (G1, G2), R209A (H1, H2),
T212A (I1, I2), or R213A (J1, J2) at an MOI of 0.001, and viral plaques were visualized
at 30 hpi. White arrows indicate syncytia formation.
190
C1 with D1, E1). In the carboxyl terminus of UL20p, recombinant viruses expressing the
CL209 and R209A mutations produced partially syncytial plaques on Vero cells, while
those expressing the T212A and R213A did not (Fig. 4.5: compare panels G1, H1 with I1,
J1), suggesting that the R residue at position 209 is responsible for the CL209 syncytial
phenotype. On average, syncytia produced by recombinant viruses expressing the
carboxyl terminal syncytial mutations 216t, CL209, and R209A were larger than those
produced by the amino terminal syncytial mutations CL49 and Y49A.
Ultrastructural Characterization of Recombinant Viruses Carrying Mutations
Within the Amino and Carboxyl Termini of UL20p
To evaluate the intracellular virion distribution of the recombinant viruses
specifying the 216t, CL49 and Y49 UL20 mutations, Vero cells infected with UL20p
wild-type and mutant viruses at an MOI of 5 were examined via electron microscopy at 24
hours post infection. As previously shown (Foster et al., 2004), the ∆20 (UL20-null) virus
caused accumulation of unenveloped capsids into the cytoplasm in infected Vero cells
(Fig. 4.7: panel A). In contrast, the ∆20-rescued virions were transported to extracellular
spaces and exhibited no apparent cytoplasmic defects (Fig. 4.7: panel B). Importantly,
infection with the 216t mutant virus resulted in an accumulation of capsids in the
cytoplasm (Figure 7: panels C, D, E, F), similar to the UL20-null phenotype. Similarly,
infection with the CL49 mutant resulted in the accumulation of capsids as well as small
percentage of enveloped particles within the cytoplasm (Figure 8: panels A, B). The
Y49A mutant virus produced the same ultrastructural phenotype as the CL49 mutant (Fig.
8: panel C). The S50A and R51A mutants exhibited no apparent defects in intracellular
morphogenesis, appearing to have an intracellular distribution similar to wild type KOS
virus (Fig. 8: panels D and E).
191
Figure 4.7: Electron micrographs of Vero cells infected with ∆20DIV5 (A), ∆20rescue (B) or 216t (C, D, E, F) viruses. Confluent cell monolayers were infected at an
MOI of 5, incubated at 37°C for 24 hours, and prepared for transmission electron
microscopy. Panel D shows a higher magnification of panel C. Panel F shows a partiallyenveloped capsid often seen in UL20-null infected cells. Nuclear (n), cytoplasmic (c) and
extracellular (e) spaces are marked.
192
193
Figure 4.8: Electron micrographs of Vero cells infected with the CL49 (A, B), Y49Α
(C), S50A (D) or R51A (E) viruses. Confluent cell monolayers were infected at an MOI
of 5, incubated at 37°C for 24 hours, and prepared for transmission electron microscopy.
Nuclear (n), cytoplasmic (c) and extracellular (e) spaces are marked.
194
DISCUSSION
The UL20 gene has been implicated in virion egress, virus-induced cell fusion,
Golgi fragmentation, and Golgi-dependent glycosylation of HSV-1 viral glycoproteins
(Avitabile et al., 1995; Avitabile et al., 1994; Baines et al., 1991; Fuchs et al., 1997).
Recently, we showed that a precise deletion of the UL20 gene caused accumulation of
capsids into the cytoplasm indicating that UL20 functioned in cytoplasmic events of
virion morphogenesis. Moreover, lack of UL20p prevented virus-induced cell fusion
caused by syncytial mutations in either gB or gK suggesting that UL20p plays important
regulatory roles in membrane fusion phenomena associated with virion morphogenesis
and virus-induced cell fusion (Foster et al., 2004). In this report, we constructed and
characterized a panel of 31 different mutations within UL20p to delineate domains of
UL20p functioning in virion morphogenesis and virus-induced cell fusion. Analysis of the
various mutant phenotypes reveals that UL20p contains at least three domains involved in
virion morphogenesis and virus-induced cell fusion: The amino and carboxyl termini of
UL20 (domains I and V), which are predicted to lie on the cytoplasmic side of
membranes, and UL20p domain IV, which is predicted to lie extracellularly. Overall, the
data suggest that the UL20p’s role in virus-induced cell fusion can be functionally
separated from its role in cytoplasmic virion morphogenesis, and that certain UL20p
domains that function in gB-syn3-mediated virus-induced cell fusion are distinct from
those functioning in gKsyn1-mediated virus-induced cell fusion.
The Predicted Membrane Topology of UL20p
Computer assisted prediction of the membrane spanning domains of UL20p
indicated that UL20p spanned membranes four times placing both the amino (domain I)
195
and carboxyl terminal (domain V) portions within the cytoplasm of cellular membranes
and internal to the virion envelope. This predicted structure includes two domains, a very
small domain of 5 amino acids (domain II) and a second domain of 25 amino acids
(domain IV), predicted to be localized in the lumen (extracellular) and external to the
virion envelope. Initial epitope tagging results indicate that domain I is located
intracellularly, while domain IV is located extracellularly in support of this model (data
not shown). However, additional work is required to fully confirm this UL20p topology
model. Domain I is the largest domain (63 aa) and it includes stretches of acidic amino
acid (D, E) clusters, which could form electrostatic interactions with other proteins. These
acidic clusters as well as the amino acid motif YXXΦ (YSRL) have been shown to
function in endocytosis of alphaherpesvirus envelope proteins from plasma membranes to
the TGN (Alconada, Bauer, and Hoflack, 1996; Alconada et al., 1999; Brideau et al.,
1999; Tirabassi and Enquist, 1999; Zhu et al., 1996). The acidic cluster motifs appear to
direct TGN localization by binding to a cellular connector protein, PACS-1, which
connects the glycoproteins to the AP-1 complex (Wan et al., 1998), while the YXXΦ
motif binds adaptor proteins directly (2, 3, 40).
UL20p Domains that Function in Infectious Virus Production and Virus-Induced
Cell Fusion
To delineate the functional domains of UL20p, we constructed a panel of
mutations within the UL20 gene coding for cluster-to-alanine replacements, single amino
acid substitutions and carboxyl terminal truncations. Complementation experiments of the
196
Table 4.3: Differential effects of mutations on infectious virion production and cell
fusion.
Group
Mutants
Complementation for
Infectious Virion
Production
Complementation for Virus-Induced Cell
Fusion
gKsyn1
gBsyn3
I
CL5, CL11, CL16, CL23, CL30,
CL34, CL121, CL173, CL177,
CL209
Yes
High
High
II
CL153, 66t, 149t, 181t, 204t,
211t
No
No
No
III
CL49, 216t
No
Low
Low (CL49)
High (216t)
IV
CL38, CL41, CL46
Yes
Low
Low (CL38)
High (CL41, CL46)
197
HSV-1 (KOS) UL20-null virus, in the presence or absence of either the gBsyn3 or
gKsyn1 mutations, using each mutant UL20 gene revealed that most mutations could be
broadly categorized into four major groups based on their differential effect on infectious
virus production and virus-induced cell fusion (Table 4.3). Group I mutations
complemented infectious virus production and virus-induced cell fusion. These mutations
included: the CL5, CL11, CL16, CL23, CL30 in the amino terminus of UL20p (domain
I); CL121 (domain III); CL173, CL177 (domain IV); CL209 (domain V). Considering
that these cluster mutations involved replacement of multiple amino acids to alanine
residues, these results indicate that UL20p regions spanned by these mutations were not
crucial for the structure and functions of UL20p.
