Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
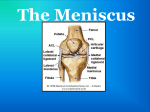
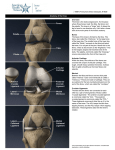
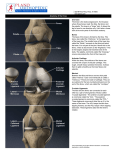
THE ANATOMICAL RECORD 248:269–278 (1997) Development of the Human Knee Joint JUAN A. MÉRIDA-VELASCO,1* INDALECIO SÁNCHEZ-MONTESINOS,1 JOAQUÍN ESPÍN-FERRA,1 JOSÉ F. RODRÍGUEZ-VÁZQUEZ,2 JOSÉ R. MÉRIDA-VELASCO,2 AND JUAN JIMÉNEZ-COLLADO2 1Human and Experimental Embryology Research Group, Department of Morphological Sciences, University of Granada, E-18071 Granada, Spain 2Department of Morphological Sciences II, University Complutense of Madrid, Madrid, Spain ABSTRACT Background: Many studies have been published on the development of the human knee joint, but different investigators disagree on its morphogenetic time table. Most discrepancies center on the cavitation of the knee joint and the participation of the superior tibiofibular joint in the joint knee system. Methods: We summarize our observations of the development of the knee joint in 50 serially sectioned human embryonic and fetal lower limbs (26 embryos and 24 fetuses). Results: The epiphysis of the femur and tibia become condryfied from O’Rahilly stage 18, and ossification begins during the 13th week of development. The patella appears as a dense blastema during O’Rahilly stage 19, becomes condryfied during O’Rahilly stage 22, and begins its ossification during the 14th week of development. The knee joint cavity appears during O’Rahilly stage 22, initially as the femoropatellar joint. This process begins at the periphery of the articular interzone. The superior tibiofibular joint communicates with the lateral meniscotibial joint between 10 and 11 weeks of development and becomes separated from the 13 week on. The menisci arise from the eccentric portions of the articular interzone during O’Rahilly stage 22; however, until week 9 of development, they are not easily distinguishable. Conclusions: We establish the morphogenetic time table of the human knee joint. Anat. Rec. 248:269–278, 1997. r 1997 Wiley-Liss, Inc. Key words: articular interzone; superior tibiofibular joint; meniscus; development; human During recent decades, a wealth of information about the development of bones and joints has been published (Bernays, 1878; Hagen-Torn, 1882; Bardeen, 1907; Walmsley, 1940; Whillis, 1940; Haines, 1947, 1953; Gray and Gardner, 1950; Gardner and O’Rahilly, 1968; among others). More attention has been given to the development of the knee joint than to any other joint. This intense interest must be seen in the light of the size of this joint and of its complexity and clinical importance. The elements that have received the most attention are the cavitation of the knee joint and the role of the head of the fibula in the formation of the knee joint system. The first signs of cavitation of the knee joint are seen in peripheral areas (Gray and Gardner, 1950; O’Rahilly, 1952; Palacios and Rhode, 1980; Cáceres and Caja, 1980) or near the center (Andersen, 1961; Andersen and Bro-Rasmussen, 1961) of the medial layer of the articular interzone; however, Mitrovic (1978) believed the process to take play simultaneously in central and peripheral areas. This process is initially detectable in the femoropatellar interzone (Lucien, 1904; Walmsley, 1940; Gray and Gardner, 1950) and r 1997 WILEY-LISS, INC. appears immediately thereafter in the femoromeniscal interzone. The role of the head of the fibula in the formation of the knee joint has been widely debated. The superior tibiofibular interzone appears during O’Rahilly stage 22 (Gray and Gardner, 1950; Haines, 1952, 1953; Andersen, 1961; Palacios and Rhode, 1980); nevertheless, McDermott (1943) reported that the superior tibiofibular interzone is not yet clearly established during weeks 8 and 9. The cavitation of the superior tibiofibular joint becomes evident during week 10 (Andersen, 1961), in 60-mm specimens (Gray and Gardner, 1950), or during week 15 of development (Palacios and Rhode, 1980). A clear communication forms between the superior tibiofibular and lateral meniscotibial articular cavities during weeks 9.5 (Keith, 1933; Gray and Gardner, 1950) or 10 and 11 of development *Correspondence to: Juan A. Mérida-Velasco, Departamento de Ciencias Morfológicas, Facultad de Medicina, Avda. de Madrid 11, E-18071 Granada, España. Received 23 July 1996; accepted 11 December 1996. 