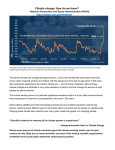
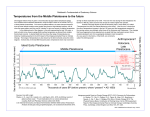
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
The Pleistocene and the Origins of Human Culture: Built for Speed Peter J. Richerson Department of Environmental Science and Policy University of California Davis, California USA 95616 [email protected] Robert Boyd Department of Anthropology University of California Los Angeles, California USA 90024 [email protected] Abstract. A number of authors have advanced the argument that the onset Pleistocene climate fluctuation is responsible for the evolution of human anatomy and cognition. This hypothesis is in contrast to the common idea that humans represent a revolutionary breakthrough rather than a conventional adaptation to a particular ecological niche. The Pleistocene hypothesis is, as proposed, not wen specified. How did Pleistocene fluctuations specifically favor the particular adaptations that characterize humans? The revolutionary breakthrough hypothesis is similarly weak. Our large brain and all it can do does seem responsible for our present dominance of the earth. If so, what has prevented many animal lineages in the remote past from evolving large brains? Theoretical models of the cultural evolutionary process suggest some answers to these questions. Learning, including social learning, is rather generally a useful adaptation in variable environments. The progressive brain enlargement in many mammalian lineages during the last few million years suggests that climatic deterioration has had the general effect predicted by the Pleistocene hypothesis. Increased dependence on simple social learning was a preadaptation to the evolution of a capacity for complex traditions. The evolution of a costly capacity to acquire complex traditions is inhibited because, initially, complex traditions will be rare. Having the capacity to learn things that are far too complex to invent for oneself is not useful until traditions are common, but traditions cannot become common before the capacity to acquire them is common. This problem may explain why many animals became more sophisticated learners in the Pleistocene, but why only the human lineage found its way around the not-favored-when-rare barrier, and thus evolved the capacity for complex cultural traditions. Version 1.1. February, 1998. For presentation at 5th Biannual Symposium on the Science of Behavior: Behavior, Evolution, and Culture. February 23, 1998, University of Guadalajara, Mexico. Please do not cite without permission. Comments welcome! Introduction The evolution of humans is a major event in the Earth's biotic history. This statement is not just anthropocentric hubris. Never before has a single species of organism dominated the planet to the extent that we do. Even before the development of food plant production, hunting and gathering peoples penetrated to every continent except Antarctica. Most accounts of human origins take the adaptive superiority of our species for granted. If so, the interesting question is what pre-adaptive breakthrough that led to the transcendent, fully human, hyper-adaptive complex of characters-tool making, language, complex social organization, and the like. According to Landau (1984) accounts of human origins read like heroic fables. The identical title, The Ascent of Man, that Bronowski (1973) gave his popular account and Pilbeam (1972) his textbook illustrate the pattern. This form is a consequence of the assumption of transcendent adaptive superiority. More recently, following Alexander (1974) and Wilson (1975), human sociobiologists have pursued a different line of research. They assume that humans, in the words of Foley (1987), are just "another unique species." In the theory of evolution by natural selection, the driving force of change is adaptation to local environments. These scholars are suspicious of taking human adaptive superiority for granted or of granting human culture and its products special status. Rather than being a history of a breakthrough to a new adaptive plane, human evolution is more likely a history of adaptation to local environments that happens to have resulted in our current ecological dominance by accident. The deterioration of the Earth's climate since the Miocene, leading to the Pleistocene ice age during the last 2 million years, is a major event in the history of the planet's physical environment. Over the last 6 million years, the climate has gotten colder, drier, and more variable. Geology records several other glacial episodes, but the last was the PermoCarboniferous ice ages that ended 250 million years ago (Lamb, 1977:296). Perhaps the ongoing evolution of the hominid lineage was driven by the ongoing changes in the Earth's climate. DeMenocal (1996) and Potts (1996) give impressive evidence in favor of this hypothesis. In this paper, we argue that the specific mechanism by which humans mastered the Pleistocene is our capacity to evolve adaptations to the variation of Plio-Pleistocene environments via cultural traditions. The results of theoretical modeling of cultural evolution suggest that the basic adaptive function of social learning is an enhanced ability to respond to temporal and spatial variations in the environment. Many animal lineages seem to have taken advantage of the potential of social learning. In many respects, human culture is nothing more than a straight-forward adaptation to climatic deterioration. Humans do differ from proto-cultural animals in having the ability to evolve complex, multi-part traditions that must evolve cumulatively, normally over many generations. Technology and social organization furnish many examples of complex traditions that evolve by descent with modification like complex organic adaptations, albeit at a faster rate. Other animals show non-existent to marginal abilities to acquire complex traditions. Whether complex traditions truly represent a breakthrough to a qualitatively superior system of adaptation or only an especially clever mechanism to adapt to the Pleistocene is a moot question. Plio-Pleistocene Climate Deterioration In the quarter-century since Shackelton and Opdyke (1973) published the first detailed ocean sediment core spanning the entire Pleistocene, our understanding of this episode has deepened remarkably. Using a variety of proxy measures of past temperature, rainfall, ice volume, and the like, mostly from cores of ocean sediments, lake sediments, and ice caps, paleoclimatologists have constructed a stunning picture of climate deterioration. Lamb (1977), Schneider and Londer (I 984), and Dawson (1992) give accessible reviews of the methods used. Since the mid-Miocene, about 14 million years ago, the Earth's temperature has dropped several degrees. The causes of the drop are not well understood, but are probably the result of basic geological processes (Partridge, et al., 1995). The arrangement of the continents is probably important. Continental drift has produced a situation with Antarctica in the South Polar region, insulated from warm ocean currents by the circumAntarctic currents. Similarly, the Arctic Ocean is closely surrounded by land masses, insulating it from the penetration of warm ocean currents to the north polar region. Ice cover at high latitudes reflects much sunlight back to space, significantly lowering the Earth's total heat income. Slowing sea-floor spreading rates over the whole Tertiary period may have lowered the output of CO2 from the Earth's interior. A lower concentration of this important greenhouse gas in the atmosphere win result in a lower global temperature. The amplitude of fluctuations in rainfall and temperature increased as mean temperature dropped. Figure 1 from Opdyke (1995) shows schematically how the envelope of temperature fluctuations has increased step-wise since the Miocene, as measured in Deep-Sea Drilling Project cores. As time series analysts say, the climate record of the past few million years is highly non-stationary. Not only does the climate vary, but the statistics that describe the variation-the variance and patterns of autocorrelation--change with time. The pattern of fluctuation in climate is very complex. Much of the variation seems to arise from an enhanced sensitivity to radiation changes caused by periodic variations in the Earth's orbit. The radiation income in high northern latitudes has a 20% range of variation due to these effects. Milankovitch (1941) developed this hypothesis in its modem form. Broecker and Denton (1990) give a good introductory discussion to this and other aspects of the physics of climate variation. The eccentricity of the Earth's orbit varies on a 95,800 year time scale, the inclination of its axis with a periodicity of 41,000 years, and the precession of the equinoxes with a periodicity of 21,700 years. The magnitude of the direct forcing of climate by these cycles is out of phase in the Northern and Southern Hemispheres, yet the cycles of ice growth and decay are in phase. The Milankovitch cycles reach far back into the earth's history, but the ice age began in earnest only two million years ago. As the deterioration has proceeded, different cycles have dominated the pattern. The 21,700 year cycle dominated the early part of the period, the 41,000 year cycle between about 3 and 1 million years ago, and the 95,800 year cycle from 1 million years ago to the present (de Menocal and Bloemendal, 1995). Thus, complexities of the response of the atmosphere-ocean-ice sheet system must somehow amplify and coordinate the effects of the orbital periodicities. Alternatively, the Milankovitch theory may be incorrect (Broecker, 1992; Brownlee, 1995). Most likely, the present disposition of the continents and oceans affects ocean currents and wind patterns in such a way as to make the global climate very sensitive to small fluctuations in insolation. For example, the Arctic Ocean is sufficiently isolated from the warm Atlantic and Pacific oceans that it is frozen. Its ice cover reflects the sunlight falling on it, substantially preventing it from storing heat in summer. This in turn means that the high Northern latitudes can build ice sheets, which reflect still more sunlight, leading to other chilling. The small forcing from the Milankovitch cycles in high northern latitudes can thus be amplified by the growth and wasting of continental ice in the north. The exact driving mechanisms of the late Cenozoic climate system are still unknown. The most influential hypothesis is that of Broecker et al. (1985). The ocean-atmosphere coupling includes elements that are affected by the deep circulation of the ocean. Under the current climatic regime, the North Atlantic near Greenland is the source of much of the bottom water for the World Ocean. A subsurface current of warm, rather saline, water moves north in the Atlantic and upwells during the winter near Greenland. This water then loses an immense amount of heat to the atmosphere, becomes heavier, and sinks to become the North Atlantic Deep Water. NADW eventually flows out the South Atlantic to the Indian and Pacific Oceans where it reaches the surface in upwelling zones. Thus, there is a great ocean conveyor concentrating immense amounts of heat in the North Atlantic and moderating the climate of the surrounding land masses, especially NW Eurasia. (When the poles were not occupied by or surrounded by continents, similar conveyors presumably kept the poles ice free.) Among other things, the conveyor circulation efficiently draws greenhouse gases, especially CO2, into the ocean abyss. This circulation apparently shuts down during glacial conditions, probably because glacial melt water flowing from the Eastern North American ice sheet makes the ocean surface water too fresh and hence too light to sink even when chilled to maximum density. Once in a glacial mode, the growth of reflective ice sheets and the dust from the dry climates of the glacial period further chill the earth. Modeling work indicates that at least parts of Broecker's scenario are plausible (Manabe and Stouffer, 1995). For the last 120,000 years, data is available from ice cores taken from the deep ice sheets of Greenland and Antarctica with time resolution as high as few decades. During the last glacial (65,000-12,000 years before present, Emiliani oxygen isotope stages 2-4), the climate was highly variable on time scales of centuries to millennia (GRIP, 1993; Lehman, 1993; Ditlevsen, et al., 1996), as can be seen in figure 2. Even when the climate is in the grip of the ice, there were brief excursions of about a thousand years duration, called Dansgaard-Oeschger events, in which the climate briefly reached interglacial or near interglacial warmth. The Dansgaard-Oeschger events themselves come in clumps in which the most extreme warm spikes are just preceded by the appearance of ice-rafted debris in North Atlantic sediments, called Heinrich events. The most highly touted hypothesis to explain the Dansgaard-Oeschger-Heinrich excursions is the cyclic production of glacial melt waters that affect the NADW conveyor, wind patterns over the North Atlantic, and other major regulators of global climate (Lehman, 1993). The last Interglacial (65,000-130,000 years before present, oxygen isotope stage 5) may also have been highly variable on the millennial time scale. Interpretation of the deeper portions of the Greenland ice cores is controversial because of the possibility that ice from colder and warmer periods has been folded by ice movement to create false fluctuations (Grootes, et al., 1993). Nevertheless, many lower-resolution records of the last interglacial also suggest that it was frequently punctuated by episodes of near-glacial cold (e.g. Lamb 1977: 333). As figure 2 shows, the Holocene interglacial period of the last I 0,000 years (oxygen isotope stage 1) has been extraordinarily tranquil. The resolution of the ice core data is extraordinary. Near the top of the GRIP core the resolution to seasons is possible, and at 0 1,000 years before present resolution is estimated to by 1O years (Ditlevsen, et al. 1996). High resolution data do not yet exist prior to the last Interglacial to indicate just how unusual the Holocene is compared to the whole Pleistocene record. The possibility that Holocene climate stability might be easily tipped into a regime of much greater variability by relatively weak forcing is cause for considerable worry regarding human caused increases in carbon dioxide and other greenhouse gasses (Broecker, 1997). High latitudes chilled much more than the equator as the climate cooled during the latter part of the Cenozoic. New habitats arose, such as periglacial tundras. Other habitats, especially arid and semiarid deserts and grasslands replaced forests over large areas from equatorial to temperate latitudes. The spatial as well as the temporal variability of the earth thus increased considerably as the Plio-Pleistocene deterioration advanced. Culture As An Adaptation To Variable Environments Simple Models of Social Learning It seems intuitively as if social learning should be especially useful as a means of coping with spatially or temporally varying environments. Social learning, Eke ordinary individual learning, is a mechanism of phenotypic adaptation. Phenotypic adaptations, by acquiring some information about local environmental conditions, allow organisms to adjust their anatomy and behavior contingently in situations where environmental conditions vary on too small a scale to adapt by genetic adaptations. Presumably, systems of phenotypic adaptation have costs. In the case of learning, an individual will have to expend time and energy in learning, incur some risks in trials that may be associated with large errors, and support the neurological machinery necessary to learn. Social learning can economize on the trial and error