Group II mutations were unable to complement infectious virus production and
virus-induced cell fusion. These mutations included the CL153, 66t, 149t, 181t, 204t, and
211t mutations. The 66t, 149t and 181t mutations specify large carboxyl terminal UL20p
truncations, which are expected to drastically affect the synthesis, transport and membrane
topology of UL20p. Therefore, it is not surprising that these mutations failed to
complement both infectious virus production and virus-induced cell fusion. The 204t and
211t mutations encode carboxyl terminal truncations of 18 and 11 aa, respectively,
encompassing UL20p domain V. Although, these deletions are rather small and not
predicted to affect the membrane incorporation and topology of UL20p (not shown), they
may still affect the structure and function of UL20p. Alternatively, this UL20p carboxyl
terminal domain may be involved in protein-protein interactions necessary for UL20passociated viral functions. The UL20p domain IV is predicted to lie in the lumen or
extracellular side of membranes. Based on the fact that the CL173 and CL177 mutations
198
complemented the UL20-null virus, but the CL153 did not, the data suggests that amino
acids 150-156 are important in infectious virus production. However, none of the single
amino acid changes, E153A, T154A, S156A and D158A significantly altered UL20p’s
ability to complement the UL20-null virus suggesting that the combined effect of the
CL153 mutations may affect the conformation of this domain. Alternatively, this domain
may interact with lumenal portions of other viral glycoproteins, such as gB or gK, to
facilitate infectious virus production and virus-induced cell fusion.
Group III mutations included the CL49 and 216t mutations, located at the amino
and carboxyl termini of UL20p, respectively, which complemented for virus-induced cell
fusion at a reduced level, but failed to efficiently complement infectious virus production.
The carboxyl terminus of UL20p (Domain V) is composed of 18 amino acids and
predicted to localize intracellularly. The 216t complementation results suggest that the
terminal six amino acids of UL20p are crucial for infectious virus production. This result
was further corroborated by the recombinant virus expressing the 216t mutation, which
produced an ultrastructural phenotype similar to the UL20-null virus characterized by the
accumulation of unenveloped capsids into the cytoplasm. The 216t mutation was able to
complement both gBsyn3 and gKsyn1-induced cell fusion, albeit at lower levels than the
wild-type UL20 gene. However, the recombinant virus carrying the 216t mutation in the
∆20syn1 and ∆20syn3 genetic backgrounds revealed no apparent differences in the size
and extent of syncytial plaque formation in comparison to the corresponding ∆20syn1 and
∆20syn3 viruses expressing the wild-type UL20 allele indicating that the UL20 216t
mutation did not adversely affect virus-induced cell fusion.
199
The CL49 mutant phenotype in the amino terminus of UL20p is of particular
interest because alignment of the predicted amino acid sequences of multiple UL20
homologues encoded by alphaherpesviruses revealed conservation of certain amino acid
motifs within the UL20p amino terminus (not shown). One such motif is the YXXΦ
(YSRL) amino acid sequence overlapping the CL49 mutated sequence, which is
conserved in HSV-1, HSV-2, and cercopithecine herpesvirus 1 and 2, but not in varicella
zoster (VZV) or pseurodabies virus (PRV) (not shown). This motif is known to function
in the retrieval of cell-surface expressed proteins to the TGN compartment (Canfield et
al., 1991; Johnson and Kornfeld, 1992; Kirchhausen, Bonifacino, and Riezman, 1997;
Marks et al., 1996; Mellman, 1996; Trowbridge, Collawn, and Hopkins, 1993).
Mutagenesis of the Y residue of a YXXΦ (YTKI) motif within gK domain IV, shown to
lie in the cytoplasmic side of membranes, produced a gK-null phenotype (Foster and
Kousoulas, 1999). Similarly, site directed mutagenesis of the Y49, S50 and R51 residues
within this motif revealed that only the Y49A mutation caused failure to complement the
UL20-null virus indicating that this tyrosine residue played an important role in infectious
virus production. Tyrosine residues within YXXΦ motifs are known to be potentially
phosphorylated. Therefore, it is possible that the Y(49) residue of UL20p is
phosphorylated, and that this phosphorylation may be required for proper intracellular
localization and function of UL20p. This result was further supported by the
ultrastructural phenotype of the recombinant viruses expressing the CL49 and Y49A
mutations, both of which showed an accumulation of unenveloped capsids into the
cytoplasm similar to that of the UL20-null virus. However, the CL49 and the Y49A
mutations complemented, albeit at reduced levels, both gBsyn3 and gKsyn1 induced cell
200
fusion suggesting that these mutations only partially inactivated UL20p functions required
for virus-induced cell fusion. This result was further supported by the small size, but
syncytial plaque phenotype of the recombinant viruses expressing the CL49 mutation in
either the gKsyn1 or gBsyn3 genetic background.
Group IV mutations exhibited a variable effect on virus-induced cell fusion, but
did not drastically reduce infectious virus production. Specifically, the CL38 UL20
mutant gene complemented virus-induced cell fusion caused by the ∆20gBsyn3 and
∆20gKsyn1 viruses at low levels, while the CL41 and CL46 mutant genes complemented
∆20gKsyn1 at low levels but ∆20gBsyn3 at high levels and equivalent to
complementation levels produced with the wild-type UL20 gene. The initial
complementation results were further supported by the recombinant viruses carrying
either the CL41 or CL46 mutation in the gKsyn1 or gBsyn3 genetic background.
Specifically, the gKsyn1 recombinants carrying either the CL41 or CL46 mutations
produced small syncytial plaques, while the gBsyn3 recombinants carrying either the
CL41 or CL46 mutations produced large syncytial virus plaques indistinguishable to the
gBsyn3 recombinant virus carrying the wild-type UL20 allele. These results suggest that
the UL20p amino terminus domain demarcated by the CL38-CL46 mutations is important
in virus-induced cell fusion; however, the specific amino acid requirements within this
domain may be different for gBsyn3 versus gKsyn1-induced cell fusion.
UL20p Mutations that Cause Virus-Induced Cell Fusion
Characteristically, recombinant viruses expressing the CL49, Y49A, CL209,
R209A and 216t UL20p mutations in a wild-type genetic background formed small
syncytial plaques on Vero cells. Recombinant viruses specifying the T212A and R213A
201
mutations did not produce syncytial plaques on Vero cells indicating that the R209A
mutation was responsible for the syncytial phenotype of the CL209 mutation. Thus, it is
evident that the amino and carboxyl termini of UL20p are directly associated with virusinduced cell fusion in the context of wild-type gB or gK expression. Therefore, the ability
of these mutations to complement either gBsyn3 or gKsyn1 virus-induced cell fusion
discussed earlier may be in part due to the inherent fusogenic character of the mutated
UL20p. Most likely, this is not the case because recombinant viruses carrying the UL20
R209A syncytial mutation as well as either the gBsyn3 or gKsyn1 mutation did not
exhibit increased virus-induced cell fusion in comparison to viruses specifying either the
gBsyn3 or gKsyn1 mutations and the wild-type UL20 gene (not shown). Alternatively,
UL20p may function as a regulator of virus-induced cell fusion through interactions with
other viral proteins such as gB and gK and disruption of these interactions by specific
UL20p mutations may result in syncytia formation.