270 J.A. MÉRIDA-VELASCO ET AL. TABLE 1. Features of the specimens used Embryos’ C-R length (mm) X-12 GG-1 BE-1 BB-5 E-19 JD-7 PT-9 JD-2 R-1 MA-7 X-6 PE-8 HA-2 CH-1 X-14 EA-3 BB-4 GV-4 HE-1 FA-5 NA-1 RI-4 X-18 BB-2 H-23 X-4 15 17 17 18 19 19 20 20 21 22 22.5 23 23 24 24 24.5 26 27 28 28 29 29 30 30 31 31 Plane of section O’Rahilly stage Transverse Transverse Transverse Transverse Transverse Transverse Transverse Transverse Transverse Transverse Transverse Transverse Transverse Transverse Transverse Sagittal Transverse Transverse Transverse Transverse Transverse Transverse Transverse Transverse Transverse Transverse 18 18 19 19 20 20 20 20 21 21 21 22 22 22 22 22 22 22 23 23 23 23 23 23 23 23 (McDermott, 1943; O’Rahilly, 1951; Olivier, 1965). However, Gray and Gardner (1950), Haines (1953), Andersen (1961), Palacios and Rhode (1980), Cáceres and Caja (1980) found no evidence that the head of the fibula is involved in the formation of the knee joint during development. These studies are now especially pertinent because of the increasing interest in congenital anomalies of the skeleton. The purpose of this study was to analyze the development and establish the morphogenetic time table of the knee joint. MATERIALS AND METHODS Fifty human embryos and fetuses from the embryo collections of the Department of Morphological Sciences of the universities of Granada and Madrid (Universidad Complutense) were used in the present study. Crown–rump (C-R) length, plane of section, and stage of development (O’Rahilly and Müller, 1987) are shown in Table 1. The usual laboratory procedures were used to prepare 10–15-µm-thick transverse or sagittal serial Abbreviations F L P T = 1 2 3 4 5 6 7 8 9 lower epiphysis of the femur cruciate ligaments patella upper epiphysis of the tibia cartilage canal interzone of the knee joint interzone of the superior tibiofibular joint menisci articular capsule condylopatellar ligaments femoropatellar joint cavity femoromeniscal joint cavity meniscotibial joint cavity superior tibiofibular joint cavity Fetuses’ C-R length (mm) CA-1 RI-1 H-19 BB-1 GV-3 AM-1 PE-7 ZO-1 SA-1 GV-1 MA-3 SA-3 X-8 MA-2 MA-1 X-11 BB-3 H-4 SA-4 PE-3 JM-1 OL-1 ZO-2 BU 35 38 39 39 41 41 41 42 44 45 46 48 50 50 52 53 53 62 63 70.5 80 83 102 113 Plane of section Weeks of development Transverse Transverse Transverse Transverse Transverse Transverse Transverse Transverse Transverse Transverse Transverse Transverse Sagittal Transverse Transverse Transverse Transverse Transverse Transverse Transverse Transverse Transverse Sagittal Transverse 9 9 9 9 10 10 10 10 10 10 10 10 11 11 11 11 11 12 12 12 13 13 14 14 sections, which were stained with hematoxylin–eosin (McManus and Mowry, 1968) for light microscopic study. RESULTS O’Rahilly Stages 18 and 19 During O’Rahilly stage 18, chondrification began at the lower portion of the femur and upper part of the tibia (Fig. 1A). An articular interzone, consisting of a single band of mesenchymal tissue, formed between these two areas (Fig. 1A). At the end of O’Rahilly stage 19, a condensation of mesenchyme representing the future patella became visible ventral and cranial to the condyles of the femur (Fig. 1B). O’Rahilly Stages 20 and 21 Chondrification continued at the lower end of the femur and upper part of the tibia. The femoral condyles Fig. 1. A: Human embryo GG-1. The epiphysis of the femur (F) and tibia (T) begin their condryfication. Between them is the articular interzone of the knee (1). Note that the interzone consists of an unlayered dense mesenchyme. B: Human embryo BB-5. The articular interzone of the knee (1), as an unlayered dense mesenchyme, is located between the epiphysis of the femur (F) and tibia (T). Ventrally to the femur, the organization of