part of learning; if kids learn from mom, they can avoid repeating her mistakes. Learning requires sensory organs to obtain information from the environment, some sort of decision-making system to evaluate the outcome of trials, and a memory system to store the results. Social learning may also economize on evaluation costs; what worked for Mom will probably work for her kids. At least, she,, because of inclusive fitness considerations, has only limited reasons to deliberately deceive them. Sophisticated learning systems will require larger sensory and nervous systems. Considering only the energetic costs of maintaining the nervous tissue, the incremental costs are quite large (Eisenberg, 1981: 235-6). Martin (1981) reports that mammalian brains vary over about a 25-fold range, controlling for body size. Human brains are about 5 times as large as the brains of average mammals of our body weight. Modem ungulates and carnivores have average brains. Average living mammals in turn have brains about 5 times as large as those of the smallest brained mammals, such as insectivores and many marsupials. Aiello and Wheeler (1995) report that human brains account for 16% of our basal metabolism. Thus average mammals will have to allocate only about 3% of basal metabolism to their brains, and some get by with less than 1%. Total metabolism runs about 1.8 times that of resting metabolism, mostly because of the mobilization of large masses of otherwise low-metabolic-rate skeletal muscle during exercise. How nervous system metabolic rate varies with "exercise" is poorly understood. Even disregarding mental exercise, humans must expend something like 9% of their total metabolism on their brain versus a little more than 1% for average animals and well under one for the least smart mammals. These differences are large enough to generate significant evolutionary tradeoffs. Aiello and Wheeler hypothesize that one major tradeoff in the human case is that we sacrificed gastrointestinal tissue, which is a little more energyintensive even than nervous tissue. This sacrifice, in turn requires the human diet to focus on higher quality foods to compensate for inefficient assimilation. Note that we have left out of this calculation other significant costs of big brains such as increased difficulty at birth, greater vulnerability to head trauma, and the time and trouble necessary to fill it with usable information. The overhead cost of our large brain must be on the order of s = -.2 (thinking here of the common expression of fitness differentials in terms of departures from unity for a reference fitness, here a mammal with an average encephalization). Selection differentials of under 1% can drive evolution right along, so brains will not become large without correspondingly large adaptive payoffs. How do humans foot our large-brain overhead bill? We are still a long ways from being able to construct a complete analysis of these payoffs. Economists and population biologists common study simple, heuristic mathematical models to school their intuitions about complex processes. In this spirit, we have constructed very basic models of the individual/social learning process along the following fines: Suppose that the individual learning process is the primitive state. Virtually all animals show at least rudimentary abilities to learn. When animals like birds and mammals come to have extended maternal care, they have the opportunity to learn socially. In our models, we assume that individuals have two sources of information, their own experience and the vicarious experience of individuals with whom they are in social contact. In two different kinds of models, one based on quantitative characters (Boyd and Richerson, 1985: Ch. 4), and another based on discrete characters (Boyd and Richerson, Boyd and Richerson, 1989), we asked how evolution might optimize the relative dependence on the two sources of information. In the discrete character model, we assumed that there were two behaviors (e.g. forage collectively or solitarily) and two environments, (e.g. wet and dry). There is a fitness benefit for behaving correctly (forage collectively if environment dry, forage alone if wet). Individuals collect some more or less error-prone information about the state of the environment by individual learning. They can also opt to imitate another individual. The degree of dependence on social versus individual learning was controlled by a confidence-interval-like learning threshold, d, that we assume would be set by selection at an optimal point. While growing up individuals gain some idea of the state of the environment. This information will not lie on average, but in a noisy environment, even if the environment is dry on average, some individuals will experience an unusual run of rainy years. They are vulnerable to mistakenly deciding that the state of the environment is wet. The confidence parameter d tells individuals how seriously to take their noisy samples. If d is large, individuals look for definitive evidence that the environment really is in the wet or dry state, say entirely quite wet or quite dry during their formative years. If they do not see such evidence, and most will not, they imitate someone, say Mom, at random. Figure 3 illustrates graphically how the decision rule works. If d is small enough, any information from personal experience is deemed definitive, and reamers go with the main chance as dictated by their personal experience. In a spatially or temporally varying world, some mixture learning and social learning are generally advantageous. In a noisy world, an evolving population is tending to integrate the experiences of many individuals. One can be saved from the perils of small number statistics by trusting a sample of the population, even a sample of 1, over the noisy data from the environment. On the other hand, in a variable environment, the individuals one might learn from: (1) may have gotten caught in an environment switch, (2) might have migrated from nearby environment in the other state, (3) might have unluckily gotten seemingly definitive information that the environment is in the state that it is not. If your personal experience is pretty definitive, it is liable to be the better guess. The optimal confidence rule (value of d) depends upon the nature of the environmental variation and the quality of the evidence available from personal experience. If the environment fluctuates too rapidly in time, or if the spatial habitat mosaic is so tight that individuals often migrate to a patch different from their parents, individuals should depend entirely on their own experience. In such a world, mom's advice is useless, and there is nothing but to trust to the main chance of personal experience. As the statistical resemblance between parental generation and offspring generation increases, it is safer to depend upon imitation and demand ever more definitive personal evidence before breaking with tradition. In a world where the environment hardly ever really changes, but which is rather noisy, a combination of natural selection and rather conservative reliance on own experience will result in a population in which most individuals are doing the right thing. Social learning becomes quite trustworthy. Figure 4 shows how the relative dependence on social versus own experience should evolve in different environmental situations. The quantitative character model is similar in spirit. We imagined one continuous character (frequency of foraging alone versus cooperatively) and a continuum of environments from wet to dry. In any given environment at any one point in time or space, there is an optimal mix of solitary and cooperative foraging. Bayesian considerations suggest that individuals should use a weighted average of social learning and own experience to determine how to behave. The optimal weighting parameter in this model behaves qualitatively just as d does in the discrete character model. It is comforting that two models with a rather different structure give the same results. We used the quantitative model to run a sort of mathematical tournament comparing the fitness advantages of using a genes-plus-individual learning to a system of social learning plus individual learning. Suppose that there is some cost to being able to learn socially. Under what circumstances might there be a fitness advantage to adding social learning to a repertoire where genes represent the wisdom of evolutionary history and individual learning bears the sole weight of running up the phenotypic fine tuning? A typical example of the results is shown in figure 5. The social learning system is a potential advantage over a wide range of conditions. The advantage of social learning is especially large when the environmental variance (VH) is high and the degree of autocorrelation is high. Under the parameter values we chose for illustration at least, the dependence on social learning is often fairly high, on the order an 75% dependence on social models and a 25% dependence on individual learning. At very high autocorrelations, environments become so slowly changing that genes can track perfectly well, and the advantage of social learning disappears. Think of it like this: Social learning is a form of Lamarckian inheritance. It couples a mechanism of phenotypic flexibility, learning, to a scheme for transmitting the results of past learning via social learning. A system of social learning evolves in response to the Lamarckian pressure of learning as well as the pressure of natural selection. We can the Lamarckian force in these models "guided variation" as it acts a lot like an adaptively nonrandom form of mutation. Guided variation causes a population's behavior to track environmental change in time or space more quickly than genes, lacking the Lamarckian property, can. These results nicely support the intuitive argument. Without any further argument, you can see the temptation to attribute the evolution of cultural systems to the onset of Plio-Pleistocene climatic deterioration. The pattern of increasing environmental variation, but variation organized into autocorrelated patterns on time scales from a few generations to many, is just the thing to give social learning an adaptive advantage. Alan Rogers (1989) threw a dash of skepticism onto this argument. In his model, a population composed of two types, social learners and individual learners. The population lives in a variable environment whose state can be in either one of two regimes (e.g. wet or dry). Social learning short-cuts the costly trials of individual learning, and hence in is potentially an adaptive advantage compared to individual learning. Indeed, when social learners are rare they do have an advantage. They practically always copy an individual learner, getting the same, usually adaptive, behavior at lower cost. The trouble is, in Rogers' model, they keep on increasing in frequency until the chances of a social learner copying another social learner becomes high. As the chain connecting copiers to reamers increases, there is an increasing chance that the environment has changed and that the social learner will copy the wrong trait. The only equilibrium in the system is when the fitness of social reamers falls to the same level as reamers. At equilibrium the mean fitness of the population is exactly the same as a population of individual reamers. Through a sort of parasitic system the potential benefits of social learning are dissipated by excessive copying. This effect is robust with respect to several generalizations of the model (Boyd and Richerson, 1995). The reason that in our models social learning does increase the mean fitness of populations in autocorrelated variable environments is that our individuals capture the benefits of both types of learning, whereas Rogers' individuals capture only one or the other. Nevertheless there is an element of the Rogers result in our models too. In an unpublished study, we replicated a classic argument from patent economics. In a situation where the results of a valuable innovation can diffuse to a larger population, the total benefits of the innovation can greatly exceed the private benefits to the innovator. In our models, individuals take risks when they learn, but the costs born of this risk-taking help keep the population current with a changing environment. The optimal amount of individual learning is higher from the group than from the individual point of view. Absent group selection, the Rogers effect will "under supply" individual learning, perhaps often to the point where little if any adaptive advantage in the sense of increasing the mean fitness of the population. We have studied a number of other models in which the rules of social learning are more sophisticated than the copying of a random member of the population like Mom (Boyd and Richerson, 1985: chs. 5-7). For example, a socially learning individual might use several adults as models. If they exhibit two or more behaviors, the social learner might try each out and retain the one with the highest rewards. Most behaviors current in a population are probably better than the trials that individual reamers can attempt on their own. Plagiarism is easier than originality. Gathering a number of plausible initial guesses about the right behavior and using one's own experience to choose the best among them has advantages similar to the guided variation process discussed above. We call the series of forces on cultural evolution that result from non-random social learning "biased transmission." Another interesting pattern in human cultural evolution is our tendency to evolve boundaries between cultures that have the effect of subdividing human populations into semi-isolated sub-populations. Ethnic groups are a common example. Often, such groups are specialized to exploit particular habitats or economic roles (Barth, 1969). Models show that cultural badges-different language, dress, religious practice-can evolve to erect barriers to the free flow of ideas in spatially heterogeneous environments (Boyd and Richerson, 1987). Ethnic groups thus form the cultural analogs of reproductively isolated species. The main difference is that the barriers are much more permeable and the rate of evolution of culture is much higher than that of genes. Human cultural niche shifting is faster than that of animals that adapt mainly by organic evolution. Using this "psuedospeciation7' mechanism, Late Pleistocene humans developed such a diverse array of subsistence economies that our species spread to the ends of the habitable earth (Bettinger, 1991:203-5). Humans may not be the only species that uses this mechanism. Many birds learn their songs by imitating adults, creating local song traditions. Females may prefer to mate with males that sing the songs their fathers sang. Nottebohm (I 975) argued that these dialects subdivide local populations and allow the frequency of locally adapted genes to increase by restricting gene flow between groups. Social Learning A Response to Variability Selection Richard Potts (1996: 231-238) argues that the fluctuating climates of the Plio-Pleistocene have imposed a regime of "variability selection7' on the Earth's biota. He suggests (p. 23 7) that under variability selection "genetic variations favor open programs of behavior that vary and extend the adaptive possibilities of the individual. These are conserved in the gene pool over time because of the inconsistency in the short-term effects of natural selection. Organisms eventually build up an inheritance system that enables them to buffer larger and larger disturbances in the factors governing survival and successful reproduction." This is an interesting supposition, though lacking in detail about mechanisms. We suggest that social learning is one such