Deletion of the gK gene or lethal gK mutations cause entrapment of enveloped
virions within cytoplasmic vesicles presumably originating from the TGN compartment
and the accumulation of unenveloped capsids in the cytoplasm (Foster and Kousoulas,
1999; Jayachandra, Baghian, and Kousoulas, 1997). Similarly, deletion of the UL20 gene,
or lethal UL20p mutations cause accumulation of unenveloped capsids into the cytoplasm
and a small portion of enveloped capsids within cytoplasmic vesicles (Foster et al., 2004;
Fuchs et al., 1997). These results suggest that UL20p and gK may have interrelated
functions with regard to cytoplasmic virion envelopment. The fact that the Y49A, CL49
and 216t mutant viruses produced syncytial plaques, although their ultrastructural
phenotypes seemed to be similar to that of the UL20-null virus suggests that UL20
202
functions for virus-induced cell fusion may be distinct from those functioning in
cytoplasmic virion envelopment. Alternatively, the same domains may function in both
cytoplasmic virion envelopment and virus-induced cell fusion; however, virion
envelopment may be more susceptible to the negative effects of these mutations in
comparison to virus-induced cell fusion.
Overall, our previous findings and the results presented in this paper suggest the
existence of functional interactions between gK and UL20p as well as between UL20p
and gB. In this regard, it is interesting to note that syncytial mutations in gB lie in the
intracellular carboxyl terminus of gB allowing the possibility that intracellular domains of
UL20p (domains I and V) directly or indirectly interact with the carboxyl terminus of gB
altering gB’s fusogenic properties.
REFERENCES
Aiyar, A., Xiang, Y., and Leis, J. (1996). Site-directed mutagenesis using overlap
extension PCR. Methods Mol Biol 57, 177-91.
Alconada, A., Bauer, U., and Hoflack, B. (1996). A tyrosine-based motif and a casein
kinase II phosphorylation site regulate the intracellular trafficking of the varicellazoster virus glycoprotein I, a protein localized in the trans-Golgi network. Embo J
15(22), 6096-110.
Alconada, A., Bauer, U., Sodeik, B., and Hoflack, B. (1999). Intracellular traffic of herpes
simplex virus glycoprotein gE: characterization of the sorting signals required for
its trans-Golgi network localization. J Virol 73(1), 377-87.
Avitabile, E., Di, G. S., Torrisi, M. R., Ward, P. L., Roizman, B., and Campadelli-Fiume,
G. (1995). Redistribution of microtubules and Golgi apparatus in herpes simplex
virus-infected cells and their role in viral exocytosis. J Virol 69(12), 7472-82.
Avitabile, E., Ward, P. L., Di Lazzaro, C., Torrisi, M. R., Roizman, B., and CampadelliFiume, G. (1994). The herpes simplex virus UL20 protein compensates for the
differential disruption of exocytosis of virions and viral membrane glycoproteins
associated with fragmentation of the Golgi apparatus. J Virol 68(11), 7397-405.
203
Baines, J. D., Ward, P. L., Campadelli-Fiume, G., and Roizman, B. (1991). The UL20
gene of herpes simplex virus 1 encodes a function necessary for viral egress. J
Virol 65(12), 6414-24.
Bjerke, S. L., Cowan, J. M., Kerr, J. K., Reynolds, A. E., Baines, J. D., and Roller, R. J.
(2003). Effects of charged cluster mutations on the function of herpes simplex
virus type 1 UL34 protein. J Virol 77(13), 7601-10.
Bond, V. C., and Person, S. (1984). Fine structure physical map locations of alterations
that affect cell fusion in herpes simplex virus type 1. Virology 132(2), 368-76.
Brideau, A. D., del Rio, T., Wolffe, E. J., and Enquist, L. W. (1999). Intracellular
trafficking and localization of the pseudorabies virus Us9 type II envelope protein
to host and viral membranes. J Virol 73(5), 4372-84.
Bzik, D. J., Fox, B. A., DeLuca, N. A., and Person, S. (1984). Nucleotide sequence of a
region of the herpes simplex virus type 1 gB glycoprotein gene: mutations
affecting rate of virus entry and cell fusion. Virology 137(1), 185-90.
Canfield, W. M., Johnson, K. F., Ye, R. D., Gregory, W., and Kornfeld, S. (1991).
Localization of the signal for rapid internalization of the bovine cationindependent mannose 6-phosphate/insulin-like growth factor-II receptor to amino
acids 24-29 of the cytoplasmic tail. J Biol Chem 266(9), 5682-8.
Debroy, C., Pederson, N., and Person, S. (1985). Nucleotide sequence of a herpes simplex
virus type 1 gene that causes cell fusion. Virology 145(1), 36-48.
Desai, P., DeLuca, N. A., Glorioso, J. C., and Person, S. (1993). Mutations in herpes
simplex virus type 1 genes encoding VP5 and VP23 abrogate capsid formation and
cleavage of replicated DNA. J Virol 67(3), 1357-64.
Dietz, P., Klupp, B. G., Fuchs, W., Kollner, B., Weiland, E., and Mettenleiter, T. C.
(2000). Pseudorabies virus glycoprotein K requires the UL20 gene product for
processing. J Virol 74(11), 5083-90.
Foster, T. P., Alvarez, X., and Kousoulas, K. G. (2003). Plasma membrane topology of
syncytial domains of herpes simplex virus type 1 glycoprotein K (gK): the UL20
protein enables cell surface localization of gK but not gK-mediated cell-to-cell
fusion. J Virol 77(1), 499-510.
Foster, T. P., and Kousoulas, K. G. (1999). Genetic analysis of the role of herpes simplex
virus type 1 glycoprotein K in infectious virus production and egress. J Virol
73(10), 8457-68.
Foster, T. P., Melancon, J. M., Baines, J. D., and Kousoulas, K. G. (2004). Herpes simples
virus type 1 (HSV-1) UL20 protein modulates membrane fusion events during
cytoplasmic virion morphogenesis and virus-induced cell fusion. J Virol
78(10):5347-57.
204
Fuchs, W., Klupp, B. G., Granzow, H., and Mettenleiter, T. C. (1997). The UL20 gene
product of pseudorabies virus functions in virus egress. J Virol 71(7), 5639-46.
Hirokawa, T., Boon-Chieng, S., and Mitaku, S. (1998). SOSUI: classification and
secondary structure prediction system for membrane proteins. Bionformatics 14,
378-379.
Hofmann, K., and Stoffel, W. (1993). TMbase-database of membrane spanning segments.
J. Biol. Chem. 377, 166.
Hutchinson, L., Goldsmith, K., Snoddy, D., Ghosh, H., Graham, F. L., and Johnson, D. C.
(1992). Identification and characterization of a novel herpes simplex virus
glycoprotein, gK, involved in cell fusion. J Virol 66(9), 5603-9.
Hutchinson, L., and Johnson, D. C. (1995). Herpes simplex virus glycoprotein K promotes
egress of virus particles. J Virol 69(9), 5401-13.
