Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
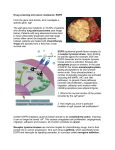
The Oncologist This material is protected by U.S. Copyright law. Unauthorized reproduction is prohibited. For reprints contact: [email protected] ® Small-Molecule Epidermal Growth Factor Receptor Tyrosine Kinase Inhibitors MAARTEN L. JANMAAT, GIUSEPPE GIACCONE Department of Medical Oncology, VU University Medical Center, Amsterdam, The Netherlands Key Words. EGFR · ZD1839 · Iressa · OSI-774 · Tarceva A BSTRACT The growth and proliferation of cells are usually tightly regulated processes that are activated by stimuli from their environment. Epidermal growth factor (EGF)-related peptides represent a class of molecules that can trigger cell proliferation, among several cellular processes, such as differentiation, migration, and survival. Binding of EGF-like peptides to the EGF receptor (EGFR) at the cell surface leads to a cascade of intracellular reactions that transduce signals to the nucleus, resulting in particular gene expression patterns. However, in many tumor cells, the regulation of EGFR activity is lost, due to increased or aberrant expression of the receptor or its ligands, and this contributes to many processes important for tumor growth, including cell proliferation, survival, angiogenesis, invasion, and metastasis. Many strategies have been developed that specifically target the EGFR and inhibit its activity. Of these, small-molecule tyrosine kinase inhibitors represent one of the most promising classes of anticancer agents. Here, we describe the status of small-molecule EGFR tyrosine kinase inhibitors in preclinical and clinical development. The Oncologist 2003;8:576-586 THE EPIDERMAL GROWTH FACTOR RECEPTOR PATHWAY The epidermal growth factor receptor (EGFR) (ErbB-1 or HER-1) is a member of the ErbB family of receptor tyrosine kinases (RTKs) that also includes the closely related ErbB-2 (HER-2/Neu), ErbB-3 (HER-3), and ErbB-4 (HER-4) receptors [1]. The EGFR is a 170-kDa protein located at the cell surface, consisting of an extracellular ligand-binding domain, a single transmembrane region, and an intracellular domain with tyrosine kinase activity. Activation of the receptor occurs when a ligand, such as transforming growth factor-α or EGF, binds to the ectodomain of the receptor, resulting in receptor dimerization with either another EGFR protein or another ErbB receptor; ErbB-2 is the preferred dimerization partner [2, 3]. The dimerization triggers activation of the intracellular kinase domain of the receptor, autophosphorylation of tyrosine residues in the intracellular domain, and subsequent recruitment and activation of downstream signaling molecules. Two important signaling routes that are activated by the EGFR involve the Ras-Raf-MEK-ERK and the PI3K-PDK1-Akt kinase pathways, which are implicated in cell proliferation, survival, and gene expression [1] (Fig. 1). Alternatively, the EGFR can be activated by stimuli that do not directly bind the receptor, such as hormones, lymphokines, and stress factors [4]. This transactivation of the EGFR appears to be essential for several mitogenic responses induced by the various stimuli, as suggested by the results of experiments using dominant negative EGFR mutants and specific kinase inhibitors [4]. Of note, Gilmore et al. recently demonstrated that insulinlike growth factor (IGF) transactivates the EGFR and that therapeutic inhibition of the EGFR by gefitinib (Iressa®, ZD1839; Astra Zeneca, Inc.; London, UK; http://www. astrazeneca.com) counteracts IGF-mediated inhibition of the proapoptotic protein BAD, resulting in cell death [5]. Activation of the EGFR can, thus, induce many different cellular responses. The actual physiological response Correspondence: Giuseppe Giaccone, M.D., Ph.D., Department of Medical Oncology, VU University Medical Center, De Boelelaan 1117, PO Box 7057, MB 1007 Amsterdam, The Netherlands. Telephone: 31-20-444-4340; Fax: 31-20-444-3844; e-mail: [email protected] Received March 14, 2003; accepted for publication August 18, 2003. ©AlphaMed Press 1083-7159/2003/$12.00/0 The Oncologist 2003;8:576-586 www.TheOncologist.com Small-Molecule EGFR-TKIs 577 Figure 1. The EGFR signaling pathway. Upon binding of a ligand, the EGFR dimerizes, resulting in the activation of the intracellular kinase (K) domain and subsequent autophosphorylation of tyrosine residues. This leads to the activation of intracellular kinase pathways such as the Ras-Raf-MEKERK and the PI3K-PDK1-AKT pathways, which, in turn, activate transcription factors in the nucleus. Several known biological responses to EGFR activation are shown. Two successful strategies to block the EGFR are depicted. EGFR-specific monoclonal antibodies, which compete with ligands for receptor binding thereby preventing