candidate mechanism. OdlingSmee (this symposium) traces out in some detail how the genetic and social transmission might coevolve under an extended regime of variability selection. Testing this idea is a formidable challenge. Certainly not every lineage on the earth responded to the onset of extreme glacial fluctuations by evolving social learning. Further, humans are the only species to respond to the ice age by evolving the very complex forms of social learning usually given the term "culture." The unique importance of social learning in humans and the spectacular ecological success we become using culture is an embarrassment the hypothesis. How can an environmental event that affected the entire earth account for the evolution of one species' peculiar adaptation? The skeptic might ask, if social learning is an adaptation to the Pleistocene, why aren't many species capable of human-like feats of social learning? Is there any evidence that social learning has anything to do with climatic deterioration? How does social learning fit into a pattern of responses to variability selection if indeed this concept is useful? General Evolutionary Responses to Plio-Pleistocene Climate Deterioration Evolutionists have known the general outline of the Plio-Pleistocene climatic deterioration for a long time, and its evolutionary consequences are the subject of much classical work. For example, Stebbins and Major (1965), and Raven and Axelrod (1978), discuss the evolution of the California flora in response to climate change. The evolution of social learning is not open to plants. Plants show a variety of responses to the onset of cooler, drier, summer drought conditions of the novel Mediterranean climate that accompanied the Pleistocene. Trees responded mainly by range changes of existing species, not by Darwinian evolution. As a result, California has a number of "paleoendemics," mostly trees, that once had a much wider distribution and are now restricted to narrow niches. Redwoods are the most famous example. Many of California's most widespread tree species, for example our evergreen oaks, evolved in the Sierra Madre mountains of Mexico and invaded California with the onset of the semiarid climate. Other elements of the vegetation, including many shrub and herb genera, have radiated under the new conditions to form flocks of "neoendemics." Wild lilac and lupine, represented by a few dozen closely related species, are examples. Such groups respond to changing environments with niche shifts via speciation. It might be thought that this pattern is the result of evolutionary conservatism on the part of long-lived species like trees and greater flexibility on the part of species with short generations. However, some patterns do not conform to this expectation. Beetle species from the early Pleistocene can almost all be referred to modem species (Coope, 1979), whereas mammals have been evolving quite rapidly, with an average species duration of only a million years or so (Stanley, 1979). No simple patterns of response to climate deterioration have so far emerged from studies of selected sets of species. As far as we are aware, no one has proposed that plants and beetles have responded to climatic deterioration by any special set of evolutionary mechanisms. Rather, the conservative species like trees and beetles have responded by dramatic niche-chasing range shifts as climate changes. Coope (1979) describes a species of beetle from a Pleistocene cold period fauna in Britain that was thought to be one of the few without a modem population until one was found in a remote location it Tibet. Clark et al. (1998) review evidence suggesting that large, long-lived trees like beeches and oaks can move surprisingly rapidly in response to climate change. Rates of 150-500 m/yr are necessary to account for rates of range expansion characteristic of the end of the last glaciation. Trees could thus respond even to events on the time scale of the Dansgaard-Oeschger events. The rapid to and fro movements of the large woody vegetation created Pleistocene forests with diverse, probably non-equilibrium, mixtures of species that do not normally co-occur today. Other kinds of populations seem simply to have gone extinct or undergone niche changing speciation in response to climate change. Elizabeth Vrba (1995) has proposed a bold hypothesis about the impact of Plio- Pleistocene climatic fluctuations on mammalian evolutionary patterns. She argues that the fluctuating climate should generate waves of extinctions and speciations. Consider a warm climate that is turning cold impacting a lineage that is generally cool adapted. In the warm climate, such a lineage will only persist as fragmented alpine relicts. Under such fragmentation, new species will arise, but many old ones will become extinct. As the climate cools again, formerly fragmented populations, now new species, will expand their ranges and become temporary dominants. Under very cold conditions, fragmentation will occur again, with a new wave of extinctions and an incipient radiation. DeMenocal and Bloemendal (1995; see also De Menocal, 1995) argue patterns of hominid evolution in the Plio-Pleistocene map onto the stepwise changes in the variability in the climate record shown in Figure 1. The evidence for Vrba's hypothesis is mixed (see other chapters in Vrba et al., 1995), but there is no doubt that ordinary Darwinian evolution can be rapid on the geological time scale (Carroll, 1997: Ch. 3). Thus, many lineages have apparently adapted to climate deterioration without any mechanisms outside the limits of conventional ecological succession (niche chasing), and/or adaptive phyletic evolution and speciation (niche shifting). Brain Size Evolution in the Neogene Mammals are the one group that shows clear signs of responding to variability selection with Potts' hypothesized increased behavioral flexibility. Harry Jerison's (1973) classic treatment of the evolution of brain size documents a major trend towards increasing brain size in many mammalian lineages that persists right up to the present. Figure 6 summarizes his data. The data are presented in the form of cumulative frequency distributions of encephalization quotients of carnivores and ungulates over the whole CenozoicEra. The sample includes: (1) archaic creodont carnivores (an extinct order), (2) archaic ungulates and carnivores from the extinct orders Condylartha and Amblypoda from the Paleogene (65-22.5 million yrs before present), (3) members of the still extant ungulate and carnivore orders from the Neogene (22.5-2.5 million years before present), and (4) a selection of living species of ungulates and carnivores. The time trends illustrated by the figure are complex. There is a progressive increase in average encephalization throughout the Cenozoic. However, there is an interesting tipping of the cumulative curves to the north east through time as well. Many relatively small-brained mammals persist to the present even in orders where some species have gotten rather large brains. The diversity of brain size increases toward the present. Mammals continue to be under strong selection pressure to minimize brain size, and those that find an effective way to cope with climatic deterioration by niche chasing or organic niche shifting do so. Other lineages evolve the means to exploit the temporal and spatial variability of the environment by using behavioral flexibility instead. These last, we suppose, pay for the cost of encephalization by exploiting the ephemeral niches that niche chasing and niche shift' species leave under-exploited. Humans merely anchor the tail of the recently much-stretched distribution of brain sizes in mammals. We are the largest brained member of the largest brained mammalian order, sharing our position on the frontier with a few cetaceans (porpoises). This is comforting to a Darwinian hypothesis. Large gaps between species are hard to account for by the processes of organic evolution. That we are part of a larger trend suggests that a large scale, general selective process such as we propose is really operating. Nevertheless, there is some evidence that human culture is more than just a more sophisticated form of typical animal social learning. More on this vexing issue below. Note that biggest shift per unit time by far is the shift from Neogene to modem species. In the 2.5 million years from the late Pliocene to the end of the Pleistocene, encephalization increases were somewhat larger than the steps from Archaic to Paleogene and Paleogene to Neogene, each of which represent tens of millions of years of evolution. Nevertheless, the Plio-Pleistocene leap in brain size is part of a trend that reaches back before the beginning of the Cenozoic. Since detailed records of climate variability are so far only published for the last 6-7 million years (Figure 1), it is not possible to say whether a record of climate deterioration accompanies the earlier increases in encephalization. Based upon the long-term trend of encephalization, our hypothesis predicts that climatic variability in the early and middle Cenozoic must have exceeded that of the Mesozoic when encephalization was still more modest. The degree of deterioration in these earlier episodes deterioration should have been less dramatic that the Pleistocene, but still appreciable. Further work on deep sea cores should provide a test as for back as there is oceanic crust to examine. Social Learning Versus Individual Learning? Increases in brain size could signal adaptation to variable environments via either individual or social learning. The mathematical models suggest that that the two systems work together. There should be an optimal balance dictated by the spatio-temporal structure of the variability selection imposed. Given the tight constraints imposed on brains, we would expect to find a tradeoff between social and individual learning abilities. Those species that exploit the most variable niches should emphasize individual learning while those that five in more highly autocorrelated environments should devote more of their nervous systems to social learning. Following this line of reasoning, Lefebvre and Giraldeau (1996) conducted experiments on two species of pigeons, one social and opportunistic and the other more conservative and less social. The social and opportunistic species, they reasoned, should be able to learn socially more easily than the more conservative species, and the conservative species should be better individual reamers. They also reviewed the data of other investigators comparing social and individual learning in a small sample of songbird species. Surprisingly, the prediction fails. Species that are good social reamers are also good individual reamers. Is this counter-intuitive finding a product of a very small, potentially misleading, sample? Perhaps not. It may be that individual and social learning are not strongly competing processes and might even be synergistic. Jerison (1973) argued that the expansion of the neocortex, which accounts for most of the tissue involved in encephalization trends, is devoted to "maps' of the environment. Animals with more detailed maps need to acquire the information to fill them out. Both social and individual learning will help do so. Perhaps the information evaluating neural circuits used in social and individual learning are also partly shared. Once animals become social the potential for social learning arises. If the marginal benefit of reducing the neural circuits unique to individual learning are modest, social species enlarging their maps to take advantage of social learning may come under selection to improve individual learning as well. If the two systems share the overhead of maintaining the memory storage system and much of the machinery for evaluating the results of experience, the marginal benefits in quality or rate of information gain may be large relative to the cost of more specialized nervous tissue. If members of the social group tend to be kin, investments in individual learning may also be favored because sharing the results by social learning will increase inclusive fitness. The hypothesis that the tradeoff between social and individual learning may be very marginal resonates with the mechanisms of social learning found in most wen-studied cases of social learning. Galef (1988, 1996), Laland et al. (1993), and Heyes and Dawson (1990) argue that the most common forms of social Teaming result from very simple mechanisms that piggyback on individual learning. In social species, naive animals follow more experienced parents, nestmates, or flock members as they traverse the environment. The experienced animals select highly non-random paths through the environment. They thus expose naive individuals to a highly selected set of stimuli that form the basis for acquisition of behaviors by ordinary mechanisms of reinforcement. Social experience acts, essentially, to speed up and make less random the individual Teaming process, requiring little additional, specialized, mental capacity. Social Teaming, by making individual Teaming more accurate without requiring much new neural machinery, tips the selective balance between the high cost of brain tissue and advantages of flexibility in favor of more flexibility. As the quality of information stored on a map increases, it makes sense to enlarge the scale of maps to take advantage of that fact. Eventually, diminishing returns to map accuracy will limit brain size. Eisenberg's (1981: Ch 23) review of a large set of data on the encephalization of living suggests that high encephalization is associated with longer times of association with parents, late sexual maturity, extreme iteroparity, and long potential life-span. These life cycle attributes all seem to favor social learning. It may be that large brains cannot be supported in the absence of the opportunity to learn socially from parents at least, and that investments in social Teaming must be amortized over a long life. Even marginally social species may come under selection for behaviors that enhance social Teaming, as in the well known case of mother housecats to bring partially disabled prey to their kittens for practice of killing behavior (Caro and Hauser, 1992). If the relationship between social and individual learning is as tight as this evidence suggests, then we can expect to find social learning in many if not most social species. Indeed, the best studied example of social Teaming is the food choice system of norway rats (Galef, 1996). This species, with an encephalization quotient of about 0.4 (0.8 using Eisenberg's, 1981:499, re-estimated allometry relationship), is among those that have participated only modestly in the Cenozoic encephalization trend (Jerison, 1973: 212, 218). Lefebvre and Palameta (1988) provide a long list of animals in which social learning has been more or less convincingly documented. Recently, Dugatkin (1996) and Laland and Williams (1997) have demonstrated social learning in guppies. The idea that brain size, social Teaming and individual Teaming are all tied to a rather generalized environment mapping system is contrary to the attractive, widely held, idea that brains are collections of highly specialized modules (Fodor, 1983). Tooby and Cosmides (1989) argue that modular specialization of brain function is to be expected on general theoretical grounds. Lefebvre and Giraldeau's result is perhaps more congenial to the connectionist hypothesis holding that much brain tissue functions as a rather generalized pattern recognition device. We have neither the space nor the competence to review this issue in detail. Tononi et al. (1994) note that, at the neurological level brains are complex just because they are at once extensively modular and richly integrated. Thus, individual Teaming