Jacobson, J. G., Chen, S. H., Cook, W. J., Kramer, M. F., and Coen, D. M. (1998).
Importance of the herpes simplex virus UL24 gene for productive ganglionic
infection in mice. Virology 242(1), 161-9.
Jayachandra, S., Baghian, A., and Kousoulas, K. G. (1997). Herpes simplex virus type 1
glycoprotein K is not essential for infectious virus production in actively
replicating cells but is required for efficient envelopment and translocation of
infectious virions from the cytoplasm to the extracellular space. J Virol 71(7),
5012-24.
Johnson, D. C., and Huber, M. T. (2002). Directed egress of animal viruses promotes cellto-cell spread. J Virol 76(1), 1-8.
Johnson, K. F., and Kornfeld, S. (1992). The cytoplasmic tail of the mannose 6phosphate/insulin-like growth factor-II receptor has two signals for lysosomal
enzyme sorting in the Golgi. J Cell Biol 119(2), 249-57.
Kirchhausen, T., Bonifacino, J. S., and Riezman, H. (1997). Linking cargo to vesicle
formation: receptor tail interactions with coat proteins. Curr Opin Cell Biol 9(4),
488-95.
MacLean, C. A., Efstathiou, S., Elliott, M. L., Jamieson, F. E., and McGeoch, D. J.
(1991). Investigation of herpes simplex virus type 1 genes encoding multiply
inserted membrane proteins. J Gen Virol 72(Pt 4), 897-906.
Marks, M. S., Woodruff, L., Ohno, H., and Bonifacino, J. S. (1996). Protein targeting by
tyrosine- and di-leucine-based signals: evidence for distinct saturable components.
J Cell Biol 135(2), 341-54.
Mellman, I. (1996). Endocytosis and molecular sorting. Annu Rev Cell Dev Biol 12, 575625.
205
Mettenleiter, T. C. (2002). Herpesvirus assembly and egress. J Virol 76(4), 1537-47.
Pellett, P. E., Kousoulas, K. G., Pereira, L., and Roizman, B. (1985). Anatomy of the
herpes simplex virus 1 strain F glycoprotein B gene: primary sequence and
predicted protein structure of the wild type and of monoclonal antibody-resistant
mutants. J Virol 53(1), 243-53.
Pogue-Geile, K. L., Lee, G. T., Shapira, S. K., and Spear, P. G. (1984). Fine mapping of
mutations in the fusion-inducing MP strain of herpes simplex virus type 1.
Virology 136(1), 100-9.
Roizman, B., and Sears, A. E. (1996). Herpes Simplex Viruses and Their Replication.
Third edition ed. In "Feilds Virology" (B. N. Feilds, D. M. Knipe, and P. M.
Howley, Eds.), Vol. 2, pp. 2231-2295. 1 vols. Lippincott-Raven Publishers,
Philadelphia, PA.
Ryechan, W. T., Morse, L. S., Knipe, D. M., and Roizman, B. (1979). Molecular genetics
of herpes simplex virus. II. Mapping of the major viral glycoproteins and of the
genetic loci specifying the social behavior of infected cells. J Virol. 29, 677697.
Sanders, P. G., Wilkie, N. M., and Davison, A. J. (1982). Thymidine kinase deletion
mutants of herpes simplex virus type 1. J
Gen Virol 63(2), 277-95.
Spear, P. G. (1993a). Entry of alphaherpesviruses into cells. Seminars in Virology 4, 167180.
Spear, P. G. (1993b). Membrane fusion induced by herpes simplex virus. In "Viral fusion
mechanisms" (J. Bentz, Ed.), pp. 201-232. CRC Press., Boca Raton, Fla.
Spear, P. G., Eisenberg, R. J., and Cohen, G. H. (2000). Three classes of cell surface
receptors for alphaherpesvirus entry. Virology 275(1), 1-8.
Tirabassi, R. S., and Enquist, L. W. (1999). Mutation of the YXXL endocytosis motif in
the cytoplasmic tail of pseudorabies virus gE. J Virol 73(4), 2717-28.
Tomishima, M. J., Smith, G. A., and Enquist, L. W. (2001). Sorting and transport of alpha
herpesviruses in axons. Traffic 2(7), 429-36.
Trowbridge, I. S., Collawn, J. F., and Hopkins, C. R. (1993). Signal-dependent membrane
protein trafficking in the endocytic pathway. Annual Reviews of Cell Biology 9,
129-61.
Wan, L., Molloy, S. S., Thomas, L., Liu, G., Xiang, Y., Rybak, S. L., and Thomas, G.
(1998). PACS-1 defines a novel gene family of cytosolic sorting proteins required
for trans-Golgi network localization. Cell 94(2), 205-16.
206
Ward, P. L., Campadelli-Fiume, G., Avitabile, E., and Roizman, B. (1994). Localization
and putative function of the UL20 membrane protein in cells infected with herpes
simplex virus 1. J Virol 68(11), 7406-17.
Zhu, Z., Hao, Y., Gershon, M. D., Ambron, R. T., and Gershon, A. A. (1996). Targeting
of glycoprotein I (gE) of varicella-zoster virus to the trans-Golgi network by an
AYRV sequence and an acidic amino acid-rich patch in the cytosolic domain of
the molecule. J Virol 70(10), 6563-75.
207
CHAPTER V
CONCLUDING REMARKS
SUMMARY
The Herpes Simplex Virus life cycle contains a number of membrane fusion
events that must function properly to ensure a productive infection, including: virus
attachment and entry into susceptible cells, de-envelopment at the outer nuclear
membrane, and virus-induced cell-to-cell fusion. Virus-induced cell-to-cell fusion is
mediated by viral glycoproteins expressed on cellular membranes and has been
extensively studied for several reasons: as a probe of the structure and function of
cellular membranes during infection reflected in their behavior, as a tool for analysis of
the functions of viral membrane proteins, and as a model of the initial interaction
between HSV and susceptible cells that results in fusion of the viral envelope with the
cellular plasma membrane (Campadelli-Fiume and Serafini-Cessi, 1985; Roizman, 1962;
Spear, 1985; Spear, 1993).
A virus-free cell fusion assay was recently developed in order to attempt to
understand the underlying mechanisms that are responsible for viral fusion events. In this
system, transiently expressed gB, gD, gH, and gL are necessary and sufficient to induce
syncytia formation in certain cell types (Turner et al., 1998). The transient fusion assay
was utilized in Chapter II to investigate the effect of mutations targeted to the carboxyl
terminus of gB. First, it was demonstrated that the predicted α−helical domain H17b
within the carboxyl-terminus of gB is involved in both virus-induced and virus-free
fusion systems. Second, heparin was shown to be a specific inhibitor of gB-mediated
208
fusion in both systems. Third, resistance to heparin inhibition of gB-mediated cell fusion
was associated with the predicted α−helical structure H17b. Thus, gB-mediated fusion is
functionally similar for both fusion assays. The observed enhancement of cell fusion
resulting from the disruption of H17b raises the question of how H17b regulates
membrane fusion. In this regard, it is possible that during a productive infection other
viral proteins interact with this region in the carboxyl terminus of gB to inhibit aberrant
membrane fusion from occurring. Alteration of the conformation of the gB carboxyl
terminus may cause an activation of the fusogenic potential of gB, disengaging gB from
other viral or cellular controls and resulting in unregulated membrane fusion.