EGFR kinase activation and EGFRTKIs that directly interact with the EGFR tyrosine kinase domain and inhibit its activity. Monoclonal antibodies (e.g. C225) Ligand Tyrosine kinase inhibitors (e.g. Iressa™) EGFR KK Ras Grb2 SOS SHC P13K Raf PDK1 Signal transduction MEK AKT Erk Gene transcription PP cyclin D1 myc Cyclin D1 DNA Jun Fos Myc depends on the cell context [1] as well as the particular ligand that is bound, which, in turn, regulates the dimer formation and subsequent signaling pathways that are activated [6, 7]. Adhesion Proliferation EGFR AS A TARGET FOR ANTICANCER THERAPEUTICS EGFR activity is essential for embryonic development, as null mutations in genetically modified mice are lethal [8-10]. More specifically, EGFR is involved in organogenesis of many mesoderm- and ectoderm-derived organs, such as the brain, heart, and lung [8-10]. In contrast to its critical role in embryogenesis, the EGFR lacks an essential physiological role in the adult organism, although ErbB receptors are involved in the development of the mammary gland [11]. However, it has become clear that the EGFR has a role in the development and progression of cancer, since increased or aberrant expression of the EGFR or its ligands are frequent in many types of tumors and correlate with a more aggressive disease and poorer prognosis [12-14]. In contrast, a discrepancy exists in the literature about the role of EGFR (over)expression as a prognostic factor in some tumor types, in a large part because of the wide variety in detection methods and cutoffs used to define overexpression [15]. Although EGFR expression is a strong prognostic factor in some tumors, such as head and neck, ovarian, breast, esophageal, bladder, and cervical cancers, it will be important to introduce a widely accepted standard test to evaluate EGFR expression levels in tissues. Amplification of the gene for the EGFR is most frequently detected in human Migration Survival Differentiation gliomas (40%) and is often associated with gene rearrangements [16]. In many cases, exon 2-7 is deleted, yielding a constitutively active receptor that is truncated and lacks most of the extracellular domain. This so-called EGFRvIII mutant has also been detected in medulloblastomas, breast and ovarian cancer, and non-small cell lung cancer (NSCLC) [17], though the impact of this mutant on tumor progression is not clear in these types of carcinomas. In addition to high expression levels and/or mutations, other potential mechanisms can induce aberrant EGFR kinase activity, such as ligand overexpression, heterodimerization with other ErbB members, in particular HER-2, and transactivation by heterologous signaling networks. Uncontrolled EGFR activity has been implicated in many aspects of tumor growth, including the promotion of cell proliferation, angiogenesis, invasion and metastasis, and survival [13, 14, 18]. Together, this provides a rationale for the inhibition of the EGFR as anticancer therapy, as suggested by Dr. Mendelsohn in the early 1980s [19-21]. Indeed, many agents have been developed that specifically target the receptor, ranging from toxin-conjugated anti-EGFR antibodies or ligands to antisense oligonucleotides [18, 22]. Among these, monoclonal antibodies directed against the ectodomain of the receptor and smallmolecule tyrosine kinase inhibitors (TKIs) are the most advanced in clinical development (Table 1), and the latter class of agents is discussed below. Janmaat, Giaccone 578 Table 1. EGFR-TKIs, their specificities, reversibilities, and development stages Class of compound Quinazolines Name Specificity Reversible Highest development stage gefitinib EGFR Yes Approved for third-line treatment of NSCLC in Japan and the U.S. OSI-774 EGFR Yes Phase III CI-1033 pan-ErbB No Phase II EKB-569 EGFR No Phase I PD-183805 pan-ErbB No Phase I Pyridopyrimidines PD-158780 pan-ErbB Yes Preclinical Pyrrolopyrimidines PKI-166 EGFR/ErbB-2 Yes Phase I, discontinued Other compounds GW-572016 EGFR/ErbB-2 Yes Phase II AG-1478 EGFR Yes Preclinical PRECLINICAL STUDIES WITH EGFR-TKIS The finding that mutations in the ATP-binding site of the intracellular domain of the EGFR disable ligand-induced responses [23-25] indicates that that part of the receptor is essential for EGFR tyrosine kinase activity and downstream signaling. In order to specifically inhibit EGFR kinase activity, hundreds of natural and synthetic compounds that compete with ATP for EGFR binding were screened. Many compounds of different chemical classes that effectively inhibit EGFR kinase activity have been identified, some of which are in late clinical development (Table 1). These molecules generally differ in their abilities to bind the EGFR ATP-binding pocket—either reversibly or irreversibly—or in their capacities to additionally inhibit