the patella (P) begins as a dense blastema. C: Human embryo MA-7. Condryfication of the epiphysis of the femur (F) and tibia (T) continue, and the first signs of organization of the femoral and tibial condyles are observed. Between these, the articular interzone of the knee (1) begins to form as a three-layered structure. D: Human embryo CH-1. The articular interzone of the knee (1) appears as a three-layered structure. Two dense eccentric layers lean against the condyles of the femur (F) and tibia (T) and represent the future articular cartilage. Between these layers is a lax medial layer. Bars 100 µm in A,B, 200 µm in C, 50 µm in D. Fig. 1. 272 J.A. MÉRIDA-VELASCO ET AL. and upper surface of the tibia began to be distinguishable (Fig. 1C). The articular interzone of the knee was formed of two eccentric bands of mesenchymal tissue that covered the condyles of the femur and upper surface of the tibia (Fig. 1C) and a medial band that began to appear more lax in comparison with the eccentric layers (Fig. 1C). O’Rahilly Stage 22 The eccentric bands of the interzone followed the shape of the femoral and tibial condyles, forming a dense band of perichondrial connective tissue (Fig. 1D) that marked the first sign of organization of the articular cartilage. At the same time, the medial layer of the interzone appeared more lax than the eccentric layers (Fig. 1D). The lateral parts of the interzone became densified and began to form the menisci (Fig. 2A). The joint capsule began to attach peripherally to the menisci (Fig. 2A & 2B). The medial layer of the interzone appeared lax. Laterally, at the level of the medial layer, small cavities began to appear between the meniscus and the lateral condyles of the femur and tibia. These cavities were crossed by conjunctive tissue septae (Fig. 2B). Chondrification of the patella began, as did cavitation of the femoropatellar interzone (Fig. 2C). On the upper surface of the tibia, the intercondylar eminence began to form, and immediately dorsal to this, the tibial insertion of the posterior cruciate ligament appeared (Fig. 2C). In the superior tibiofibular interzone, no sign of cavitation was apparent (Fig. 2D). O’Rahilly Stage 23 The articular capsule was visible, and densification of the condylopatellar ligaments was evident (Fig. 3A). From the lateral margins of the patella, the articular capsule surrounded the femoral condyles (Fig. 3A) and became attached to the eccentric surface of the menisci (Fig. 3B). The femoropatellar (Fig. 3A), femoromeniscal, and meniscotibial joint cavities (Fig. 3A & 3B) were distinguishable. The interzone of the superior tibiofibular joint was visible but showed no signs of cavitation (Fig. 3C). Week 9 Development of the menisci proceeded, giving rise to the femoromeniscal and meniscotibial joint cavities (Fig. 3D & 3E), in which a few tracts of connective tissue were still visible. The patella lay opposite of, and was articulated mainly with, the lateral condyle. The triangular space with an anterior base that formed below the patella was occupied by mesenchymal tissue, which may represent the first sign of formation of the fat pad (Fig. 3D). Peripherally, the menisci were attached by the coronary ligaments to the capsule (Fig. 3E). Weeks 10 and 11 The degree of development of the knee joint at this time was similar to that seen during week 9, with slight modifications. The superior tibiofibular joint cavity first appeared, and a connection between this cavity and the lateral meniscotibial cavity was visible (Fig. 4A). Development of the medial meniscus continued and brought about the organization of the internal femoromeniscal and meniscotibial joints (Fig. 4B). The anterior horn of the medial meniscus was attached to the anterior aspect of the upper surface of the tibia (Fig. 4B). The lateral meniscus was clearly evident, and its formation brought about the organization of the lateral femoromeniscal and meniscotibial joints (Fig. 4C). Weeks 12 and 13 The knee joint cavity attained its adult appearance during this time, when the communication