capacities may be positively correlated with capacities for social teaming because they are mostly sub-served by the same modules and integrating circuits. Aspects of the social learning system in animals do show signs of adaptive specialization. For example, Terkel (1996) and Chou (1989, personal communication) obtained evidence from laboratory studies of black rats that the main mode of social learning is from mother to pups. This is quite unlike the situation in the case of norway rats, where Galef (1988, 1996) and coworkers have shown quite conclusively that mothers have no special influence on pups. In the black rat, socially learned behaviors seem to be fixed after a juvenile learning period, whereas norway rats continually update their diet preferences (the best-studied trait) based upon individually acquired and social cues. Black rats seem to be adapted to more slowly changing and norway rats more rapidly changing environments. Terkel studied a rat population that has adapted to open pinecones in an exotic pine plantation in Israel, a novel and short-lived niche by most standards, but one that will persist for many rat generations. Norway rats are the classic rats of garbage dumps, where the sorts of foods available change on a weekly basis. Interestingly, in recent decades, norway rats have been expanding at the expense of black rats (e.g. Bentley, 1964). Its seems possible that modem garbage dumps present a much more varied resource for rats than traditional ones, and that the spread of norway rats reflects their better adaptation to human modernity. The theory we have described suggests that selection on social reamers might well tune the social learning system to match the statistical properties of the environmental variation in the specific niche the animal occupies. This very thin bit of data suggests that the hypothesis is worth pursuing. In the human case, we have at least one highly specialized social learning systen-4 language. On the other hand, we readily learn to make a living using a spectacular array of techniques. As the famous language learnability argument of Chomsky shows, a completely general learning machine cannot work (Pinker, 1994: Ch 9). A finite learner must have a nervous system that in effect makes many assumptions about the environment in order even make the most basic map of its environment. For example, primates have a visual system imposes order on nerve impulses coming from the rods and cones to produce a fairly veridical image of objects in the world (Spelke, 1990). It win not do just to have a flood of impulses flowing from a large array of sensory cell. There must be built in expectations about what sorts of objects are out there to sense. For example, the visual system assumes that a set of spatially contiguous points in the visual field that have a similar color, defined border, and coherent movement is a solid object. This innate physics correctly recognizes a rolling ball as a solid object, though it misidentifies clouds as such. On the other hand, the adaptive reason to have learning and social learning is the flexibility to adapt to unforeseen contingencies. Experience teaches us that the solidity of clouds is an illusion. Perhaps the neocortex of the brain is an adaptation like the beak of birds. The basic beak is nothing more than a moderately complex, functionally integrated, forceps-like device with multiple functions. It is usually a food acquisition, handling, and processing organ, fighting weapon, environment probe, and grooming tool, all in one. Nevertheless, despite great commonality of form and function, bird beaks are endlessly stretched, bent, thickened, widened, deepened. and sharpened to support the diverse niches birds occupy. Only occasionally are new parts, like the pouch of pelicans, added to create a new adaptation. It strikes us that the modularity-connectionist debate does not exhaust the possible models for the relationship between form and function in brains. We wish to underline how little we know about the adaptive tradeoffs in brain design. Neurophysiologists, cognitive scientists, and behavioral ecologists each have something to contribute to the puzzle of how some species can support large brains. However, collaborations between these disciplines to tackle this question have been lacking. Human Versus Other Animals' Culture The human species position at the tail of the distribution of late Cenozoic encephalization admits of the hypothesis that our system of social learning is merely a hypertrophied version of a common animal system. However, two lines of evidence suggest that there is more to the story. First, human cultural traditions are often very complex. Subsistence systems, artistic productions, languages, and the like are so complex that no one individual ever could or did invent them. Rather, they are built up over many generations by the incremental, marginal modifications of many innovators (e.g. Basalla 1988). We are utterly dependent on learning such complex traditions to function normally. Think of some relatively simple item like a hunting spear. The maker has to know how to make the stone tools to prepare the shaft, how to knap fine-grained stone to make a good point, how to prepare stout adhesives and fiber to mount the point, what wood makes a spear of the right strength and weight to be useful how long and stout to make the shaft for the intended purpose (throwing, stabbing, multipurpose), and so forth. Few of us could make more than a crude approximation of a (late) Stone Age spear, though we could easily learn by being taught, or even just by observing an expert spear-maker. Most animal culture appears to be much simpler. Terkel's study of the manner in which it roof rats open pine cones showed that individuals cannot normally learn to open a pine cone in a way that leads to a net energy gain. However, there is only one trick involved in the successful technique, and it is likely that a single individual innovated the trait in the beginning. This difference in the complexity of socially learned behaviors is mirrored in a major difference in mode of social learning. As we saw above, much social learning seems to be dependent mostly on the same techniques used in individual learning. Experimental psychologists have devoted much effort to trying to settle the question of whether nonhuman animals can learn by "true imitation" or not (Galef, 1988). True imitation is learning a behavior by seeing it done. True imitation is presumably more complex cognitively that merely using conspecifics' behavior as a source of cues to stimuli that it might be interesting to experience. Although there are some rather good experiments indicating some capacity for true imitation in many socially learning species (Heyes, 1994; Zentall, 1996; Moore, 1996), head-to-head comparisons of children's and chimpanzee's abilities to imitate show that children begin to exceed chimpanzees' capabilities at about 3 years of age (Whiten and Custance, 1996; Tomasello, 1996). There is still considerable doubt about the significance of imitation in wild chimpanzees. Tomasello is inclined to think that even in this species, there is remarkably little indication that apes can ape, although human reared chimpanzees do show considerably imitation. Whiten and Custance, on the other hand, argue that the marginal abilities observed under impoverished captive conditions are likely to underestimate abilities in the wild. McGrew (1992) reviews the evidence from chimpanzee material culture suggesting that chimpanzee tools are as complex as the simplest know toolkits of humans. Perhaps some chimpanzee tools, especially the hammer-and-anvil nut cracking system found in some West African populations, is a cumulative tradition that is made up of at least two independent inventions. Rehabilitating pet orangutans exhibit very impressive imitations of human behavioral routines, though they are not known to have any sign of imitatively acquired behaviors in the wild (Russon and Galdikas, 1993). Parrots seem to have acquired a quite respectable but little understood capacity for imitation (Moore, 1996). On the other hand, monkeys show scant signs of abilities to imitate, even Capuchin monkeys, which have a higher encephalization quotient than even the Great Apes (Fragaszy and Visalberghi, 1996; Eisenberg, 1981, 499). Thus, the lesson to date from comparative studies of social learning suggests that very simple mechanisms of social enhancement of cues is much more common and more important than imitation, even in our close relatives and other highly encephalized species. It may be that the ability to imitate others freely requires that individuals have a theory of mind (Premak and Woodruff, 1978; Cheney and Seyfarth, 1990: Ch. 8). Without the ability to model the intentions of other individuals, it may be difficult to translate observations of another's acts into subjective terms so that one can replicate the act. As a consequence of an inability to "see" the elements that go to make up complex behaviors when they are performed by others, most social reamers cannot imitate. They must relearn most elements of the behavior for themselves, with the social part of learning restricted to being exposed to the stimuli that tend to elicit the correct behavior in the end. Thus the complexity of proto-cultural traditions is much less than in the case of humans. Even rather encephalized animals such as the monkeys, seem to use the evaluative machinery that is used in individual learning to evaluate socially supplied cues, but to have invested little specialized machinery to imitate. Although we expect that social learning systems in non-human animals will prove to be common and varied in nature, the human ability to evolve complex cultural traditions appears to be unique, or at least uniquely hypertrophied. This generates the basic problem of accounting for human minds. Humans have apparently penetrated a "cognitive niche" (Tooby and DeVore, 1987) which made us a rather successful species under Pleistocene conditions. Using complex traditions, we successfully occupied niches from the tropics to the glaciers, penetrating to all but the harshest environments (Klein, 1989). In the Holocene, the development of food plant production has made us the Earth's dominant organism. If human traditional culture is a successful adaptation for us, why haven't other species evolved similar capacities? There are a number of possible reasons. It could be that we have simply won an evolutionary footrace to be the first animal to occupy the cognitive niche. The final evolutionary innovations that permitted complex traditions were rather late. The Upper Paleolithic Transition in Europe, the settlement of Australia (requiring tolerably sophisticated boats), and other signs of the final modernization of human cognition occur within the last 100,000 thousand years (Stringer and Gamble, 1993). It might be that upright posture, freeing the hands to make and carry artifacts, was necessary to make complex traditions useful. Our species may have been the only one with this or some other non-cognitive preadaptation necessary to permit the evolution a capacity for complex culture. Why Is Complex Culture Rare? There is also a potential evolutionary impediment to the evolution of a capacity for complex traditions. We show elsewhere that, under some sensible assumptions, a capacity for complex cumulative culture cannot increase when rare (Boyd and Richerson, 1996). The mathematical result is quite intuitive. Suppose that to acquire a complex tradition efficient imitation is required. Suppose that efficient imitation requires considerable costly, or complex, cognitive machinery, such as a theory-of-mind/imitation module. In such a case, there will be a coevolutionary failure of capacity for complex traditions to evolve. The capacity is a great fitness advantage, but only if there are cultural traditions to take advantage of But, obviously, there cannot be complex traditions without the cognitive machinery necessary to support them. A rare individual who a mutation coding for an enlarged capacity to imitate will find no complex traditions to learn, and will be handicapped by an investment in nervous tissue that cannot function. The hypothesis depends upon there being a certain lumpiness in the evolution of the mind. If even a small amount of imitation requires an expensive or complex bit of mental machinery, or if the initial step in the evolution of complex traits does not result in particularly useful ones, then there will be no smooth evolutionary path from simple social learning to complex culture. If such an impediment to the evolution of complex traditions existed, evolution must have traveled a round-about path to achieve get the frequency of the capacity high enough to begin to bring it under positive selection for its tradition-supporting function. Some have suggested that primate intelligence was originally an adaptation to manage a complex social fife (Humphrey, 1976; Whiten and Byrne, 1988, Kummer et al., 1997). Perhaps in our lineage, the complexities of managing the sexual division of labor or some similar social problem favored the evolution of a theory-of-mind capacity. Such a capacity might then incidentally make efficient imitation possible, launching the evolution of complex traditions that could drive the evolution of still more sophisticated imitation. This sort of stickiness in the evolutionary processes is presumably what gives evolution its commonly contingent, historical character (Boyd and Richerson, 1992). There is some evidence supporting this hypothesis. Even among cognitively modem humans, the maintenance of complex traditions is not unproblematic. The Tasmanian toolkit shrank in size and sophistication after their isolation from Australia by the Holocene rise in sea level. Diamond (1978) argues that this may have resulted from a sort of cultural drift. In a small population, complex skills will occasionally be lost by accident. With few people to reinvent them and no possibility of reacquiring them by diffusion, the Tasmanians were helpless to prevent an erosion of their more complex cultural traditions. Even given a capacity for complex traditions, the number of participants in a cultural system may be critical to the complexity that can be maintained. A few rare, unsophisticated imitators would presumably have a much harder time launching or maintaining a complex tradition than a larger group that can pool many piecemeal innovations. Diamond (1997) expanded this hypothesis to the continental scale. He argues that Eurasian cultural sophistication grew more rapidly than traditions on other continents because Eurasia was the largest, most populous continent, and because innovations could readily spread east and west in similar ecological zones. The Americas are not only smaller continents, but are relatively narrow and oriented on a north-south axis. Innovations will tend to spread more slowly across than along ecological zones. It is interesting that the manufacture of complex compound tools like a stone-tipped spear appears quite late in the evolution of hominids. Such tools are associated with other signs of a more complex imitative mental life, such as art and stylistic variation in utilitarian objects (Klein, 1989: 369, 379-83; Donald, 1991). Late archaic hominid populations, such as Neanderthal peoples from Europe, had a much less complex technology than the anatomical modems that followed them, despite having slightly larger brains (Stringer and Gamble, 1993). Perhaps McGrew is correct that chimpanzee tool kits nearly reach the complexity of the simplest example among modem humans. Perhaps Tomasello is correct that imitation is an ancient latent potential of ape cognition that is manifest in chimpanzee and orang behavior