other members of the ErbB family of receptors [26]. Many of these agents have been investigated in preclinical studies; of these, gefitinib and OSI-774 (erlotinib, Tarceva™; OSI Pharmaceuticals; Tarrytown, NY; http://www.osip.com) have been most extensively studied [27]. Gefitinib Gefitinib is an orally active, selective, and reversible EGFR-TKI that chemically belongs to the class of anilinoquinazolines [27]. Similar to the effects initially found with EGFR-blocking monoclonal antibodies [19-21], in vitro effects of gefitinib as a single agent were mainly cytostatic [28], although cytotoxic effects have been observed in a few cases [5, 29]. It has been suggested that gefitinib favors several proapoptotic mechanisms involving Bcl-2 family members, as the proapoptotic protein BAD is activated by gefitinib in breast cancer cells [5], while, conversely, overexpression of the apoptosis-suppressor Bcl-2 reverts gefitinib-induced cell death of vulval carcinoma cells [29]. Gefitinib is active against a wide variety of tumor cell lines [28, 29, 30-33], and the sensitivity of cells to gefitinib is likely to be affected by multiple factors, as the EGFR is part of a large signaling network [1]. Several cellular factors have been investigated that may be implicated in the sensitivity of cells to gefitinib. EGFR expression levels were initially found to be unrelated to response to gefitinib, since xenografts expressing high, moderate, and low amounts of EGFR all showed growth inhibition upon gefitinib treatment [28, 30], while later publications showed a correlation between the two in some cases [31]. In contrast to expression levels, EGFR activity status may be more important [15]. EGFR activity can be modulated by several mechanisms, for example, increased ligand expression or heterodimerization with ErbB2. Importantly, cells expressing high levels of ErbB-2 have been shown to be particularly sensitive to gefitinib [29, 32, 33], although no direct evidence has demonstrated an association between ErbB-2 levels and anti-EGFR response. Gefitinib-like agents may additionally inhibit ErbB-2 activity by the sequestering of ErbB-2 through the induction of signaling-inactive EGFR-ErbB-2 heterodimers [34, 35]. Furthermore, it has been established that proteins that are involved in cell cycle progression, in particular p27kip1 and cyclin D, play essential roles in the G1-phase cell cycle arrest observed in many cells treated with anti-EGFR agents [36, 37]. Finally, we and others have suggested that intrinsic and persistent activity of kinase pathways downstream of EGFR, such as the Ras-Raf-MEK-ERK and the PI3K-PDK1-Akt pathways, may provide a mechanism of resistance to gefitinib [5, 31, 32, 36]. Gefitinib has been combined with a variety of cytotoxic agents, resulting in enhanced antitumor effects in cultured cells and in vivo models, except in combination with gemcitabine [28, 30]. In addition, gefitinib treatment resulted in synergistic effects in combination with radiation [38-40]. Importantly, sequence-dependent effects were reported in cells treated with combinations of gefitinib with radiation or chemotherapy (cisplatin and/or 5-fluorouracil), with the best results observed when gefitinib was given before radiation and before or during chemotherapeutic treatments; whereas 579 some antagonistic effects were observed when gefitinib was given after cytotoxic treatments [38]. These data suggest the need for caution in the design of clinical trials using combinations of gefitinib and cytotoxic agents. A relatively new development is the preclinical evaluation of treatment with gefitinib combined with other novel, biological agents, such as the ErbB-2 monoclonal antibody trastuzumab [29, 32, 41]. ErbB-2 is the preferred dimerization partner for EGFR, and EGFR/ErbB-2 dimers are thought to induce more potent signals [2, 3]. Simultaneous inhibition of the EGFR and ErbB-2 resulted in additive or synergistic effects in ErbB-2-overexpressing breast cancer cells [29, 32, 41]. Those preclinical data initiated a currently ongoing phase II trial combining trastuzumab with gefitinib in breast cancer patients [42]. In addition to the often limited antiproliferative effects observed in vitro, several mechanisms of action that are only active in vivo, such as inhibition of angiogenesis and invasion and metastasis, have been attributed to gefitinib [40, 43-46]. The antiangiogenesis effect of gefitinib has been proposed to be the result of reduced secretion of angiogenesis factors [43, 44], while gefitinib can also directly inhibit the growth and cell-cell interactions of endothelial cells [40, 45]. Furthermore, the combination of gefitinib with a cytotoxic treatment, such as paclitaxel or radiation, potentiated the antiangiogenesis effect [40, 43]. OSI-774 OSI-774 