between the lateral meniscotibial and superior tibiofibular cavities disappeared. During week 13, ossification began in the epiphyses of the elements comprising the knee joint and first became apparent in the lower epiphysis of the femur and upper epiphysis of the tibia (Fig. 5A & 5B). Cartilage canals invaded the perichondrial zone of the condyles and penetrated from superficial to deep areas (Fig. 5A & 5B). In the femur, these canals penetrated preferentially from the margins and deep portion of the intercondylar notch of the femur (Fig. 5A). In the tibia, they penetrated preferentially from the anterior and superior margins (Fig. 5B). The invading vessels came from the pericapsular arterial network, which in turn was dependent on the genus arteries. Week 14 Ossification of the patella began, with cartilage canals penetrating from the anterior and superior surface (Fig. 5C). DISCUSSION The elements that constitute the knee joint begin to chondrify during O’Rahilly stage 18 (Gardner and O’Rahilly, 1968; Cáceres and Caja, 1980; Palacios and Rhode, 1980; Clark and Odgen, 1983), when the apposed ends of the femur and tibia are covered with a dense layer of perichondrium. However, Andersen (1961) noted that in 20-mm embryos, ‘‘the distal end of the femur as well as the proximal end of the tibia and fibula still persist as a dense blastemal precartilage.’’ McDermott (1943) reported that chondrification of the femur and the tibia began on days 47–51 of development, and O’Rahilly (1952) first observed chondrification during stage 27. O’Rahilly et al. (1956) reported chondrification of the femur and tibia in a few O’Rahilly stage-17 embryos, whereas Haines (1953) found that this process was first evident in the knees of 13-mm embryos. The patella first becomes visible at the end of O’Rahilly stage 19 (Bardeen, 1907; Gardner and O’Rahilly, 1968). Some investigators have reported that it makes its first appearance later, when the embryo has reached 20 mm (Walmsley, 1940; Haines, 1947), 23 mm (Andersen, 1961), 24.5 mm (Caja and Cáceres 1980), 42 mm (Eberl-Rothe and Sonnenschein, 1950), or 7.5 weeks of development (Gray and Gardner, 1950). According to Gardner and O’Rahilly (1968), the patella begins to undergo chondrification during O’Rahilly stage 22, although they also noted that in four of the HUMAN KNEE JOINT Fig. 2. A: Human embryo PE-8. The lateral portions of the articular interzone of the knee form the menisci (3) that are located between the condyles of the femur (F) and tibia (T). The knee joint capsule (4) is attached to the eccentric margin of the menisci. B: Human embryo PE-8. Formation of the meniscofemoral (7) and meniscotibial (8) knee joint cavities has begun. Both cavities are crossed, at this time, by mesenchymal trabeculae. Knee joint capsule (4) is attached to the 273 eccentric margin of the menisci. C: Human embryo PE-8. condryfication of the patella (P) and the organization of the femoropatellar cavity (6) have begun. The posterior cruciate ligament (L) is observed in the medial layer of the knee joint interzone (1). D: Human embryo EA-3. The articular interzone of the superior tibiofibular joint (2) is observed. At this time, no sign of cavitation is evident. Bars 200 µm in A,C,D, 100 µm in B. 274 J.A. MÉRIDA-VELASCO ET AL. Fig. 3. HUMAN KNEE JOINT specimens they examined, signs of chondrification were evident during O’Rahilly stage 21. Walmsley (1940) reported the earliest signs of chondrification in 30-mm embryos, whereas McDermott (1943) distinguished this phenomenon during fetal development at 9 and 10 weeks. The epiphysis of the femur and tibia begin to ossify during week 13, and in the patella, ossification begins during week 14. However, Gray and Gardner (1950) and Palacios and Rhode (1980) found signs of the formation of cartilage canals in the knee in 12-week-old fetuses. The vessels penetrated from superficial to deep levels after invading the perichondrial zone of the condyle. In the lower epiphysis of the femur, vessels invaded from the margins and the deep portion of the intercondylar