given human models. Perhaps most or all hominid brain enlargement throughout the Pleistocene was largely the product of selection forces other than those exerted by the opportunities afforded by complex traditions. Perhaps hominids as modem as Neanderthals stir used their large brains largely for individual learning and the correlated ability to learn socially using the simple stimulus enhancement mechanisms quite widely used in social animals, not imitation. There is no evidence that the past 100,000 years of the Pleistocene environment has deteriorated significantly since the shift from dominance of the 41,000 year Milankovitch cycle to tie dominance of the 98,500 cycle about a million years ago. The ice core data do not go back beyond the last interglacial (oxygen isotope stage 5). Thus, it is not possible to say if phenomena such as the Dansgaard-Oeschger events are restricted to the last glacial or whether they are more ancient. Records from old African lakes may one day test directly whether hominid evolutionary events of the last glacial cycle could have been driven directly by climate or not. On present evidence, a cognitive niche for a sophisticated, imitative social learner would seem to have existed for at least a million years before the adaptation appeared. Perhaps human cognition came under direct selection for the capacity to imitate only within the last 100,000 years or so as some preadapted populations began to develop complex traditions, finessing the maladaptive-when-rare problem. The anomalously old, complex, Howieson's Poort artifacts (Klein, 1989: 308), and the recent discovery of a sophisticated bone points dating back 90,000 years in Central Africa (Brooks et al. 1995), suggest that some populations in Africa began creating complex traditions as early as the last interglacial. Anatomically modem populations also date back to perhaps 90,000 years ago. However, in the Near East, anatomical modems are clearly associated with artifacts identical to those made by Neanderthal populations that sometimes succeeded them in the same and nearby sites (Klein, 1989: 303-305). The critical evolutionary steps to the imitative capacity must have occurred in a sub-population of anatomical modems, probably living somewhere in Africa something like 90,000-50,000 years ago. The latest Neanderthal Chatelperronian industry in Europe does show signs of the diffusion of complex elements from anatomically modem populations that were just entering Europe (Klein, 1989: 335-336). Neanderthals seem capable of acquiring traditions more complex than they ever produced without such a stimulus. Much as with apes, they seem to have had a cryptic capacity for imitation larger than they routinely used. One gets the impression that apes and archaic hominids are near and yet so far from the imitative/complex tradition adaptation during the whole Pleistocene Epoch. It is even possible that there has been no significant coevolution of human minds for imitative capacities at all. At the very least, the paleoanthropological evidence rather strongly suggests that nearly modem human minds evolved by preadaptation while making artifacts that are much less sophisticated than those made after some sort of Upper Paleolithic "revolution." At most, the period of coevolution of human minds to reach the full range of complex traditions would appear to be bracketed by the slightly modernized Howiesons Poort industry after 100,000 thousand years ago and the intrusion of anatomically modern humans making the Aurignacian industry into Europe 35-40,000 years ago (Klein, 1989: Ch.6-7). On the present evidence, the character of climate variability has been unchanged since I million years ago. Thus, it is tempting to think that Homo erectus and archaic Homo sapiens spent several hundred thousand years adapted to living under conditions of extreme environmental fluctuation, using a system of relatively simple technological adaptations represented by the Acheulean stone tool tradition. This tool tradition and its close relatives occur from about 1.5 million years ago in Africa to about 200,000 years ago (Klein, 1989: Ch.4). Indeed, considerable brain enlargement occurred in the transition from Homo erectus to archaic H. sapiens, without any evidence of change in the archeological assemblage at all (Klein, 1989: Ch. 5). It is hard to understand from the paleoanthropological record as it stands how archaic hominids were paying the costs ofbrain enlargement, as conservative as technology apparently was. Relatively lately and relatively suddenly, culture became much more complex, as if some final preadaptive breakthrough permitted the emergence imitation. To judge from brain size, the main reliable clue, the advanced cognitive capacity preceded the first complex traditions by a few tens of thousands of years at least. Perhaps there was a final cognitive modernization not reflected in brain size under the influence of coevolution with complex traditions. It would seem surprising if this were not so, but the evidence is unimpressive. It does seem to be the case that the encoding and decoding of speech occurs at such a rapid rate that specialized structures must be involved (Friederici, 1996). The gross anatomy of the vocal tract of modem humans also seems rather clearly to be specialized for speech (Lieberman, 1984). The extant data at least permit the hypothesis that the ability to acquire complex traditions by imitation is wholly a preadaptation, and encourage the hypothesis that it is largely so. Donald's (1991) admirably well-specified scenario for the origins of human cultural complexity involves a stage of sophisticated motor mimicry preceding and laying the neurological basis for language. It is plausible, as Donald shows, that quite complex behavior can be acquired by mimicry in the absence of language. 19th Century accounts of the abilities of deaf-mutes to acquire many sorts of useful economic and social skills without language suggests that they could easily learn to make a serviceable stone-tipped spear by observation, without any linguistic aids. However, since major increases in tool complexity postdate anatomically modern skulls, it seems possible that this argument needs to be reversed to account for the final origin of complex traditions. Perhaps Achuelean and Mousterian level tools were made with only minimal imitative capacities. On the one hand, Acheulean tools show rather stereotyped features suggesting a more sophisticated form of social learning that mere stimulus enhancement. On the other, the stereotyped pattern remains very similar across large geographic distance and great spans of time. There is no indication that stone tool traditions of the middle Pleistocene were capable of sensitive adaptation to local environment. It is as if the "cultural" patterning of Acheulean artifacts was in fact innate. Thus, if anatomically modem skulls signal the development of modem linguistic capacities, that change would be roughly contemporaneous with the modest shift in tool complexity from the Acheulean to the Mousterian industry. Some Mousterian populations (anatomical modems) evolved the capacity for complex language systems, while others (Neanderthals) did not. The Upper Paleolithic artifact revolution in this scenario would follow language, not antedate it. The key step might have been the evolution of language to manage some aspect of social behavior, for example the emergence of a stable division of labor. It would presumably require complex negotiations between men and women to manage such a social system. Ditto for the multi-family cooperative, risk-sharing hunting and gathering band of ethnographic fame. It is plausible that small steps in the increase of communication complexity would allow the coevolution of a language capacity and a cultural linguistic system. The evolution of language might not have the evolutionary impediment to the evolution of complexity, at least not so severe a one. We are used to thinking that language is the ultimate human cultural achievement. The evidence that much of the complexity of language is innate (Pinker, 1994) suggests that this system is actually more primitive than other systems that appear to have a much smaller innate component, such as post-Acheulean technology. If children have enough innate information to largely create a new language in one generation (Bickerton, 1984), this system may not really depend upon the existence of complex traditions, although it produces them as a byproduct. The linguistic system perhaps made the first complex technical traditions possible by making it easy to express and memorize cultural principles verbally. In this way, a culture which initially become complex using the oral mode of transmission may have selected for more facile mimetic capabilities for those common learning tasks where a picture is worth a thousand words. Whatever the exact sequence of events, there is independent evidence from human paleodemography that supports the idea that some sort of rare evolutionary accident was necessary to achieve a functional capacity for complex cultural traditions. Rogers and Harpending (1992) argue that human mitochondrial DNA records evidence of a human population explosion between 33,000 and 150,000 years ago. In small populations, genetic drift limits the diversity of mitochondrial genomes in the population. If a small population suddenly expands, it will begin to preserve more variation. The age of diversification of mitochondrial DNA lineages can be estimated from the number of mismatches separating different lineages. If a population suddenly expands, a large number of new mitochondrial lineages will arise more or simultaneously and will be preserved against loss by drift. As these lineages accumulate more mismatches over time they generate a clock that allows us to estimate the time of the original population explosion. Although confidence intervals are wide, the data suggest that the population ancestral to all modem humans was quite small until something like 60,000 years ago, when it expanded suddenly by a factor of more than 100 fold. The pre-expansion population was between 1,000 and 7,000 breeding females. At the time of the final modernization of the human mind, we were most likely a rare and, given the nature of the Pleistocene, endangered species. At our point of origin, our numbers were probably much smaller than those of the archaic populations that we replaced when our population exploded. The date of the explosion is consistent with the dates of the appearance of more sophisticated tools, though so far only the aforementioned replacement of Neanderthals by modem Homo sapiens in Europe around 35,000 years ago is well document. Exactly what quirk of the evolutionary process led to the final modernization is likely to prove very difficult to recover. Our rarity at the crucial time does suggest that only after the capacity for complex culture was in place, and complex traditions in fact began to evolve, did the main adaptive advantage of the cognitive change become apparent. The crucial cognitive innovations leading to the capacity for complex culture would seem to have occurred in a small population that was barely paying the overhead for their large brain. It is easy to imagine such a population winking out in under the severe, variable conditions of the last Glacial before the complex culture innovation was fully functional. If it had, who knows how many more glacial cycles would have passed with big-brained hominids poised on the threshold of the complex culture adaptation without crossing it. It is thus quite plausible that the transition to complex culture required crossing an evolutionary divide. It is also plausible that the crossing was a result of a long history of preadaptation rather than directly adaptation for complex traditions. How else are we to account for the near stasis of material culture (or protoculture) during long spans of time when brain enlargement was significant? Humans in the HoloceneA final issue concerns us. Since the climate of the Holocene has been unusually tranquil, why have not humans and other animals begun to re-evolve smaller brains? Kurt Vonnegut in his novel Galapagos invites us to imagine a time in the future when the troublesomely large human brain has shrunk considerably. Several possible reasons come to mind. Perhaps there hasn't been enough time for the shrinkage process to proceed appreciably. Domestic animals under artificial selection, however, have considerably smaller brains, scaled for body weight, than their wild progenitors. A few thousand years is certainly sufficient time for shrinkage to occur if selection is strong. Perhaps the modest fluctuations of the Holocene are at the right time scale to maintain selection for encephalization. As Lamb (1977) describes, the Holocene fluctuations are troublesome enough. The Pleistocene fluctuations are much more spectacular to be sure, but perhaps they are at sufficiently long time scales as to be mainly irrelevant. The most promising hypothesis we think is that humans themselves have become a major source of perturbation in the Holocene. The development of food plant production set off a demographic revolution in many parts of the world. Even in areas of the world like Western North America where participation in agriculture was rare, subsistence and social organizational innovations led to considerable human population increase and movement (Bettinger and Baumhoff, 1982). In essence, the end of the Pleistocene ushered in an intense competitive arms race between human populations where the winners were typically those with the more sophisticated traditions. The penetration of humans to virtually all of the World's habitats ensures that other mammals have an especially wily predator and competitor to deal with. Human dynamism in the Holocene seems easily sufficient to maintain selection for complex cognitive capacities in ourselves and other mammals. Should the quite Holocene climate persist for long enough, it is interesting to contemplate whether Vonnegut's scenario would obtain in the end. Conclusion Humans are a difficult species. The Darwinian project is committed to bringing us into the same basic explanatory framework as all other organisms. In his M notebook on August 16 1838, while he was in full cry in pursuit of his first formulation of natural selection, Darwin wrote "Origin of man now proved.--Metaphysics must flourish.