is an EGFR-specific quinazoline derivative that inhibits the activity of purified EGFR in intact cells at nanomolar concentrations (50% inhibitory concentrations of 2 nM and 20 nM, respectively). An initial report showed that OSI-774 induced apoptosis and growth inhibition in several tumor cell lines in vitro, which was associated with the induction of p27kip1 expression and blockade in the G1 phase of the cell cycle [47]. The upregulation of p27kip1 and, possibly, restoration of Rb function are required for the antitumor action of EGFR inhibitors, including OSI-774 [37]. Moreover, OSI-774 has shown a substantial effect on the tumor growth of human HN5 xenografts growing in athymic mice [48] and on pancreas-derived xenografts, which was associated with a decrease in phosphorylation of ERK but not of Akt [49]. In skin and tumor biopsy specimens, phosphorylated forms of EGFR, Akt, and ERK were lower after treatment with OSI-774 [50, 51]. Synergistic effects were observed when OSI-774 was combined with cisplatin in preclinical models [48]. In addition, an enhancement of cytotoxic effects was reported when OSI-774 was combined with doxorubicin and gemcitabine, among other cytotoxic agents [52]. Moreover, OSI-774 selectively inhibited molecular effectors involved in the invasion of human glioblastoma cell lines expressing the EGFRvIII mutant [53]. This mutant Small-Molecule EGFR-TKIs receptor cannot be recognized by EGFR-specific monoclonal antibodies such as cetuximab, illustrating one of the advantages of EGFR-TKIs over monoclonal antibodies. OSI-774 is currently being tested in advanced clinical trials in various human malignancies (Table 1). Other Small-Molecule EGFR-TKIs Many other small-molecule inhibitors have been investigated in preclinical studies, some of which inhibit other ErbB kinases in addition to the EGFR. Several of these inhibitors are currently being tested in the clinic and are discussed here. PD-183805 and its water-soluble analog, CI-1033, are irreversible inhibitors of all ErbB receptors (Table 1). These agents have exhibited in vitro and in vivo antitumor activities as single agents and in combination with cytotoxic drugs in several tumor models [54-57]. Interestingly, Erlichman et al. showed synergistic effects of combinations of CI-1033 with the topoisomerase inhibitors SN-38 and topotecan in cells expressing the drug transporter breast cancer resistance protein (BCRP) [55]. A possible explanation for these results is that CI-1033 is a substrate of the BCRP pump and competes with SN-38 and topotecan for binding places, leading to an increased intracellular accumulation of the topoisomerase inhibitors. Recently, Schuetz et al. showed that gefitinib potently reversed BCRP-mediated resistance to topoisomerase inhibitors in vitro, resulting in synergistic antitumor effects in xenograft models, which were attributed to dramatic increases in the bioavailability of the topoisomerase inhibitors in mice [58]. Although the precise mechanism of BCRP inhibition by EGFR-TKIs is not known, it could be an important factor in the design of clinical combination studies with EGFR-TKIs. Synergistic apoptotic responses were found when cells were treated with combinations of CI-1033 and gemcitabine [57]. In that study, phosphorylation of ERK and Akt was inhibited, similar to observations with other EGFR-targeted agents, while concurrent activation of the p38 stress pathway was reported, together contributing to the apoptotic response. PKI-166 is a dual EGFR/ErbB-2 inhibitor that induces growth inhibition in several tumor-derived cell lines expressing high levels of EGFR and/or ErbB-2 [59] and exhibits antitumor activity against several human tumor models in mice, commonly associated with antiangiogenesis and antiinvasion effects [60-64]. Interestingly, when compared with an EGFR-specific kinase inhibitor (CGP-59326), PKI-166 was more efficient at inhibiting the in vitro growth of tumor cells in the presence of EGF-related ligands [59], suggesting greater antitumor effects for inhibitors that target both the EGFR and ErbB-2 compared with EGFR-specific agents. Similar results have been published with several other small-molecule EGFR-TKIs in preclinical studies. Of note, Janmaat, Giaccone GW-572016 is a novel dual EGFR/ErbB-2 kinase inhibitor that is currently being tested in the clinic and has been demonstrated to be active against in vitro and in vivo human tumor models [65, 66]. EKB-569 is an irreversible inhibitor of EGFR activity that potently inhibits the growth of cells overexpressing EGFR or ErbB-2 but has little effect on cells with low expression levels of these receptors [67, 68]. In summary, the mechanism of action of the small-molecule EGFR-TKIs involves the direct inhibition of EGFR activity and/or the activities