notch, and in the upper epiphysis of the tibia, invasion proceeded from its anterior and superior part surfaces (Palacios and Rhode, 1980). In the patella, vessels penetrated from the superior and anterior margins (Kreutz and Kreutz, 1981; Doskocil, 1985) and not from the inferomedial margin of the kneecap, as noted by Palacios and Rhode (1980). Formation of the capsule of the knee joint begins during O’Rahilly stage 22. According to Archer et al. (1994), hyaluronan appears to play a central role in this process. However, in one of the O’Rahilly stage-23 embryos studied by Gardner and O’Rahilly (1968), there was no evidence of the joint capsule in the knee; and in another stage-23 embryo, one of the knees showed signs of incipient cavitation near the femur and patella. In material studied by Bardeen (1907), Gray and Gardner (1950), and O’Rahilly and Gardner (1975), cavitation began during O’Rahilly stage 23; however, Walmsley (1940), Haines (1947), Moll (1948), and Palacios and Rhode (1980) first observed this process at 9 weeks of development, and Dubinkin and Motnenko (1931) noted this process in 11.8-cm fetuses. The first signs of cavitation of the knee joint are seen in peripheral areas of the medial layer of the articular interzone (Gray and Gardner, 1950; O’Rahilly, 1952; Cáceres and Caja, 1980; Palacios and Rhode, 1980). This process is initially detectable in the femoropatellar interzone (Lucien, 1904; Walmsley, 1940; Gray and Gardner, 1950) and appears immediately thereafter in the femoromeniscal interzone. However, Andersen and Bro-Rasmussen (1961) and Andersen (1961) reported that cavitation began near the center of the interzone and spread toward the periphery, whereas (Mitrovic, 1978) believed the process to take play simultaneously Fig. 3. A: Human embryo X-4. The femoropatellar (6) and meniscofemoral (7) cavities are observed. The knee joint capsule (4), attached to the eccentric margin of the menisci (3), is strengtened by the condylopatellar ligaments (5). B: Human embryo NA-1. The meniscofemoral (7) and meniscotibial (8) cavities are clearly distinguishable. In the middle of the knee joint, organization of the cruciate ligaments (L) continues. Knee joint capsule (4). C: Human embryo HE-1. At the level of the superior tibiofibular joint (2), a three-layered interzone is present; hence, no sign of articular cavitation is evident. D: Human fetus RI-1. The patella (P) is basically articulated with the lateral condyle of the femur (F). Below the patella, a triangular space forms. This space is occupied by a mesenchymal tissue that gives rise to the intraarticular pad of fat. Lateral meniscus (3). E: Human fetus RI-1. Enlargement of D. Observe the knee joint capsule (4) attached to the eccentric margin of the menisci (3), which organize the coronary ligament. The femoromeniscal (7) and meniscotibial (8) cavities are clearly distinguishable. Bars 200 µm in A–D, 100 µm in E. 275 in central and peripheral areas, although this author noted that cavitation was more advanced at the periphery. The role of the head of the fibula in the formation of the knee joint has been widely debated. In our material, we observed the appearance of a superior tibiofibular interzone during O’Rahilly stage 22 (Gray and Gardner, 1950; Haines, 1952, 1953; Andersen, 1961; Palacios and Rhode, 1980). Nevertheless, McDermott (1943) reported that the superior tibiofibular interzone was not yet clearly established in weeks 8 and 9. We also found that cavitation of the superior tibiofibular joint became evident during week 10 (Andersen, 1961) and not in 60-mm specimens (Gray and Gardner, 1950) or during week 15 of development (Palacios and Rhode, 1980). A clear communication forms between the superior tibiofibular and lateral meniscotibial joint cavities during weeks 10 and 11 (McDermott, 1943; O’Rahilly, 1951; Olivier, 1965). This space was also seen by Keith (1933) and Gray and Gardner (1950) in a 9.5-week fetus. However, Gray and Gardner (1950), Haines (1953), Andersen (1961), Palacios and Rhode (1980), and Cáceres and Caja (1980) found no evidence that the head of the fibula was involved in the formation of the knee joint during development, and Gray and Gardner (1950) reported that in most cases the knee joint cavity was separated from the superior tibiofibular joint cavity by either a small amount of very loose tissue or very thin strands of somewhat denser tissue. O’Rahilly (1951) described a femorofibular interzone in embryos of 16–20 mm, although in our material no such interzone was visible. Discrepancies also exist with regard to the final organization of joint cavity, considered to arise from the confluence of five initially separate cavities: the femoropatellar, two femoromeniscal, and two meniscotibial cavities (McDermott, 1943; Gray and Gardner, 1950; Palacios and Rhode, 1980). However, one investigator considered only three cavities to be involved, i.e., the femoropatellar and two femorotibial cavities (Andersen, 1961), and another investigator (Doskocil, 1985) reported that only two symmetrical cavities initially separated by a medial mediastinum were involved. Our observations show that initially a femoropatellar cavity and two femoromeniscal cavities exist and that subsequently two meniscotibial cavities are formed. Nonetheless, a sixth space, the superior tibiofibular cavity, is in communication with the meniscotibial cavity during weeks 10 and 11 of development and thus forms part, at least temporarily, of the developing knee joint cavity. The formation of a single cavity, with the disappearance of communication with the superior tibiofibular cavity, takes place during weeks 12 and 13 of development according to Palacios and Rhode (1980) and during the eighth week according to Keith (1933). Organization of the menisci begins during O’Rahilly stage 22 (Haines, 1947; Andersen, 1961; Gardner and O’Rahilly, 1968; Cáceres and Caja, 1980; Clark and Odgen, 1983), although Gardner and O’Rahilly (1968) observed the lateral meniscus in one of the nine O’Rahilly stage-20 specimens they studied. The menisci cannot be distinguished clearly until week 9 (Gray and Gardner, 1950; Palacios and Rhode, 1980); however, some investigators have reported that they form at later stages, i.e., week 8 (Palacios and Rhode (1980), 276 J.A. MÉRIDA-VELASCO ET AL. Fig. 4. A: Human fetus GV-1. The superior tibiofibular joint cavity (9) shows its communication with the lateral meniscotibial joint cavity (8). Lateral meniscus (3). Posterior cruciate ligament (L). B: Human fetus GV-1. The medial meniscus (3) is completely formed, and the medial meniscofemoral (7) and meniscotibial (8) joint cavities are established. The anterior horn of the medial meniscus is attached to the anterior aspect of the upper surface of the tibia. Anterior cruciate ligament (L). C: Human fetus BB-3. The lateral meniscus (3) is completely organized, and the lateral meniscofemoral (7) and meniscotibial (8) joint cavities are established. Observe the communication between the superior tibiofibular (9) and the lateral meniscotibial (8) joint cavities. Bars 5 200 µm. HUMAN KNEE JOINT 277 Fig. 5. A: Human fetus OL-1. Cartilage canal (arrow) penetrates the lower end of the femur at the level of its posterior surface. B: Human fetus OL-1. Cartilage canals (arrow) penetrate the upper end of the tibia. Ossification begins on the superior and anterior surfaces of the tibia. The anterior horn of the lateral meniscus (3) is also distinguishable. C: Human fetus ZO-2. Cartilage canals (arrow) penetrate the superior and anterior surface of the patella (P). Bars 100 µm in A,B, 200 µm in C. 278 J.A. MÉRIDA-VELASCO ET AL. weeks 9 and 10 (McDermott, 1943), or even as late as in the 3.5-month fetus (Dubinkin and Motnenko, 1931). The joint capsule is attached to the anterior margin of the external meniscus during O’Rahilly stage 22 and to the eccentric surface of both menisci during O’Rahilly stage 23. According to McDermott (1943) and Gray and Gardner (1950), this final attachment does not take place until week 12 of development. During week 10 of development, the horns of the menisci clearly