-He who understand baboon would do more toward metaphysics than Locke" (Barrett, 1974: 281). Darwin realized that failing to account for human behavior left a dangerous gap in his theory through which critics could and did try to attack his whole theory (Richerson and Boyd, in press). When, in the Descent of Man, Darwin reluctantly undertook to explain our species, a hostile critic in the Quarterly Review, whom Darwin believed to be his persistent critic St. George Mivart, took advantage of perceived problems in the human story to attack the whole ediface. He gloated, it "offers a good opportunity for reviewing his whole position7' (and rejecting it, Anonymous, 197 1). If the gap between humans and our animal ancestors is too large, it is difficult to explain how ordinary evolutionary processes could explain our origins. If one species escapes the net of evolution by natural selection, the whole of so comprehensive a theory is brought into question. On the other hand, if there is not some "great gap" between our ape ancestors and ourselves it is difficult to account for our runaway success. Our nearest living ancestors are forest dwelling creatures with modest ranges and abundances. Our more immediate fossil ancestors began to extend their ranges beyond Africa a million or so years ago. Even then, the range of Homo erectus grade hominids was not strikingly greater than that of some large carnivores like lions and pumas. However, by latest Paleolithic times, Homo sapiens sapiens, using toolkits of great sophistication, penetrated to the furthermost reaches of the Old World. With the waning of the ice at the end of the Pleistocene we exploded into the New World. In the Holocene, human societies have become very complex, rivaling in this regard the eusocial insects. In the early 20th Century, the Mendelian revolution eventually ended biologists' interest in the inheritance of acquired variation. The emerging social sciences sought disciplinary autonomy from biology (and from each other). As a result, for many decades only a few scattered specialists and individual thinkers worked on humans in the framework outlined by Darwin (Campbell, 1965, 1975). The important works on the topic in mid-century, for example Dobzhansky (1962: 18-22), in his well respected Mankind Evolving, faced an unbridgeable disciplinary gap. In the short theoretical section of his book cited above, he acknowledged the autonomy claims of the social sciences with statements like "In producing the genetic basis for culture, biological evolution has transcended itself-it has produced the superorganic." However, in adjacent passages he hewed to his Darwinian roots: "The fact which must be stressed, because it has been missed or misrepresented, is that the biological and cultural evolutions are parts of the same natural process." In 1962 there was neither the theory nor the empirical work to make sense out of these seemingly contradictory statements. The scientific community was content to five with such ambiguities, a few controversy provoking figures like Konrad Lorenz aside. In the late 20th Century, evolutionary biologists became much more interested the evolution of behavior. W.D. Hamilton's (1964) famous papers on inclusive fitness launched the theoretical study of social evolution, and a growing body of empirical ethologists began conducting theoretically relevant investigations. Richard Alexander's (1974) review paper and Edward Wilson's (1975) treatise Sociobiology left no doubt that evolutionary biologists were going to apply this theory to humans with little or no respect for claims of disciplinary autonomy based on superorganismic claims for cultural processes. At the same time, the population geneticists Lucca Cavalli-Sforza and Marcus Feldman (1973) initiated the study of mathematical models of cultural evolution. Other threads important to the problem of understanding humans in a comparative framework were picked up in that period. For example, Bennett Galef (1977) began his important work on social learning in norway rats, and the first deep-ocean cores began to reveal the true dimensions of late Cenozoic climate deterioration. Advances in paleoanthropology in recent years have likewise been spectacular. After 25 years of relatively intensive work, there are still many important questions outstanding. This essay is an attempt to fink the results from theoretical modeling the evolutionary properties of culture with the main relevant empirical work to produce a synthetic hypothesis to explain the origin of human culture. We make no strong c@ for its particular postulates. Our frequent repetition of the word "perhaps" and its cognates emphasizes our belief that many important questions remain to be answered. We believe much more strongly that our scenario has the right general ingredients to be a successful explanation. It takes a stab at addressing the adaptive economics of large brains and connecting these considerations to the environmental changes that must have driven the encephalization trends of the Cenozoic. It takes a stab at explaining how humans, so lately derived from ancestors with relatively modest systems of social learning, could at once be spectacularly successful using culture but yet not have a crowd of competitors for the complex-culture cognitive niche. The hypothesis makes at least a few predictions that are testable. If it is true, the pattern of climatic deterioration of the Cenozoic should predict the pattern of mammalian encephalization. The correlation is good back 6 million years, and it appears that the stepwise climate deterioration is mirrored in events in hominid evolution (deMenocal and Bloemendal, 1995). It should also hold for the more modest brain size increases of the middle and early Cenozoic. If there is a problem getting the evolution of a capacity for complex culture started, we should expect that apes will show pre-adaptations for culture. That is, they should have cognitive capacities that are homologous to those that we use to support complex culture, but which are subsidized, if not entirely supported by, other functions. Chimps and orangs have some capacity for imitation that is apparently little if at all used in the wild. What function does it serve? Similarly, if parrots and other animals do prove to have capacities for imitation that might support complex cultural traditions, they should serve other functions. There are a number of important questions about which information is currently scanty. The evolutionary cognitive economics of brains is, despite the cognitive revolution, poorly understood. Is it really true that the incremental cost of improving simple social learning at a given level of individual learning is small? Is it true that even relatively rudimentary imitation requires costly cognitive machinery, making it impossible for selection to favor true imitation incrementally, beginning with rather simple traditions and working smoothly towards complexity on the human scale? Is it true that capacities for learning and social learning are closely tuned to environmental variation? The empirical support for our hypothesis comes from climate proxy data that are very far from measuring variations relevant to the fitness of particular organisms. Only the beautifully detailed ice core data permits us to look at variation on the generational time scale. Beyond about I 00,000 years ago, the ice core record disappears and we have only the coarser scale of deep-sea cores. Much more will be revealed in the future, for example from the very long ice core currently being drilled in Antarctica, or from cores from deep, old lakes like Lake Tanganyika. The challenge of the origin of the human mind is the same as it was in Darwin's day. Humans must have evolved by the same basic processes as other organisms yet we are highly unusual in our mode of adaptation by cultural traditions and in our ecological success. Quite likely, the climatic deterioration of the late Cenozoic, especially of the Pleistocene, played a key role in the evolution of culture. Capacities for social learning expanded as a means of adapting to the highly variable environment of the ice age, probably in many mammalian lineages. However, only our species went on to evolve the capacity to acquire complex cultural traditions by imitative social learning. The events leading up to the Upper Paleolithic revolution in cultural complexity remain obscure, but the preadaptive breakthrough hypothesis certainly cannot be ruled out. Literature Cited Aiello, L.C., and P. Wheeler. 1995. The expensive tissue hypothesis: The brain and the digestive system in human and primate evolution. Current Anthropology 36: 199-221.Alexander, R.D. 1979. Darwinism and Human Affairs. Seattle: University of Washington Press. Anonymous. 1871. Review of the Descent of Man and Selection in Relation to Sex by Charles Darwin. The Quarterly Review 131 (216):47-90. Barrett, P.H. 1974. Darwin's Early and Unpublished Notebooks. In H. E. Gruber, Darwin on Man. New York: Dutton. Pp. 259-426. Barth, F. 1969. Introduction. In: F. Barth, ed., Ethnic Groups and Boundaries. Boston: Little, Brown. Basall, G. 1 98 8. The Evolution of Technology. Cambridge: Cambridge University Press. Bentley, E. W. (1964). A further loss of ground by Rattus rattus L. in the United Kingdom during 1956-61. Journal of Animal Ecology 33: 371-3. Bettinger, R.L. 1991. Hunter-Gatherers: Archaeological and Evolutionary Theory. New York: Plenum. Bettinger, R.L. and M.A. Baumhoff. 1982. The Numic spread: Great Basin cultures in competition. American Antiquity 47: 485-503. Bickerton, D. 1984. The language bioprogram hypothesis. Behavioral and Brain Sciences 7: 173-221. Boyd, R. and P.J. Richerson. 1996. Why culture is common, but cultural evolution is rare. Proc. Brit. Acad. 88: 77-93. Boyd R. and P.J. Richerson. 1995. Why does culture increase human adaptability'.? Ethol. Sociobiol. 16: 125-143.Boyd, R, and P.J. Richerson. 1992. How microevolutionary processes give use to history. In: M.H. and D.V. Nitecki, eds., History and Evolution. Albany: The State University of New York Press.Boyd, R. and P.J. Richerson. 1989. Social learning as an adaptation. Lectures on Mathematics 'm the Life Sciences 20: 1-26. Boyd, R. and P.J. Richerson, 1987. The evolution of ethnic markers. Cultural Anthropology 2: 65-79. Boyd, R. and P.J. Richerson. 1985. Culture and the Evolutionary Process. Chicago: Chicago University Press. Broecker, W.S. 1997. Thermohaline circulation, the Achilles heel of our climate system: Will man-made C02 upset the current balance? Science 278: 1582-1588.Broecker, W.S. 1992. Climate cycles: Upset for Milankovitch hypothesis. Nature 359: 779-780. Broecker, W. S. and G.H. Denton. .1 990. What drives glacial cycles? Scientific American, January, pp. 49-56. Broecker, W.S., D.M. Peteet, and D. Rind. 1985. Nature 315: 21-25. Bronowski, J. 1973. The Ascent of Man. Boston: Little, Brown. Brooks, A.S., D.M. Helgren, J.S. Cramer, A. Fr W. Hornyak, J.M. Keating, R.G. Klein, W.J. Rink, H. Schwarcz, J.N. Lieth Smith, K. Stewart, N.E. Todd, J. Verniers, and J.E. Yellen. 1995. Dating and context of three Middle Stone Age sites with bone points in the Upper Semliki Valley, Zaire. Science 268: 548-553. Brownlee, D.E. 1995. Interplanetary dust: A driver of glaciation cycles? Nature 378: 558. Campbell, D.T. 1975. On the conflicts between biological and social evolution and between psychology and moral tradition. Amer. Psychol. 30: 1103-1126. Campbell, D.T. 1965. Variation and selective retention in sociocultural evolution. In H.R. Barringer, G.I. Blanksten, and R.W. Mack, eds., Social-Change in Developing Areas: A Reinterpretation of Evolutionary Theory. Cambridge, Mass.: Schenkman. Caro, T, and M. Hauser. 1992. Is there teaching in nonhuman animals? Quart. Rev. Biol. 67:151-174. Carroll, R.L.1997.Pattems and Processes of Vertebrate Evolution. Cambridge: Cambridge University Press. Cavalli-Sforza, L.L. and M.W. Feldman. 1973. Models for cultural inheritance. 1. Group mean and within group variation. Theoret. Pop. Biol. 4: 42-55. Cheney, D.L., and R.M. Seyfarth. 1990. How Monkeys See the World: Inside the Mind of Another Species. Chicago: University of Chicago Press. Chou, L-S. 1989. Social Transmission of Food Selection by Rats. Ph.D. Dissertation, University of California-Davis.Clark, J.S., C. Fastie, G. Hurtt, S.T. Jackson, C. Johnson, G.A. King, M. Lewis, J. Lynch, S. Pacala, C. Prentice, E.W. Schupp, T. Webb 111, P. Wyckoff. 1998. Reid's paradox of rapid plant migration: Dispersal theory and interpretation of paleoecological records. BioScience 48: 13-24.Coope, G.R.1979. Late Cenozoic fossil Coleoptera: Evolution, biogeography, and Ecology. Ann. Rev. Ecol. Syst. 10: 247-67.Dawson, A.G. 1992. Ice Age Earth: Late Quaternary Geology and Climate. London: Routledge.deMenocal, P.B. 1995. Plio-Pleistocene African Climate. Science 270: 53-59. deMenocal, P.B., and J. Bloemendal. 1995. Plio-Pleistocene climatic variability in subtropical Africa and the paleoenvironment of hominid evolution: A combined datamodel approach. In: E. S. Vrba, G. H. Denton, T. C. Partridge, L. H. Burckle, eds., Paleoclimate and Evolution With Emphasis on Human Origins. New Haven: Yale University Press. Pp. 262-298. Diamond, J. 1997. Guns, Germs, and Steel: The Fates of Human Societies. New York: Norton. Diamond, J. 1978. The Tasmanians: The longest isolation, the simplest technology. Nature 273: 185-6. Ditlevsen, P.D., H. Svensmark, and S. Johnsen. 1996. Contrasting atmospheric and climate dynamics of the last-glacial and Holocene periods. Nature 379: 810-812.Dobzhansky, T. 1962. Mankind Evolving: The Evolution of the Human Species. New Haven: Yale University Press. Donald, M. 1991. Origins of the Modem Mind: Three Stages in the Evolution of Culture and Cognition. Cambridge, Mass., Harvard University Press. Dugatkin, L.A. 1996. Copying and mate choice. In: C.M. Heyes and B.G. Galef, Jr., eds., Social Learning in Animals: The Roots of Culture. San Diego: Academic Press. Pp. 85-105. Eisenberg, J.F. 1981. The Mammalian Radiations: An Analysis of Trends in Evolution, Adaptation, and Behavior. Chicago: University of Chicago Press. Friederici, A.D. 1996. The temporal organization of language: Developmental and neuropsychological aspects. In: B.M. Velichkovsky and D.M. Rumbaugh, eds., Communicating Meaning: The Evolution and Development of Language. Mahwah, New Jersey: Lawrence Erlbaum. Fodor, J.A. 1983. The Modularity of the Mind: An Essay on Faculty Psychology. Cambridge Mass: NUT Press. Foley, R. 1987. Another Unique Species: Patterns in Human Evolutionary Ecology. London: Academic Press. Fragaszy, D. and Visalberghi. 1996. Social learning in monkeys: Primate "primacy" reconsidered. In: C.M. Heyes and B.G. Galef, Jr., eds., Social Learning in Animals: The Roots of Culture. San Diego: Academic Press. Pp. 65-84. Galef, Jr., 1996. Social enhancement of food preferences in norway rats: A brief review. In: C.M. Heyes and B.G. Galef, Jr., eds., Social Learning in Animals: The Roots of Culture. San Diego: Academic Press. Pp. 49-64. Galef, Jr. B.G. 1988. Imitation in animals: History, definition, and interpretation of data from the psychological laboratory. In: T.R. Zentall and B.G. Galef, Jr., eds., Social Learning: Psychological and Biological Perspectives. Hillsdale, New Jersey: Lawrence Erlbaum. Pp. 3-28. Galef, Jr., B.G. 1977. Mechanisms for the social transmission of food preferences from adult to weanling rats. In: L.M. Barker, M. Best, and M. Domjan, eds., Learning Mechanisms in Food Selection. Waco, Texas: Baylor University Press. Pp. 123-150. GRIP (Greenland Ice-core Project Members). 1993. Climate instability during the last interglacial period recorded in the GRIP ice core. Nature 364: 203-207. Grootes, P.M., M. Stuiver, J.W.C. Climate, S. Johnsen, and J. Jouzel. 1993. Comparison of oxygen isotope records from the GISP2 and GRIP Greenland ice cores. Nature 366: 552-554. Hamilton, W.D. 1964. The genetical theory of social behavior, 1, 11. J. Theoret. Biol. 7: 1 52.Heyes, C.M., and G.R. Dawson. 1990. A demonstration of observational learning using a bidirectional control. Quart. J. Exper. Psychol. 42B: 59-71.Humphrey, N.K. 1976. The social function of intellect. In: P.P.G. Bateson and R.A. Hinde, eds., Growing Points in Ethology. Cambridge: Cambridge University Press. Pp. 303-317. Jerison, H.J. 1973. Evolution of the Brain and Intelligence. New York: Academic Press. Klein, R- G. 1989. The Human Career: Human Biological and Cultural Origins. Chicago: University of Chicago Press. Kummer, H., L. Daston, G. Gigerenzer, and J.B. Silk. 1997. The social intelligence hypothesis. In: P. Weingart, S.D. Mitchell P.J. Richerson, and S. Maasen, eds., Human by Nature: Between Biology and the Social Sciences. Mahwah, New Jersey. Pp. 157-79. Laland, K.N., P.J. Richerson, and R. Boyd. 1993. Animal social learning: Toward a new theoretical approach. Perspectives in Ethology 10: 249-77. Laland, K.N. and K. W@. 1997 Shoaling generates social learning of foraginginformation in guppies. Animal Behaviour 53: 1161-1169. Lamb, H.H. 1977. Climatic History and the Future. Princeton: Princeton University Press. Landau, M. 1984. Human evolution as narrative. American Scientist 72: 262-68. Lefebvre, L. and B. Palameta. 