The sole requirement for gB, gD, gH, and gL in the transient membrane fusion
system is similar to the requirement for virus entry. An important difference between the
virus-free and virus-induced membrane fusion systems is that virus-expressed gBmediated low amounts of cell-to-cell fusion, while transient expression of gB causes
significant amounts of cell-to-cell fusion. This observation has prompted the hypothesis
that additional viral proteins exert negative control on gB-mediated cell fusion, with the
primary candidates being viral proteins and glycoproteins such as UL20p and gK that
themselves contain syncytial mutations. In this regard, syncytial mutations in gK and
UL20p could be thought to inactivate the negative regulatory control of these molecules
on gB function, thus releasing the natural fusogenic properties of gB. In support of this
hypothesis, gK was shown to inhibit cell fusion resulting from transiently expressed gB,
gD, gH and gL, while a syncytial gK variant showed a decreased ability to inhibit cell
fusion (Avitabile, Lombardi, and Campadelli-Fiume, 2003). In addition, it was
209
previously shown that gB is absolutely required for virus-induced cell fusion to occur
resulting from syncytial mutations in UL53 (gK) (Cai, Gu, and Person, 1988).
However, in herpesviruses, complexity seems to prevail over simplicity; and we
showed in Chapter III that gK can not solely act as a negative regulator of gB-mediated
membrane fusion, since gK is absolutely required for virus-induced cell fusion to occur
resulting from the gBsyn3 syncytial mutation. Moreover, the gBamb1511 mutation that
disrupts H17b could not cause virus-induced cell fusion in the absence of gK. These
findings definitively abolish the hypothesis that gK is an inhibitor of virus-induced cell
fusion and suggests a more complicated relationship between gK and gB. Interestingly, a
similar result was reported for gM, which inhibited membrane fusion in the transient
coexpression system (Klupp, Nixdorf, and Mettenleiter, 2000; Koyano et al., 2003), but
was required for virus-induced cell fusion caused by syncytial mutations in gB (DavisPoynter et al., 1994). In addition, gE, gI, and UL20 were all shown to be required for the
virus-induced fusion of cellular membranes (Davis-Poynter et al., 1994; Foster et al.,
2004a), while these proteins were not required for fusion to occur in the transient system.
Consequently, these results indicate that regulation and activation of the herpesvirus
membrane fusion machinery during virus-induced cell fusion is mediated by a complex
of many different viral proteins. Alternatively, additional functions of other glycoproteins
may be required prior to gB-mediated membrane fusion. These functions may involve
tighter binding to additional cellular co-receptors on opposed membranes that may
render a more favorable environment for enabling gB’s fusogenic properties.
HSV-1 is thought to acquire an initial envelope when capsids formed in the
nucleus of cells bud into the perinuclear space. Subsequently, a putative fusion event
210
between viral envelopes and the outer nuclear membrane, termed de-envelopment, results
in capsid release into the cytoplasm. Final envelopment is thought to occur when
cytoplasmic capsids bud into vesicles derived from the TGN or endosomes that facilitate
egress of enveloped virions to extracellular spaces. Previously, it was not known whether
viral glycoproteins involved in virus entry or virus-induced cell fusion also function in
the de-envelopment step. The results presented in Chapter III clearly show that gK, UL20
and gB are not essential for virion de-envelopment. In the case of gB, previous studies
suggested that lack of gB did not adversely influence egress of virion particles to
supernates of infected cells, although these virions lacked gB and were rendered noninfectious (Cai et al., 1987; Cai, Gu, and Person, 1988). Ultrastructural examination of
the gB-null virus confirmed initial findings and clearly showed that gB is not required for
efficient production of enveloped virions in extracellular spaces. Examination of
recombinant viruses specifying single gene deletions in either gB, gK or UL20p did not
exclude the possibility that gB and gK could function in a redundant manner to facilitate
virion de-envelopment. This possibility can be discounted on the basis that both the
gB/gK double null virus and the gB/UL20 double null virus efficiently produced
unenveloped capsids into the cytoplasm of infected cells. However, the possibility still
exists that gH/gL could mediate de-envelopment in the absence of either gB and gK or
gB and UL20.
Recently, we showed that a precise deletion of the UL20 gene caused
accumulation of unenveloped capsids into the cytoplasm indicating that the HSV-1
UL20p functioned in cytoplasmic stages of virion envelopment and that the previously
reported partial deletion of the UL20 gene caused a predicted fusion of the UL20p to the
211
adjacent UL20.5 gene. In addition, syncytial mutations in either gB or gK failed to cause
fusion in the absence of the UL20 gene, suggesting that the UL20 protein was essential
for virus-induced cell fusion (Foster et al., 2004a). In Chapter IV, we delineated via sitedirected mutagenesis the functional domains of UL20p involved in infectious virus
production and virus-induced cell fusion. The data suggest that UL20p’s role in virusinduced cell fusion can be functionally separated from its role in cytoplasmic virion
morphogenesis, and that UL20p domains that function in gB-syn3 virus-induced cell
fusion are distinct from those functioning in gKsyn1 virus-induced cell fusion.
As a result of the research presented here, several hypotheses concerning the
herpesvirus fusion machinery were successfully addressed. We found that syncytial
mutations in gB can act differently in a virus-induced versus a virus-free fusion system,
while identifying a region of gB involved in the regulation of membrane fusion. In
addition, we found that gK is required for virus-induced cell fusion to occur, despite its
apparent inhibitory effect in virus-free systems. We eliminated the proposed hypothesis
of redundant functions for gB and gK in the fusion event occurring during virion deenvelopment. Furthermore, we identified the specific regions of UL20 required for virion
egress and virus-induced cell fusion and demonstrated separation of the two functions.
The results presented here represent significant progress into the understanding of
herpesvirus membrane fusion, but there is still a great deal of knowledge that must be
obtained to fully decipher the mechanisms involved.
CURRENT AND FUTURE RESEARCH CHALLENGES
The work completed on mutagenesis of the UL20 protein has left several open
questions regarding the functional interrelationship between gK and UL20p. Recently,
212
our laboratory showed that UL20 and gK are interdependent for transport out of the
endoplasmic reticulum, transport to the plasma membrane, and internalization to the
trans-Golgi network (Foster et al., 2004b); however, the domains of either protein
required for proper localization have not been examined. The data presented in Chapter
IV revealed several mutant forms of UL20p that showed defects in function for virusinduced cell fusion and/or final envelopment. As a result, we sought to identify whether
or not the deficiencies could be attributed to a malfunction in protein transport and
localization. If a mutant UL20p was unable to transport gK, this would indicate that the
mutation potentially affects either an interacting domain or the ability of the UL20
protein to fold correctly; whereas if the mutant UL20p transports gK correctly, then the
mutation would be thought to affect a UL20 domain involved in interactions with other
viral or cellular components critical for proper function.
To investigate the ability of each UL20p mutant to transport gK, Vero cells were
co-transfected with a plasmid coding for gK as well as with plasmids encoding wild-type
or mutant UL20p. The transport and localization of both UL20p and gK was determined
by confocal microscopy. An αTGN46 pAb was used to determine colocalization with
the TGN. As expected, wt UL20 and 216t, which complemented the UL20-null fusion
defect, are able to mediate gK and UL20 colocalization at the TGN (Fig. 5.1: A, F).