of other ErbB members. Inhibition of downstream Ras/ERK and PI3K/Akt kinase pathways has been linked, in many studies, with the antiproliferative and, sometimes, proapoptotic effects of the EGFR-TKIs. In addition, p27kip1 upregulation appears to be essential for the growth delay induced by these agents, while expression levels of EGFR seem to be of minor importance. In animal models, EGFR-TKIs have been shown to be potent inhibitors of tumor growth, antiangiogenesis agents, and inhibitors of invasion and metastasis of tumor cells when given as a single drug or in combination with a cytotoxic treatment. CLINICAL STUDIES WITH EGFR-TKIS Gefitinib Gefitinib is the EGFR-TKI that is the furthest advanced in clinical development, and registration was obtained in Japan in July 2002 and in the U.S. in May 2003 for third-line therapy of advanced NSCLC. The results of three phase I studies in patients with advanced solid tumors have been published [69-71]. This was one of the first targeted agents that was clinically tested, and the goals were clearly different from trials with cytotoxic agents. Instead of the maximum tolerated dose, one goal was to determine the optimal biological dose. Moreover, the common expression of the EGFR in solid tumors and preclinical evidence suggested a broad antitumor activity, leading to the inclusion of multiple tumor types and no selection of patients based on EGFR expression in their tumors. Other goals were to establish pharmacokinetic and pharmacodynamic relationships. Across the dose range tested (50-1,000 mg/day), the most frequent adverse events were dose-dependent acneiform skin rash and grade 1 or 2 diarrhea. The latter was dose limiting, being severe and frequent in patients given doses over 600 mg/day. Other toxicities that were observed include nausea and transient and asymptotic transaminitis. All of these side effects were manageable and reversible on cessation of treatment. To evaluate the effect of gefitinib on EGFR-TK activity, biopsies of the skin, which are known to express the EGFR, were analyzed for known, EGFR-dependent molecular markers and downstream effects on proliferation before and after 28 days of treatment. These studies showed inhibition 580 of EGFR-regulated signaling in patients treated with gefitinib consistent with the proposed mechanism of action, with downregulation of activated EGFR, mitogen-activated protein kinase, and the proliferation marker Ki67 and upregulation of p27kip1, phosphorylated signal transducer and activator of transcription 3, and apoptotic cells [70, 72]. In those phase I studies, promising antitumor activities were observed in patients with NSCLC, and head and neck, ovarian, colorectal, prostate, and breast cancers [69-71, 73]. The promising phase I results prompted the rapid initiation of phase II studies in pretreated patients with NSCLC, and prostate, breast, colorectal, and head and neck cancers. Currently, data are available from two large randomized phase II studies in pretreated NSCLC patients randomized to receive either a 250-mg or a 500-mg daily dose of gefitinib, doses that achieved sufficient blood concentrations to inhibit EGFR activation in preclinical models. One study, the Iressa Dose Evaluation in Advanced Lung Cancer 1 (IDEAL1) trial, included 210 patients who had received one prior chemotherapy regimen, which included a platinum drug [74]. The response rate was 18.5%, while approximately 40% of patients experienced symptom improvements. The other study (IDEAL2) included 216 patients who had received two or more chemotherapy regimens, which included a platinum drug and docetaxel. In that study, the response rate was 10% [75]. In both studies, withdrawals and grade III-IV adverse events were more frequent in patients receiving the higher dose, while the response rates were comparable, indicating that activity achieved at doses of 250 mg and higher doses did not generate better efficacy but only greater toxicities [76]. As a result of those studies, gefitinib was approved for the third-line treatment of NSCLC in Japan and, recently, in the U.S. as well; whereas approval in the rest of the world is being assessed at this time. Other phase II studies with gefitinib as monotherapy or in combination with other therapies are under way for hormone-refractory prostate cancer, advanced breast cancer, advanced colorectal cancer, head and neck cancer, esophageal cancer, ovarian cancer, and glioblastomas, among others. Preliminary data indicate that gefitinib is generally well tolerated in combination with several cytotoxic therapies, and promising results have been observed, including those from a phase II monotherapy trial in recurrent or metastatic head and neck cancer [77] and in patients with advanced breast cancer [78]. In the latter