1988. Mechanisms, ecology, and population diffusion of socially learned, food finding behavior in feral pigeons. In: T.R. Zentall and B.G. Galef, Jr., eds., Social Learning: Psychological and Biological Perspectives. Hillsdale, New Jersey: Lawrence Erlbaum. Pp. 141-64. Lefebvre, L. and L-A Giraldeau. 1996. Is social learning an adaptive specialization? In: C.M. Heyes and B.G. Galef, Jr., eds., Social Learning in Animals: The Roots of Culture. San Diego: Academic Press. Pp. 107-28. Lehman, S. 1993. Climate change: Ice sheets, wayward winds and sea change. Nature 365: 108-9. Lieberman, P. 1984. The Biology and Evolution of Language. Cambridge, Mass.: HarvardUniversity Press. Manabe, S. and R.J. Stouffer. 1995. Simulation of abrupt climate change induced by freshwater input to the North Atlantic Ocean. Nature 378: 165-7. Martin, R.D. 198 1. Relative brain size and basal metabolic rate in terrestrial vertebrates. Nature 293: 57-60. McGrew, W. 1992. Chimpanzee material culture. Cambridge: Cambridge University Press. Milankovitch, M.M. 1941. Canon of Insolation and the Ice-age Problem. English Translation 1969. Israel Program for Scientific Translations. Washington: US Department of Commerce and National Science Foundation. Moore, B.R. 1996. The evolution of imitative learning. In: C.M. Heyes and B.G. Galef, Jr., eds., Social Learning in Animals: The Roots of Culture. San Diego: Academic Press. Pp. 245-65 Nottebohm, F. 1975. Continental patterns of song variability in Zonotrichia capensis: Some possible ecological correlates. Amer. Natur. 109: 605-624. Opdyke, N. 1995. Mammalian migration and climate over the past seven million years. In: E. S. Vrba, G. H. Denton, T. C. Partridge, L. H. Burckle, eds., Paleoclimate and Evolution With Emphasis on Human Origins. New Haven: Yale University Press. Pp. 109-14 Partridge, T.C., G.C. Bond, C.J.H. Hartnady, P.B. deMenocal and W.F. Ruddiman.1995. Climatic effects of late Neogene tectonism and vulcanism. In: E. S. Vrba, G. H. Denton, T. C. Partridge, L. H. Burckle, eds., Paleoclimate and Evolution With Emphasis on Human Origins. New Haven: Yale University Press. Pp. 8-23. Pilbeam, D. 1972. The Ascent of Man: An Introduction to Human Evolution. New York: Macmillan Pinker, S. 1994. The Language Instinct: How the Mind Creates Language. New York: William Morrow. Potts, R. 1996. Humanity's Descent: The Consequences of Ecological Instability. New York: William Morrow. Premak, D., and G. Woodruff. 1978. Does the chimpanzee have a theory of mind? Behav. Brain Sci. 1: 515-26. Raven, P.H. and D.I. Axelrod. 1978. Origins and Relationships of the California Flora. University of California Publications, Botany Vol. 72. Berkeley: University of California Press. Rogers, A.R. 1989. Does biology constrain culture? Amer. Anthropol. 90:819-3 1. Rogers, A.R. and H.C. Harpending. 1992. Population growth makes waves in the distribution of pairwise genetic differences. Molecular Biology and Evolution 9: 55269. Russon, A.E. and B.M.F. Galdikas. 1993. Imitation in free-ranging rehabilitant orangutans. J. Compar. Psychol. 107: 147-61. Schneider, S.H. and R. Londer. 1984. The Coevolution of Climate and Life. San Francisco: Sierra Club Books. Shackelton, N.J. and N.D. Opdyke. 1973. Oxygen isotope and paleomagnetic stratigraphy of equatorial Pacific core V28-238: oxygen isotope temperatures and ice volumes on a 10' and 106 year time scale. Quaternary Research 3: 39-55. Spelke, E.S. 1990. Principles of object perception. Cognitive Science 14: 29-56. Stanley, S.M. 1979. Macroevolution: Pattern and Process. San Francisco: W.H. Freeman. Stebbins, G. L., and J. Major. 1965. Endemism and speciation in the California flora. Ecol. Monger. 35: 1-35. Stringer, C., and C. Gamble. 1993. In Search of the Neanderthals: Solving the Puzzle of Human Origins. New York: Thames and Hudson. Terkel, J. 1996. Cultural transmission of feeding behavior in the black rat (Rattus rattus). In: C.M. Heyes and B.G. Galef, Jr., eds., Social Learning in Animals: The Roots of Culture. San Diego: Academic Press. Pp. 17-47. Tomasello, M.,. 1996. Do apes ape? In: C.M. Heyes and B.G. Galef, Jr., eds., Social Learning in Animals: The Roots of Culture. San Diego: Academic Press. Pp. 319-46. Tononi, G., 0. Sporns, and G.M. Edelman. 1994. A measure of brain complexity: Relating functional segregation and integration in the nervous system. Proc. Natl. Acad. Sci. USA 91: 5033-5037. Tooby, J and 1. DeVore. 1987. The reconstruction of hominid behavioral evolution through strategic modeling. In: W.G. Kinzey, ed., The Evolution of Human Behavior: Primate Models. Albany: State University of New York Press. Whiten, A., and R.W. Bryne, eds. 1988. Machiavellian Intelligence: Social Expertise and the Evolution of Intellect in Monkeys, Apes, and Humans. Oxford: Oxford University Press. Whiten, A., and D. Custance. 1996. Studies of imitation in chimpanzees and children. In: C.M. Heyes and B.G. Galef, Jr., eds., Social Learning in Animals: The Roots of Culture. San Diego: Academic Press. Pp. 291-318. Vrba, E. 1995. On the connection between paleoclimate and evolution. In: E. S. Vrba, G. H. Denton, T. C. Partridge, L. H. Burckle, eds., Paleoclimate and Evolution With Emphasis on Human Origins. New Haven: Yale University Press. Pp. 24-45. Wilson, E.O. 1975. Sociobiology: The New Synthesis. Cambridge Mass. Harvard University Press. Zentall, T. 1996. An analysis of imitative learning in animals. In: C.M. Heyes and B.G. Galef, Jr., eds., Social Learning in Animals: The Roots of Culture. San Diego: Academic Press. Pp. 221-243. Figures Figure 1. Simplification of the oxygen isotope record for the last 7 million years in marine sediments. The oxygen isotope record, based on samples of foraminiferan shells, is a proxy measure of volume of ice locked up in continental glaciers. Greater concentrations of the heavy 180 isotope indicate cold, high ice conditions. Thus, the climate over the last 7 million years has gotten cooler on average and very much more variable. A significant increase in variability occurred just after 6 million years ago, and again in the middle Pliocene. Another sharp deterioration occurred in the middle Pleistocene. (From Opdyke, 1995.) Figure 2. The oxygen isotope paleoclimate proxy from the Greenland Ice Core Project core drilled nearly 3,000 m to bedrock on a nearly stationary part of the Greenland ice cap. Ice depleted in the heavy isotope of oxygen (more negative values) indicates that large volumes of fresh water, depleted in 180, are stored in the continental glaciers. Note the very sharp peaks and troughs during the last cold period (Marine Isotope Stages 24). These are the Dansgaard-Oeschger and Heinrich fluctuations. The replicate GISP2 core agrees remarkably well with the GRIP core back to MIS 5c. The deeper part of the record, MIS 5e and 6, may be disturbed by ice flow (Grootes, et al., 1993). Note that the last I 0,000 years have been very much less variable that the other parts of the record. From GRIP (I 973). Information Available to Individual (x) Figure 3. The effect of the learning threshold (d) on the probability of acquiring the best behavior by individual learning or tradition. The curve shows the probability of obtaining a given estimate of the x of the average difference in yield between two environments from a small sample of years of experience of a young forager, assuming that the environment really is in a certain state, wet in this case. The task of the young forager is to decide what to do. If experience seems to show that the environment is indeed rather wet (x > d) the forager opts to forage alone. If experience seems to indicate that the environment is dry (x < -d), our forager incorrectly opts to forage cooperatively. If experience is ambiguous (-d < x < d), the young forager follows tradition (adopts mom's behavior). The width of the curve is a measure of the quality of information available from individual learning. In the curve illustrated, individual learning is fairly error prone, and selection is likely to favor setting wide values of d so as to avoid the chance of making an error based on noisy personal experience. However, if the environment is changing rapidly enough, it may be better for young foragers to depend on their own experience in spite of the risks because the risk that Mom is out of touch is also great. (From Boyd and Richerson, 1989.) Figure 4. The values of d, the learning threshold and L, the fi7action of young foragers acquiring their foraging mode by social teaming, as a function of reliability of personal experience (S) and the amount environmental variability fi7om generation to generation (m). The * indicate that these are the evolutionary equilibrium values of L and d, those that maximize fitness. Note that for environments that are harder to figure out (S large), the best thing to do is to rely more on social learning (d* and L* increase). Contrariwise, as the real change in the environment increases from generation to generation (m increases), it is best to trust more in own experience even at the risk happening to get the wrong answer by chance. (From Boyd and Richerson, 1989.) Figure 5. Contour plot of the differences in the fitness of populations using culture or social Teaming versus genetic transmission to convey information from the older to the younger generation. Both populations use the same individual teaming rule, the only difference is that the cultural population has the inheritance of acquired variation to that both Teaming and natural selection drive behavior in an adaptive direction. The cultural system is assumed also to have a higher random error rate. Here (a) measures the amount of social Teaming, R the environmental autocorrelation (the degree to which children's environments resemble those of their parents), and VH the amplitude of the environmental variation. The exact shape of the topography depends upon variables not pictured here, but the qualitative results hold for a wide range of those parameters. Cultural transmission is favored whenever the resemblance of parental to offspring environments is sufficiently high. It is especially favored when the environmental change is quite large but fairly slow on the generational time scale. In the situation where the advantage of the cultural system is maximal, the dependence on social Teaming is substantial, around a = 0.75. Note that if R gets large enough, genes win again because in a stable environment the higher "mutation" rate of social Teaming favors the more exact system of transmission. The late Tertiary onset of fluctuating but autocorrelated environments is, we suggest, highly conducive to a greater reliance on social Teaming. (From Boyd and Richerson, 1985: 127.) Figure 6. Cumulative frequency distributions of encephalization quotients in fossil and recent ungulates and carnivores. Encephalization coefficients measure brain size corrected for body weight. (From Jerison 1973: 311.) Humans survived ice age by sheltering in 'Garden of Eden', claim scientists By NIALL FIRTH UPDATED: 18:26 EST, 27 July 2010 • Comments (57) • Share ◦ ◦ ◦ The last humans on Earth may have survived an ice age by retreating to a small patch of land nicknamed 'the garden of Eden'. The strip of land on Africa's southern coast - around 240 miles east of Cape Town became the only place that remained habitable during the devastating ice age, scientists claim. The sudden change in temperature wiped out many species elsewhere around 195,000 years ago. Researchers believe this could account for the fact that humans have less genetic diversity than other species. Read more: http://www.dailymail.co.uk/sciencetech/article-1297765/Last-humans-Earthsurvived-Ice-Age-sheltering-Garden-Eden-claim-scientists.html#ixzz2OTt7saar Follow us: @MailOnline on Twitter | DailyMail on Facebook Some scientists even believe that the human race's population may have fallen to just a few hundred individuals who managed to survive in one location. Professor Curtis Marean, of the Institute of Human Origins at Arizona State University discovered ancient human artifacts in the isolated caves around an area known as Pinnacle Point, South Africa. 'Shortly after Homo sapiens first evolved, the harsh climate conditions nearly extinguished our species,' said Professor Marean. 'Recent finds suggest the small population that gave rise to all humans alive today survived by exploiting a unique combination of resources along the southern coast of Africa.' Humans would have been able to survive because of rich vegetation that was available in the area. The sea would have also been a good source of food as currents carrying nutrients would have passed by the shore, bringing with them a plentiful supply of fish, the team will say in a new research paper. Professor Marean said the caves contain archaeological remains going back at least 164,000 years. Professor Chris Stringer, a human origins expert at the Natural History Museum in London, said he agreed with Professor Marean's views on the early evolution of intelligence. But he said he was not convinced by the argument that one band of humans were the origin of modern man. 'However, I no longer think that there was ever a single small population of humans in one region of Africa from whom we are all uniquely descended. We know, for example, that there were early modern humans in Ethiopia 160,000 years ago and others in Morocco, and populations like those may also have contributed to our ancestry.' Many researchers believe that modern humans are thought to have evolved about 195,000 years ago in East Africa, and within 50,000 years had spread to other parts of the continent. It is thought that 70,000 years ago a dry period caused Red Sea levels to fall and the gap across its mouth to shrink from 18 miles to eight miles. A tribe of as few as 200 period took advantage of this and crossed to Arabia. Last year Professor Morean's team announced that they believed stone age blacksmiths mastered the use of fire to make tools at Pinnacle Point. Knowing how to use fire may have helped the early humans who left Africa 50,000 to 60,000 years ago to cope with colder conditions in Europe. It may also have given them a big advantage over the resident Neanderthals they encountered. By 35,000 years ago, the Neanderthals, a sub-species of humans whose own origins were in Africa, were mostly extinct. Professor Curtis Marean, , said: 'The command of fire, documented by our study of heat treatment, provides us with a potential explanation for the rapid migration of these Africans across glacial Eurasia. 'They were masters of fire and heat and stone, a crucial advantage as these tropical people penetrated the cold lands of the Neanderthal.' Read more: http://www.dailymail.co.uk/sciencetech/article-1297765/Last-humans-Earthsurvived-Ice-Age-sheltering-Garden-Eden-claim-scientists.html#ixzz2OTt5crB5 Follow us: @MailOnline on Twitter | DailyMail on Facebook The Human Paradox Several data points affirm that the human species has been around for about 200,000 years. Along the Omo River in Ethiopia, two partial human skulls were unearthed which dated to around 196,000 years ago. 1 Three partial human skulls were uncovered at Herto, Ethiopia, which dated to 160,000 years ago. This find was especially important because one of the skulls is nearly complete. Its distinctly human face has even been preserved. 2 Besides fossils, mitochondrial DNA indicates that we arose some 200,000 years ago in Africa. 