CL153 and 204t, which were not able to complement the UL20-null defect, are not able
to mediate gK translocation from the endoplasmic reticulum (ER), and both retain an ER
like distribution (Fig. 5.1: C, D). In contrast, both CL61 and 211t, which were both
unable to complement the UL20-null defect, are able to mediate colocalization with gK at
the TGN (Fig. 5.1: B, E).
213
Figure 5.1: gK localization at the TGN mediated by selected UL20p mutants. Vero
cells were co-transfected with gKD1V5 as well as with plasmids encoding wild-type or
mutant UL20p. 36 hours post-transfection, cells were washed thoroughly, fixed, and
processed for confocal microscopy. After permeabilization, rabbit αFLAG pAb was used
to identify UL20, and sheep αTGN46 pAb was used to identify the TGN.
214
215
Figure 5.2: gK cell surface expression and endocytosis to the TGN mediated by
selected UL20p mutants. Vero cells were co-transfected with gKD1V5 as well as with
plasmids encoding wild-type or mutant UL20p. 24 hours post-transfection, cells were
incubated under live conditions with aV5 (gK) mAb for 6 hours. Cells were washed
thoroughly, fixed, and processed for confocal microscopy. After permeabilization, rabbit
αFLAG pAb was used to identify UL20, and sheep αTGN46 pAb was used to identify
the TGN.
216
217
An internalization assay was also performed to determine whether gK was
expressed on cell surfaces and subsequently internalized to the TGN. Results obtained
were similar as those in the colocalization experiments. As expected, wt UL20 and 216t
are able to mediate gK surface expression and subsequent endocytosis to the TGN (Fig.
5.2: A, F). CL153 and 204t are not able to mediate gK internalization, and the mutant
UL20 retains an endoplasmic reticulum (ER)-like distribution (Fig. 5.2: C, D). Again,
both CL61 and 211t, which were both unable to complement the UL20-null defect, are
able to mediate gK surface expression and internalization to the TGN (Fig. 5.2: B, E).
The CL61 mutation is located adjacent to a predicted transmembrane region and may
affect the correct folding of domains that are not required for proper localization.
Taken together, these results indicate that UL20-mediated transport of gK to the
cell surface and subsequent internalization ot the TGN is not sufficient for UL20/gK to
function properly in virion egress or virus induced cell-to-cell fusion. Specific domains of
UL20 must be involved in interactions with viral or cellular proteins. The differing
results of the carboxyl-terminal truncations of UL20 indicate that further work must be
done to understand the function of the carboxyl terminus of UL20 in virion egress and
virus-induced cell fusion.
In the amino terminus of UL20, the mutations CL38 and CL49 previously showed
interesting but opposing phenotypes, whereby CL38 was virtually non-functional in cell
fusion and CL49 showed a severe defect in virion egress (see Chapter IV). Interestingly,
both CL38 and CL49 are able to mediate gK localization at the TGN (data not shown).
Our hypothesis was that a double mutation containing both CL38 and CL49 should
eliminate both aspects of UL20 function, and the ability of the CL38-49 mutant to
218
transport gK would be tested as well to evaluate the nature of the defect in function.
Preliminary complementation experiments indicate that the CL38-49 mutation does
indeed eliminate the function of UL20 in both virus-induced cell fusion and infectious
virus production. In addition, recombinant viruses containing the CL38-49 mutation
were not readily isolated on non-complementing cells, indicating that the mutation is
probably lethal to virus infectivity. Analysis of CL38-49 and gK transport revealed that
there is no apparent defect in cellular localization (data not shown). As a result, further
analysis must be done to characterize the nature of the CL38-49 defect. Hopefully,
deciphering the basis of function of the tyrosine residues at positions 38 and 49 in UL20
will provide a clearer picture of the role that UL20 plays in the virus life cycle.
The UL20 gene encodes a protein of 222 amino acids with a predicted mass of
24kDa. However, as depicted below, UL20p is visualized as a 30-32 kDa protein
consisting of multiple protein species (Fig. 5.3: A). Cluster-to-alanine mutagenesis
revealed that mutations within the UL20p amino terminus altered the apparent molecular
mass of UL20p-related proteins, indicating that the UL20p amino terminus is potentially
post-translationally modified (data not shown). The amino terminus of UL20p contains
multiple potential phosphorylation sites as predicted by NetPhos 2.0 (Blom, Gammeltoft,
and Brunak, 1999). Interestingly, the tyrosine residues at position 38 and 49 have the
potential for phosphorylation. Current and future experiments are targeted to determine
the nature of these UL20p modifications.
In order to ascertain whether or not UL20p is phosphorylated, infections of UL20null or wild type virus were performed with the presence of 32P (orthophosphate) in
phospahate-free media at late times post infection. A portion of extracted protein was
219
Figure 5.3: Western analysis and potential phosphorylation of UL20p. (A) Vero
cells were either untransfected or transfected with a plasmid specifying UL20p with an
amino terminal 3xFLAG epitope tag prior to infection with a UL20-null virus. Western
analysis of UL20p was performed utilizing a FLAG mAb. (B, C) Vero cells were infected
with either a UL20-null or a wild-type virus that encodes UL20p with an amino terminal
3xFLAG epitope tag at an MOI of 10. 11 hours post infection, the media was replaced
and cells were incubated with phosphate-free media for 1 hour. Phosphate-free media
containing 32P (orthophosphate, 250 µCi/mL) was then added for 6 hours before cell
lysates were prepared and immunoprecipitation of UL20 was performed. The 32P profiles
of the total lysate (B) and the UL20 immunoprecipitations (C) are shown above.
Asterisks are provided to show corresponding bands representing UL20p.
220
used for immunoprecipitation of UL20, and both the total cellular extracts and the
immunoprecipitated UL20 were analyzed after SDS-PAGE utilizing the Perkin Elmer
Cyclone Storage Phosphor Imaging System (Fig 5.3: B, C). While the 32P total viral
protein profile did not reveal any significant differences between the UL20-null and the
wild-type virus, the 32P profile resulting from the UL20 immunoprecipitation clearly
shows two bands present in only the wild-type sample around 30 kDa, which seem to
correspond with the profile of UL20 seen through western blot analysis (Fig 5.3). A
group of higher molecular weight bands are also present between 60 and 90 kDa that may
or may not be non specific background. Further experiments must be performed to
confirm the presence of phosphorylation on UL20p. One approach to identifying the
location of UL20p phosphorylation would be to screen the library of mutant UL20 genes
that delete consensus phosphorylation sites. The confirmation of phosphorylation on
UL20p would imply that UL20 functions, including those during virion egress and virusinduced cell fusion, could be regulated by cellular or viral kinases, whereby only one
form of UL20p could properly function in gK transport to cell surfaces or final
envelopment at the TGN. Alternatively, UL20p phosphorylation could be required for
interactions with gB that serve to regulate virus-induced cell fusion.