report, low levels of phosphorylated Akt (pAkt) were indicative of tumor response, suggesting low pAkt as a prognostic factor for response. At the same time, two very large randomized studies with gefitinib and combination chemotherapy were concluded, and preliminary results were presented recently [79, 80]. In those studies, chemotherapy-naïve patients with advanced NSCLC 581 were randomized to receive chemotherapy with either placebo or two different doses of gefitinib (250 or 500 mg). In the Iressa NSCLC Trial Assessing Combination Treatment 2 (INTACT2) trial, chemotherapy consisted of carboplatinpaclitaxel, which is standard in North America, and in the INTACT1 trial, chemotherapy consisted of cisplatin-gemcitabine, which is more frequently employed in Europe. Unfortunately, both studies failed to reach the major end point, which was improvement in survival by the addition of gefitinib, or any of the other end points, which included progression-free survival time, time to worsening of symptoms, objective tumor response, and quality of life. As the studies were robust and well designed, it should be concluded that these efficacy results are definitive. However, the studies did confirm the safety and tolerability of gefitinib, with comparable toxicity profiles in the control and the gefitinib treatment arms, with the exceptions of an expected dose-dependent skin toxicity and diarrhea in patients treated with gefitinib and two- to threefold more drug-related withdrawals in the 500 mg/day gefitinib arm in both studies. Although the results are disappointing, the high expectations may have been too optimistic for several reasons. One of these reasons may be the lack of patient selection, which may have diluted a possible beneficial effect of the addition of gefinitib to combination chemotherapy. The impact of EGFR expression levels on sensitivity is still an issue, even though preclinical data point to the lack of an effect. Other markers that may be used for selection are activated forms of EGFR and downstream effectors. Recently, the expression of EGFR (HER-2/Neu) was tested in 43 patients with advanced NSCLC who received 250 mg/day of gefinitib. No correlation with response or survival was observed [81]. Similarly, there was no correlation between response and the EGFR immunohistochemical results in patients who were entered in the IDEAL phase II studies [82]. It is theoretically possible that tumors pretreated with chemotherapy are more sensitive to EGFR-TKIs than chemotherapy-naïve tumors, which would be consistent with the positive results seen in the IDEAL1 and IDEAL2 trials. This hypothesis, together with the results of the INTACT studies, suggests that sequential administration of chemotherapy followed by gefinitib may be a more effective approach. Although no phase II studies have so far been performed for the first-line therapy of advanced NSCLC with gefinitib alone, data reported from the extended access program seem to indicate definite activity in that setting. Provocative results have been reported in the treatment of bronchioalveolar carcinoma [83]. This tumor type has a definitively different biology than the common adenocarcinomas of the lung, as it is more frequently observed in women and nonsmokers. Female gender and no smoking history (and Japanese race in the IDEAL1 study) were, in fact, identified in phase II studies as Small-Molecule EGFR-TKIs prognostic factors for response in advanced NSCLC [74]. However, it is evident that further investigation is needed to improve combination treatment with gefitinib in NSCLC patients and to identify the subset of patients that benefit from gefitinib treatment. Moreover, this drug is being investigated in studies of a large number of other malignancies, either as a single agent or as part of a combination therapy. OSI-774 OSI-774 is structurally related to gefitinib and has very similar toxicity and safety profiles, with skin rash and diarrhea as dose-limiting toxicities. Promising activity has been seen in phase I trials, with some reports of complete responses [84, 85]. The dose chosen for further evaluation after phase I studies is 150 mg/day, which is just below the maximum tolerated dose of 200 mg/day. To date, the results of three monotherapy phase II trials have been presented. In a study with 56 NSCLC patients, seven patients (11%) achieved partial responses, while 19 patients (34%) had stable disease [85]. The response rate to OSI-774 in NSCLC is similar to that obtained with gefitinib and C225, indicating a consistent pattern of activity of this class of agents in NSCLC. Promising phase II results have also been reported for advanced ovarian cancer and refractory head and neck cancer, with response rates of 8.8% and 13%, respectively, and disease stabilization