3 Interestingly, we had the same brain size back then as we do today. Yet, for the vast majority of the time our species has been alive, we have been living like animals. Civilization came late in the game, and only accounts for about 2.75% of the time Homo sapiens has existed as a species. The first civilization on earth was ancient Sumer in modern day southern Iraq. Its emergence was sudden and unprecedented. Sumerian civilization seems to have been born overnight. Pottery, the wheel, division of labor, organized religion, government, warfare, manufacture of tools, written language, agriculture, city building, fortification – all the critical elements of human civilization were innovated within the fourth millennium BCE. This happened along the Euphrates River. Interestingly, Genesis specifically states that the Garden of Eden was by the Euphrates River. 4 Prior to Sumerian civilization, humans had been hunter-gatherers for 200,000 years. They traveled in small family clans. They had no written language. They had primitive tools. They had no kings, no governments, and no organized religion. Nowhere on the entire earth did any humans generate anything like civilization. These were humans like us. We are not talking about Neanderthals. Neanderthals were a separate species, and DNA analysis demonstrates that they were not our ancestors. 5 We are not talking about ape-people either. Ape-people, that is the genus Australopithecus, went extinct about 1.8 million years ago, long before we arrived. Neither are we discussing Homo erectus, whose name does not necessarily imply that primitive hominids practiced same-sex stimulation. We are strictly discussing humans, Homo sapiens, the same species we are today.For 200,000 years, humans like us lived like animals! They were just like us anatomically, and in terms of brain size.Then suddenly, for no apparent reason, we became civilized. Why, after 200,000 years of living like animals did we abruptly decide to start living like we do today? Why did civilization come about? Naturalistic evolutionists have an answer – the end of the most recent glacial period of the present ice ages. Earth has experienced ice ages for about three million years. Not all that time has been spent in cold temperatures. Rather, there have been about 20 cycles of cold snaps and warm spells. 6 The cold snaps are called "glacial" periods. The warm spells are called "interglacial" periods. About ten thousand years ago, the last glacial period came to an end. Earth’s weather became much more pleasant. We entered into one of the interglacial cycles. Fertile earth began to emerge in places that had once been covered with ice and snow. People came out of their caves and discovered how to plant crops. Previously, during the glacial period, the growing seasons had been much shorter, which retarded the development of agriculture. But now, the warm summer months lasted long enough to grow and harvest food. Agricultural production became more attractive than hunting and gathering. Instead of freezing in a tent, people could build a house and live there indefinitely. Instead of battling wild animals with spears, people could raise domesticated livestock. Instead of climbing trees to pick bananas, people could reach down to dig up a potato. The agricultural life was much better. The new way of obtaining food demanded a new society. People could no longer live in small tribes. They needed to build cities and forts to defend their food stores from thieves. They needed people to administer the cities. Kings were inaugurated for this purpose. They needed priests to appease the gods so that bad weather did not destroy their crops. Certain aspects of religion were invented for this purpose. They needed some way to till the ground and harvest their crops. New tools were invented for that purpose. The end of the last glacial period of the ice ages made agriculture possible. Agriculture made civilization possible. Civilization made technology possible, and that is why we stopped living like animals. That's the naturalists' explanation. But is this explanation adequate? One problem with the view presented above is that the emergence of civilization really doesn’t correspond with the end of the last glacial period as they claim. Civilization first began about 3,500 BCE. But the last glacial period of the ice age had already ended some 4,500 years before in 8,000 BCE. Since 8,000 BCE, the average temperature of the earth has not varied any more than two degrees Celsius. 7 Temperatures have been enormously stable. The warm climate we have today is the same climate humans enjoyed in 8,000 BCE. Variations from the norm have been comparatively minimal. Did it really take humankind a full 4,500 years to realize that agricultural civilization was the wave of the future? And if so, if they only progressed toward civilization gradually over 4,500 years, then why did all major elements of civilization emerge so suddenly in 3,500 BCE? Another problem is that there have been other interglacial periods during humanity’s existence. About 125,000 years ago, an interglacial period started that was similar to the one we are currently living in. It afforded early humans about 10,000 years of warm weather like ours before the earth was battered by another cold snap. 8 The humans who were living then had all the same opportunities that we did. They had the same mild climates we enjoy today. They basked in warm temperatures comparable to ours today. Most importantly, they were humans just like we are today, with the same anatomy, same brain size, and presumably the same intellectual capability. Why didn’t they create civilization? In just 5,500 years, civilization has brought us from jungle bunnies to computer geeks, from barbarians to cell phone junkies, from nomads to moon walkers. 125,000 years ago, humans had a 10,000 year window to accomplish everything we accomplished in just 5,500 years. Why didn’t they? If civilization was caused by the end of an ice age, then why didn’t the last interglacial 125,000 years ago cause the formation of human civilization back then? Moreover, there was a third interglacial about 185,000 to 215,000 years ago, about the same time our species first arrived on the scene. So humanity has been on the earth long enough to enjoy three long periods of very agreeable weather, and yet civilization only managed to take root the third time around. If civilization was made possible by an interglacial warm spell, then why didn’t civilization emerge the other two times we enjoyed a respite from the ice ages? If the end of the most recent glacial period did not clear the way for civilization, what did? Is it possible that there was a change in our intelligence without a change in our biology? Is it possible we suddenly got smarter without a change in our brain size? And what might cause such a sudden change in intelligence? Besides our intelligence, there is something else about humankind that makes us unique. Above all other species, humans feel a need for justice, empathy, and love for others. For 540 million years, complex life forms have been killing and eating each other with no sense of remorse, and without concern for their ecological impact. Humans are the first to question the morality of such a system. So there are two sparks in the human mind that separate us from other animals – intelligence and a moral conscience. Yet neither can be satisfactorily explained by the naturalists, for our intelligence came about long after our brain reached its current size, and our moral conscience defies the brutality necessary to succeed as a species under the rules of survival of the fittest. So what caused us to gain intelligence and a moral conscience? Could it be – God? Did God impart intelligence and love upon us poor apes? For 200,000 years, we lived as animals. Then, civilization suddenly began about the same time Adam and Eve are said to have been created. Perhaps the injection of Adam and Eve's super-human God-created DNA into the gene pool of the cave-people is what made the difference. However, the idea of an Almighty Creator raises a host of difficult questions. The God Paradox According to traditional theology, the Almighty Creator God sees everything, knows everything, created everything, and can do anything. This God is the God who created earthquakes and tornadoes, hurricanes and wildfires, sharks and mosquitoes, black widows and rattlesnakes, and lions and tigers and bears, who rip out the necks of other animals and eat them alive. And this God desires a better world, as the Prophet says: "The wolf shall dwell with the lamb, and the leopard shall rest with the young goat, and the calf and the lion and fatted livestock together, and a child shall lead them. The cow and the bear shall graze together… the lion shall eat straw like a cow, toddlers will play in snakes' dens… they shall not hurt nor destroy in all my holy mountain." 9 If God wanted non-violent animals, then why didn't he just create them non-violent to begin with? Theologians have a ready answer: God did create them non-violent, but the devil was thrown to earth, and therefore it is the devil who turned the animals to violence. God wants us to see the consequences of the devil's rebellion, so that we can learn a lesson from it. That way, when God reverts the world to non-violence in the future, we humans, the crown of his creation, can better appreciate his creation. This explanation is arguably plausible within the creationists' time frame of 6,000 years. However, it falls apart if the earth is much older. Animals have been ripping each other's necks out and eating each other for 540 million years. What was God's purpose in allowing this to continue for so long? Certainly it was not to teach us a lesson about the devil's rebellion, for we did not even exist back then. Was the lesson meant for trilobites and dimwitted reptiles? Moreover, the universe is even older – standing at 13.7 billion years. If the human species is the crown of creation, the only organism "made in the image of God," as Genesis 1 says, 10 then why did it take God 13.7 billion years to create us? What could delay God for 13.7 billion years? Was it really necessary to create failed experiments like dinosaurs and dodo birds before us? Was it really necessary to waste 3 billion years creating different types of bacteria and sea scum before creating the first true plant or animal? Once the Almighty Creator God is separated from the creationist time frame of a 6,000 year-old earth, it leaves that God in a theological no-mans' land, because there is little cover from the many arguments that can be launched against that God's very existence. Also, if we are in the image of God, then why do our bodies have the marks of evolution on them? We have a worthless organ called an appendix, which is good for nothing but exploding and killing us. We have a large amount of DNA that is repressed and doesn't even code for anything. It just takes up space and nutrients inside our cells. Why would the Almighty Creator God include irrelevant DNA in his code for life? Moreover, if we are the crown of God's creation, and if he created color and beauty for our enjoyment, then why can't we see colors in their full glory? We are more dependent upon our eyes than upon any other sensory organ, yet the color capacity of our eyes is inferior to that of goldfish and chickens! Far from being highly evolved, our color vision is no better than that of the most primitive of living reptiles, the crocodiles! We have only three cones for color, but many inglamorous species have more. These include flies, jumping spiders, and a certain shrimp-like praying mantis that lives at the bottom of the ocean. 11 If we are to assume a divine hand was involved in the making of our evolutionary history, it would have to be more along the lines of Intelligent Interference than Intelligent Design, for God did not perfectly create us, nor did God perfectly morph us from the lower apes. The human body was not designed by an all-knowing God. Rather, it came about haphazardly and imperfectly. 1 McDougall, Ian; Brown, Frank; Fleagle, John. Stratigraphic Placement and Age of Modern Humans from Kibish, Ethiopia. 2005, Nature 433, p 733-736 2 Stringer, Chris. Human Evolution: Out of Ethiopia. 2003, Nature 423, p 692-694 3 Cann, Rebecca L; Stoneking, Mark; Wilson, Allan C. Mitochondrial DNA and Human Evolution. 1987, Nature 325, p 31-36 4 Genesis 2:14 5 Jobling, Mark A; Hurles, Matthew E; Tyler-Smith, Chris. Human Evolutionary Genetics: Origins, Peoples & Disease. 2004, Garland Publishing, New York, NY, p 260-261 6 Gould, Stephen Jay; Andrews, Peter; Barber, John; Benton, Michael; Collins, Marianne; Janis, Christine; Kish, Ely; Morishima, Akio; Sepkoski, J John Jr; Stringer, Christopher; Tibbles, Jean-Paul; Cox, Steve. The Book of Life: An Illustrated History of the Evolution of Life on Earth. 2001, W W Norton & Co, New York, NY, p 208 7 Jouzel, J; Lorius, C; Petit, J R; Genthon, C; Barkov, N I; Kotlyakov, V M; Petrov, V M. Vostok Ice Core: A Continuous Isotope Temperature Record Over the Last Climatic Cycle (160,000 years). 1987, Nature 329, p 403-408 8 EPICA Community Members. Eight Glacial Cycles from an Antarctic Ice Core. Jun 10, 2004, Nature 429, p 623-628 9 Isaiah 11:6-9 10 Genesis 1:27 11 Kelber, Almut; Vorobyev, Misha; Osorio, Daniel. Animal Color Vision – Behavioral Tests and Physiological Concepts. 2003, Cambridge Philosophical Society, Biological Reviews, Vol 78, Issue 1, p 83-85 Graphic of Bison Skull, Utah Museum of Natural History Graphic of Crocodile, University of Wyoming Geological Museum Ice Age Migration? New DNA study suggests people moved between continents before recorded history Rob Waugh The Daily Mail Tue, 27 Mar 2012 08:22 CD © Unknown Early man: A third of people in modern Europe show genetic traces of populations from sub-Saharan Africa, leading researchers to conclude that people migrated between the continents as early as 11,000 years ago. People moved between Africa and Europe long before recorded history - and the migrations might have been driven by Europeans moving south to 'weather' ice ages. The genetic traces of long-forgotten migrations from Africa to Europe live on in Europeans today. A third of the genetic traces of sub-Saharan lineages in today's Europe come from prehistory. Researchers think that Europeans 'pushed south' by glaciers might have met with populations from sub-Saharan Africa. People moved between the continents as early as 11,000 years ago. Geneticists used mitochondrial DNA to look for the traces of ancient migrations. Mitochondrial DNA is passed directly from mother to child with no DNA from the father - and tiny changes in the sequence come to 'characterise' different populations, which can be used to trace movements and migrations of groups of humans in the past. Large numbers of people moved between Africa and Europe during recent and welldocumented time periods such as the Roman Empire, the Arab conquest, and the slave trade - but the researchers found that a third of sub-Saharan lineages came from before these movements. 'It was very surprising to find that more than 35 percent of the sub-Saharan lineages in Europe arrived during a period that ranged from more than 11,000 years ago to the Roman Empire times,' said Dr. Antonio Salas of the University of Santiago de Compostela and senior author of the study. 'The other 65% of European haplogroup L lineages arrived in more recent times.' The authors explain that these contacts likely connected sub-Saharan Africa to Europe not only via North Africa, but also directly by coastal routes. Salas said that it still remains unknown why there was genetic flow between the Africa and Europe in prehistoric times. One possible scenario is that some bidirectional flow was promoted when the last glaciation pushed some Europeans southward, until the glacier receded and populations returned north. In addition to tracing the genetic links of Africa and Europe back to prehistoric times, Salas expects that their work will also help those individuals who want to learn more about their own ancestry.