While genetic analysis has been successful in revealing the components involved
in fusion processes, one of the biggest obstacles to truly understanding membrane fusion
in herpesviruses is the complete lack of knowledge regarding the mechanism that triggers
fusion between opposing membranes. It would be of great interest to determine whether
or not there is a conformational change in gB that results from the regulated activation of
the membrane fusion machinery following the binding of gD to an entry receptor. The
221
syncytial forms of gB provide a unique opportunity to study conformation changes in the
ectodomain of gB, as syncytial mutations in gB reside almost exclusively in the
cytoplasmic carboxyl terminus. Therefore, changes from syncytial mutations resulting in
a loss of regulation of the membrane fusion machinery could be thought to occur through
“activation” of gB due to a conformational change in its extracellular N-terminus. In
Chapter II we found that heparin inhibition of membrane fusion could be circumvented
through generation of specific mutations in the predicted alpha helical region, H17b, in
the carboxyl terminus of gB. Since the heparin binding sites of gB reside exclusively in
its ectodomain, it is reasonable to suggest that resistance to heparin inhibition resulted
from an alteration of gB’s conformation causing inability of gB to bind to heparin.
Consequently, the ability of gB to bind to labeled soluble heparin could be used to map
the potential activation of gB due to an altered conformation. By eliminating potential
heparin binding sites in the extracellular portion of gB through site directed mutagenesis,
it would also be possible to study gB and its syncytial forms by comparison of the regions
that are required for heparin inhibition of cell fusion with the regions that are modified
due to syncytial mutations in H17b.
Since UL20p and gK are required for gB-mediated virus-induced cell fusion to
occur, it would also be very important to examine whether or not the conformation of gB
is altered in the presence or absence of UL20p and/or gK, implying a direct functional
interaction between gB and UL20p or gK. A similar method to that described above
could be utilized to study the effect of additional viral proteins on the conformation of gB
through its ability to bind to soluble heparin. Such an understanding of the role of gB in
membrane fusion could provide leads into the way that gB, gD, and gH/gL interact with
222
gK, UL20p and other viral or cellular proteins to regulate the potential of the herpesvirus
membrane fusion machinery.
Perhaps the least understood aspect of the herpesvirus life cycle is virion
morphogenesis and egress. After mature capsids envelope and de-envelope at the
perinuclear space, they transit through an unknown mechanism to cytoplasmic vesicles
where they obtain a final envelope. The mechanism and components required for final
envelopment are not clear; however, genetic analysis has identified gK and UL20p as
potential candidates to mediate the final envelopment step. Deletion of either UL20p or
gK results in a cytoplasmic block in virion egress consisting of an accumulation of
unenveloped and partially enveloped capsids in the cytoplasm of infected cells. Through
extensive targeted mutagenesis of UL20p, we sought to identify domains that were
functional in protein localization and cell fusion, but not virion egress. The available
evidence seems to indicate that the carboxyl terminus of UL20 may interact with viral
tegument proteins to assist in the final envelopment step. Several candidate tegument
proteins have been identified through careful examination of their ultrastructural
phenotypes. One approach to identifying components of the egress pathway is to generate
multiple deletions of viral tegument and membrane proteins. We are currently generating
double-null viruses containing deletions in UL20 and specific tegument proteins in an
attempt to reveal redundant or sequential steps in the virion egress pathway.
FINAL COMMENTS
Membrane fusion events occur regularly as necessary biological processes in the
cell. The fusion process itself is as an energetically unfavorable event necessitating the
use of fusogenic proteins acting alone or as part of a complex to assist in overcoming the
223
forces that discourage fusion from occurring (Rand et al., 1988). Endocytosis,
exocytosis, and other intracellular membrane trafficking events are all mediated by fusion
processes that occur continuously in the cell. It is known that membrane fusion of
transport vesicles to targeted intracellular membranes are carried out by complex “fusion
machines” that consist of a core of integral proteins and also a number of peripheral
regulatory proteins.
All enveloped viruses utilize membrane fusion between their viral envelopes and
cellular membranes to facilitate entry into the cytoplasm of cells. Most enveloped viruses
make use of a single fusogenic protein to mediate entry, and that fusion protein is also
necessary and sufficient to confer fusion activity when expressed in tissue culture cells.
Such is the case for the influenza virus hemagglutinin (HA) (Wiley and Skehel, 1987),
human immunodeficiency virus type 1 (HIV-1) gp41 (Lifson et al., 1986; Sodroski et al.,
1986), Ebola virus GP2 (Takada et al., 1997), vesicular stomatitis virus glycoprotein
(VSV-G) (Florkiewicz and Rose, 1984), and the coronavirus Spike protein (corona)
(Xiao et al., 2003). Viral fusion proteins often combine into homotrimers that project
from the viral envelope. Fusion proteins are usually are synthesized in an inactive form,
and a conformational activation is required before optimal fusion activity is acquired
under physiological conditions.
The Herpes Simplex Virus life cycle contains a number of membrane fusion
events, including: virus attachment and entry into susceptible cells, de-envelopment at
the outer nuclear membrane, and virus-induced cell-to-cell fusion. In contrast to other
enveloped viruses, the entry process of HSV-1 requires the presence of at least four
glycoproteins: gB, gD, gH and gL. These four membrane proteins are also necessary and
224
sufficient to mediate cell-to-cell fusion when expressed in tissue culture cells. The
mutations that cause extensive virus-induced cell-to-cell fusion in HSV-1 map
predominantly to the genes that encode gB and gK. The differential effects of UL20p
mutations on the regulation of gB- and gK- mediated syncytia formation imply that
functional intreractions occur between UL20p, gB, and gK. The requirement for gK and
UL20p in gB-induced syncytia formation provides further evidence for a functional
protein complex. In addition, preliminary results from our laboratory indicate that
UL20p directly interacts with both gK and gB (personal communication, Dr. Timothy
Foster). Based on these results, we have proposed that the presence of a core complex of
proteins involved in regulation of HSV-1 virus-induced cell fusion that we have termed
the “Alphaherpesvirus Fuseosome” consisting of UL20p, gK, gB, gD, and the gH/gL
heterodimer (Fig. 5.4).
It is important to note that the proposed complex appears to function only at cell
surfaces during virus-induced cell fusion. As discussed earlier, gB is not essential for
either de-envelopment or cytoplasmic envelopment during virion egress. In this regard,
the UL20p/gK complex must carry out fundamentally distinct functions during
cytoplasmic envelopment relative to those carried out at cell surfaces. One potential
explanation for the distinct functions of UL20p/gK may be differential processing of
UL20p via phosphorylation or other means. In this regard, it is known that viral proteins
that are structural components of the virion are predominantly underphosphorylated
relative to infected cells. Thus, it is possible that gK/UL20p underphosphorylation
promotes interactions required for cytoplasmic envelopment, while phosphorylation of
UL20p promotes interactions of the UL20p/gK complex with gB at cell surfaces.
225
gB
UL20
gH/gL
gK
gD
?
Figure 5.4: The proposed Alphaherpesvirus Fuseosome. A schematic representation
of interactions between the core integral membrane proteins responsible for regulation of
HSV-1 virus-induced cell-to-cell fusion is shown. UL20p is thought to interact directly
with both gB and gK (blue arrows). There are also thought to be functional interactions
that must take place between gB and gD or gH/gL (green arrow) in order to activate the
fusogenic potential of gB.
226
REFERENCES
Avitabile, E., Lombardi, G., and Campadelli-Fiume, G. (2003). Herpes simplex virus
glycoprotein K, but not its syncytial allele, inhibits cell-cell fusion mediated by
the four fusogenic glycoproteins, gD, gB, gH, and gL. J Virol 77(12), 6836-44.