become attached to the anterior and posterior aspects of the upper surface of the tibia. ACKNOWLEDGMENTS We thank Ms. Karen Shashok for translating the original manuscript into English. LITERATURE CITED Andersen, H. 1961 Histochemical studies on the histogenesis of the knee joint and superior tibio-fibular joint in human foetuses. Acta Anat., 46:279–303. Andersen, H., and F. Bro-Rasmussen 1961 Histochemical studies on the histogenesis of the joints in human fetuses with special reference to the development of the joint cavities in the hand and foot. Am. J. Anat., 108:111–122. Archer, C.W., H. Morrison, and A.A. Pitsillides 1994 Cellular aspects of the development of diarthrodial joints and articular cartilage. J. Anat., 184:447–456. Bardeen, C. 1907 Studies of the development of the human skeleton. Am. J. Anat., 4:265–302 Bernays, A. 1878 Die Entwicklungsgeschichte des Kniegelenkes des Menschen, mit Bemerkungen über die Gelenke im Allgemeinen. Morph. Jb., 4:403–446. Cáceres, E., and V.L. Caja 1980 Estudio de las cavidades intraarticulares de la rodilla en los perı́odos embrionario y fetal humano. An. Desarr., 56:79–85. Clark, C.R., and J.A. Odgen 1983 Development of the menisci of the human knee joint. Morphological changes and their potential role in childhood meniscal injury. J. Bone Joint Surg., 65:538–547. Doskocil, M. 1985 Formation of the femoropatellar part of the human knee joint. Folia Morph., 33:38–47. Dubinkin, G., and A. Motnenko 1931 Uber die Entwicklung der Menisken des Kniegelenks. Arch. Klin. Chir., 8:165–178. Eberl-Rothe, G., and A. Sonnenschein 1950 Die ontogenetische Ausbildung des Kniegelenkes beim Menschen. Zschr. Anat. Entwgsch., 115:251–272. Gardner, E., and R. O’Rahilly 1968 The early development of the knee joint in staged human embryos. J. Anat., 102:289–299. Gray, D.J., and E. Gardner 1950 Prenatal development of the human knee and superior tibiofibular joints. Am. J. Anat., 86:235–287. Hagen-Torn, O. 1882 Entwickelung und Bau der Synovialmembranen. Arch. Mikr. Anat., 21:591–663. Haines, R.W. 1947 The development of joints. J. Anat., 81:33–55. Haines, R.W. 1952 The development of the femoro-tibial joint with a discussion of the alleged tibio-fibular articulation. J. Anat., 86:495– 503. Haines, R.W. 1953 The early development of the femoro-tibial and tibio-fibular joints. J. Anat., 87:192–206. Keith, A. 1933 Human Embryology and Morphology, 5th ed. Williams Wood and Co., Baltimore. Kreutz, W., and R. Kreutz 1981 Die Knorpelkanäle der menschlichen Patella bis zur Geburt. Gegenbaur Morphol. Jahrb., 127:620–624. Lucien, M. 1904 Développement de l’articulation du genou et formation du ligament adipeux. Bull. Assoc. Anat., 3:133– 135. McDermott, L.J. 1943 Development of the human knee joint. Arch. Surg., 46:705–719. McManus, J.F.A., and R.W. Mowry 1968 Técnica Histológica. Atika S.A., Madrid. Mitrovic, D. 1978 Development of the diarthrodial joints in the rat embryo. Am. J. Anat., 115:475–486. Moll, J. 1948 The embryonic development of the human knee joint. Act. Neerl. Morphol. Nor. Pathol., 6:283–284. Olivier, G. 1965 Anatomie Antropologique. Vigot Frères, Paris. O’Rahilly, R. 1951 The early prenatal development of the human knee joint. J. Anat., 85:166–170. O’Rahilly, R. 1952 The development of the synovial joints in the chick. Anat. Rec., 112:303–375. O’Rahilly, R., and E. Gardner 1975 The timing and sequence of events in the development of the limbs in the human embryo. Anat. Embryol., 148:1–23. O’Rahilly, R., and F. Müller 1987 Developmental Stages in Human Embryos. Carnegie Institution of Washington, no. 637, Washington, D.C. O’Rahilly, R., E. Gardner, and D.J. Gray 1956 The ectodermal thickening and ridge in the limbs of staged human embryos. J. Embryol. Exp. Morphol., 4:254–265. Palacios, J., and M. Rhode 1980 Embriologı́a de la rodilla. In: IV Symposium Internacional de Traumatologı́a Guillén, P., ed. Fundación Mapfre, Madrid, pp. 3–25. Walmsley, R. 1940 The development of the patella. J. Anat., 74:360– 368. Whillis, F. 1940 The development of synovial joints. J. Anat., 74:277– 283.