in 44% and 29% of patients, respectively [86, 87]. Many phase I/II studies in various malignant diseases are being carried out at this time with OSI-774 in combination with several chemotherapeutics and/or radiation [88]. In those studies, responses have been reported for NSCLC, penile carcinoma, head and neck cancer, and mesothelioma, while prolonged disease stabilization was observed in NSCLC, mesothelioma, and head and neck, bladder, ovary, stomach, and skin cancers [89-91]. Two large international studies of OSI-774 have been concluded in the first-line treatment of advanced NSCLC in combination with chemotherapy, versus chemotherapy alone, with the same chemotherapy regimens used in the gefitinib phase III studies (carboplatin-paclitaxel and cisplatin-gemcitabine). The results of those studies are awaited with interest and some trepidation in the light of the negative results with gefitinib. As with the antibody C225 [92], there has been a retrospective analysis of correlation between survival and intensity of skin rash. Across several studies and different tumor types, there was a consistent correlation between skin toxicity and survival [93]. Other Small Molecules CI-1033 is an irreversible inhibitor of all four members of the ErbB family of receptors. The data from several phase Janmaat, Giaccone I studies, in which several different administration schedules were tested, have been presented. All schedules tested were generally well tolerated. Side effects included the ones observed with the other small molecules, with diarrhea and skin rash as dose-limiting toxicities, as well as reports of thrombocytopenia and allergy [94]. Unfortunately, no complete or partial responses were reported from phase I trials, though several patients achieved stable disease [95]. Phase II studies are under way. A large, randomized phase II trial in patients with advanced NSCLC, who failed prior platinum-based chemotherapy, is presently testing CI-1033 in three different schedules and doses in patients selected on the basis of having at least one of the four HER receptors positive. Phase I studies with the dual EGFR/ErbB-2 inhibitor PKI-166 have recently concluded. The major toxicities were similar to those of the other two small molecules, but an apparently higher incidence of liver toxicity was noted with this agent. Therefore, further development of PKI-166 was discontinued. The first phase I/II data with the dual EGFR/ErbB-2 kinase inhibitor GW-572016 reveal that the compound is well tolerated in patients at concentrations up to 1,250 mg/day as monotherapy as well as in combination with chemotherapeutics [96-102]. In addition to skin rash and diarrhea, headache was one of the most common adverse events. In one study, the inhibition of activated Akt, activated ERK, and cyclin D protein was associated with tumor cell apoptosis and regression of metastasis, and was predictive of clinical response [102]. Interestingly, two trastuzumab-resistant breast cancer patients had objective responses [102], while two gefitinib-resistant NSCLC patients achieved minor responses [98], suggesting that the dual specificity of GW572016 may be more effective in some patients than more specific agents. Phase II trials are under way, including ones in patients with trastuzumab-refractory metastatic breast cancer and metastatic colorectal cancer [99, 100]. EKB-569 is an irreversible EGFR-specific TKI that inhibits the growth of tumor cell lines that overexpress EGFR or ErbB-2 in vitro and in vivo [67]. The data from three preliminary phase I trials have been presented. In those trials, several treatment schedules were tested as monotherapy in patients with advanced-stage solid tumors [103] and in combination with cytotoxic agents in patients with advanced pancreatic [104] and colorectal cancers [105]. The results again show mild diarrhea and skin rash as the major toxicities, indicating that EKB-569 was generally well tolerated. CONCLUSIONS The emerging clinical data in EGFR-targeted therapy point to a role for these types of agents in the treatment of 582 cancer. However, the disappointing results of the INTACT trials with gefitinib demonstrate that the use of these agents in combination with chemotherapy is not straightforward and that additional investigation of optimal treatment schedules and sequences is warranted. The results of phase III trials with OSI-774 in combination with chemotherapy for NSCLC patients, which are very similar to the studies with gefitinib, are eagerly awaited, because they might reveal differences between the structurally related compounds. Alternative strategies may include the combination of anti-EGFR agents with radiation or other novel, targeted agents, such as Herceptin®, which have been demonstrated to be effective combinations