Blom, N., Gammeltoft, S., and Brunak, S. (1999). Sequence and structure-based
prediction of eukaryotic protein phosphorylation sites. In "J Mol Biol", Vol. 294,
pp. 1351-62.
Cai, W., Person, S., Warner, S. C., Zhou, J., and DeLuca, N. A. (1987). Linker-insertion
nonsense and restriction-site deletion mutations of the gB glycoprotein gene of
herpes simplex virus type 1. Journal of Virology 61, 714-721.
Cai, W. H., Gu, B., and Person, S. (1988). Role of glycoprotein B of herpes simplex virus
type 1 in viral entry and cell fusion. Journal of Virology 62(8), 2596-604.
Campadelli-Fiume, G., and Serafini-Cessi, F. (1985). Processing of the oligosaccharide
chains of herpes simplex virus type 1 glycoproteins. In "The herpesviruses" (B.
Roizman, Ed.), Vol. 3, pp. 357-382. Plenum, New York.
Davis-Poynter, N., Bell, S., Minson, T., and Browne, H. (1994). Analysis of the
contributions of herpes simplex virus type 1 membrane proteins to the induction
of cell-cell fusion. Journal of Virology 68(11), 7586-90.
Florkiewicz, R. Z., and Rose, J. K. (1984). A cell line expressing vesicular stomatitis
virus glycoprotein fuses at low pH. Science 225(4663), 721-3.
Foster, T. P., Melancon, J. M., Baines, J. D., and Kousoulas, K. G. (2004a). The Herpes
Simplex Virus Type 1 UL20 Protein Modulates Membrane Fusion Events during
Cytoplasmic Virion Morphogenesis and Virus-Induced Cell Fusion. J Virol
78(10), 5347-57.
Foster, T. P., Melancon, J. M., Olivier, T. L., and Kousoulas, K. G. (2004b). Herpes
simplex virus type-1 (HSV-1) glycoprotein K (gK) and the UL20 protein are
interdependent for intracellular trafficking and trans-Golgi network localization. J
Virol, (In Press).
Klupp, B. G., Nixdorf, R., and Mettenleiter, T. C. (2000). Pseudorabies virus
glycoprotein M inhibits membrane fusion. J Virol 74(15), 6760-8.
Koyano, S., Mar, E. C., Stamey, F. R., and Inoue, N. (2003). Glycoproteins M and N of
human herpesvirus 8 form a complex and inhibit cell fusion. J Gen Virol 84(Pt 6),
1485-91.
Lifson, J. D., Reyes, G. R., McGrath, M. S., Stein, B. S., and Engleman, E. G. (1986).
AIDS retrovirus induced cytopathology: giant cell formation and involvement of
CD4 antigen. Science 232(4754), 1123-7.
227
Rand, R. P., Fuller, N., Parsegian, V. A., and Rau, D. C. (1988). Variation in hydration
forces between neutral phospholipid bilayers: evidence for hydration attraction.
Biochemistry 27(20), 7711-22.
Roizman, B. (1962). Polykaryocytosis. Cold Spring Harb Symp Quant Biol 27, 327-42.
Sodroski, J., Goh, W. C., Rosen, C., Campbell, K., and Haseltine, W. A. (1986). Role of
the HTLV-III/LAV envelope in syncytium formation and cytopathicity. Nature
322(6078), 470-4.
Spear, P. G. (1985). Glycoproteins specified by herpes simplex viruses. In "The
herpesviruses" (B. Roizman, Ed.), Vol. 3, pp. 315-356. Plenum, New York.
Spear, P. G. (1993). Membrane fusion induced by herpes simplex virus. In "Viral fusion
mechanisms" (J. Bentz, Ed.), pp. 201-232. CRC Press., Boca Raton, Fla.
Takada, A., Robison, C., Goto, H., Sanchez, A., Murti, K. G., Whitt, M. A., and
Kawaoka, Y. (1997). A system for functional analysis of Ebola virus
glycoprotein. Proc Natl Acad Sci U S A 94(26), 14764-9.
Turner, A., Bruun, B., Minson, T., and Browne, H. (1998). Glycoproteins gB, gD, and
gHgL of herpes simplex virus type 1 are necessary and sufficient to mediate
membrane fusion in a Cos cell transfection system. J Virol 72(1), 873-5.
Wiley, D. C., and Skehel, J. J. (1987). The structure and function of the hemagglutinin
membrane glycoprotein of influenza virus. Annu Rev Biochem 56, 365-94.
Xiao, X., Chakraborti, S., Dimitrov, A. S., Gramatikoff, K., and Dimitrov, D. S. (2003).
The SARS-CoV S glycoprotein: expression and functional characterization.
Biochem Biophys Res Commun 312(4), 1159-64.
228
APPENDIX
LETTERS OF PERMISSION
229
October 26, 2004
Our ref: HW/HDN/OCT04/J189
Jeffrey M Melancon
Louisiana State University
Skip Bertman Drive
Baton Rouge, 70803
USA
Dear Mr Melancon
VIROLOGY, Vol 287, No 1, 2001, pp 18 – 29, T P Foster et al, “An Alpha….”
As per your letter dated 2004, we hereby grant you permission to reprint the aforementioned material at no charge in your thesis,
in print and on the Louisiana State University web site subject to the following conditions:
1.
If any part of the material to be used (for example, figures) has appeared in our publication with credit or acknowledgement
to another source, permission must also be sought from that source. If such permission is not obtained then that material
may not be included in your publication/copies.
2.
Suitable acknowledgment to the source must be made, either as a footnote or in a reference list at the end of your
publication, as follows:
“Reprinted from Publication title, Vol number, Author(s), Title of article, Pages No., Copyright (Year), with permission
from Elsevier”.
3.
Reproduction of this material is confined to the purpose for which permission is hereby given.
4.
This permission is granted for non-exclusive world English rights only. For other languages please reapply separately for
each one required. Permission excludes use in an electronic form other than as specified above.
5.
This includes permission for UMI to supply single copies, on demand, of the complete thesis. Should your thesis be
published commercially, please reapply for permission.
Yours sincerely
Helen Wilson
Rights Manager
Your future requests will be handled more quickly if you complete the online form at
www.elsevier.com/locate/permissions
230
231
VITA
Jeffrey Michael Melancon was born April 1, 1978, to Beverly and Mike
Melancon in Kenner, Louisiana. Two years later, Jeff gained a brother and lifelong
friend when Gregory Micheal Melancon was born. Jeff spent his childhood in Kenner
and played on many athletics teams at Muss Bertolino and Lafreniere Park, while
receiving an elementary education from St. Catherine of Siena in Metairie. In 1996, Jeff
graduated summa cum laude from Jesuit High School, where he participated on the
district and state championship soccer team. Jeff was awarded a scholarship to attend
Louisiana State University and pursued a major in microbiology with a minor in
chemistry, during which time he developed an interest in the molecular biology of human
pathogens. After graduation in 2000, Jeff joined the laboratory of Dr. K. G. Kousoulas to
pursue a doctoral degree in the research of herpes simplex virus. Upon graduation, Jeff
will continue his training by attending the Louisiana State University School of Medicine
in New Orleans, where he hopes to broaden his understanding of molecular biology and
human physiology and make a beneficial contribution to the community through clinical
practice and medical research.
232