in preclinical studies [29, 32, 38-41] and are currently being tested in patients. In contrast to combination strategies, the superior response rates for pretreated NSCLC patients with gefitinib and OSI-774 as single agents, along with a high rate of disease control, symptom relief, and good tolerability, support the use of anti-EGFR agents as monotherapy, and prospective studies in chemotherapy-naïve patients need to be performed. In addition to the treatment of NSCLC, various trials have concluded or are under way to test the efficacy of EGFR-targeted therapy against other tumor types, such as breast, prostate, head and neck, and colon cancers, as single-agent therapy and in combination with chemotherapy. Preliminary data show good tolerability of EGFR-TKIs in combination treatments, and promising results with several tumor types have been observed. Another issue to be addressed is whether the broader selectivity of some EGFR-TKIs impacts their antitumor efficacies. Objective and minor responses were achieved with the EGFR/ErbB-2 inhibitor GW-572016 in patients pretreated with gefitinib or trastuzumab, indicating that a broader selectivity may be more effective in some cases. However, a major drawback to this is the higher toxicity observed in patients treated with agents targeting multiple ErbB receptors. A good alternative could be treatment with combinations of targeted agents, such as combined treatment with gefitinib and trastuzumab, which is being assessed at this time in patients with advanced breast cancer [42]. The major challenge will be to identify the subset of patients that will benefit from anti-EGFR therapy. Data from clinical trials with anti-EGFR agents show that responders had a higher incidence of skin rash than nonresponders [92, 93]. Although more evidence should be gathered, this suggests skin rash as a potential surrogate marker for clinical activity. In conclusion, EGFR-TKIs have potential antitumor activity against many types of cancer, including NSCLC. However, more work is needed to improve their efficacy as single-agent therapy (i.e., via patient selection) or in combination with chemotherapy, radiotherapy, or other novel (targeted) agents. 583 Small-Molecule EGFR-TKIs R EFERENCES 1 Yarden Y, Sliwkowski MX. Untangling the ErbB signalling network. Nat Rev Mol Cell Biol 2001;2:127-137. receptor in multiple human tumors. Cancer Res 1995;55:55365539. 2 Karunagaran D, Tzahar E, Beerli RR et al. ErbB-2 is a common auxiliary subunit of NDF and EGF receptors: implications for breast cancer. EMBO J 1996;15:254-264. 18 Woodburn JR. The epidermal growth factor receptor and its inhibition in cancer therapy. Pharmacol Ther 1999;82:241-250. 3 Graus-Porta D, Beerli RR, Daly JM et al. ErbB-2, the preferred heterodimerization partner of all ErbB receptors, is a mediator of lateral signaling. EMBO J 1997;16:1647-1655. 4 Carpenter G. Employment of the epidermal growth factor receptor in growth factor-independent signaling pathways. J Cell Biol 1999;146:697-702. 5 Gilmore AP, Valentijn AJ, Wang P et al. Activation of BAD by therapeutic inhibition of epidermal growth factor receptor and transactivation by insulin-like growth factor receptor. J Biol Chem 2002;277:27643-27650. 6 Pinkas-Kramarski R, Soussan L, Waterman H et al. Diversification of Neu differentiation factor and epidermal growth factor signaling by combinatorial receptor interactions. EMBO J 1996;15:2452-2467. 19 Mendelsohn J, Baselga J. The EGF receptor family as targets for cancer therapy. Oncogene 2000;19:6550-6565. 20 Mendelsohn J. Epidermal growth factor receptor inhibition by a monoclonal antibody as anticancer therapy. Clin Cancer Res 1997;3:2703-2707. 21 Mendelsohn J. The epidermal growth factor receptor as a target for cancer therapy. Endocr Relat Cancer 2001;8:3-9. 22 Ciardiello F, Tortora G. A novel approach in the treatment of cancer: targeting the epidermal growth factor receptor. Clin Cancer Res 2001;7:2958-2970. 23 Honegger AM, Szapary D, Schmidt A et al. A mutant epidermal growth factor receptor with defective protein tyrosine kinase is unable to stimulate proto-oncogene expression and DNA synthesis. Mol Cell Biol 1987;7:4568-4571. 7 Riese DJ 2nd, van Raaij TM, Plowman GD et al. The cellular response to neuregulins is governed by complex interactions of the erbB receptor family. Mol Cell Biol 1995;15:5770-5776. 24 Honegger AM, Dull TJ, Felder S et al. Point mutation at the ATP binding site of EGF receptor abolishes protein-tyrosine kinase activity and alters cellular routing. Cell 1987;51:199209. 8 Miettinen PJ, Berger JE, Meneses J et al. Epithelial immaturity and multiorgan failure in mice lacking epidermal growth factor receptor. Nature 1995;376:337-341. 25 Chen WS, Lazar CS, Poenie M et al. Requirement for intrinsic protein tyrosine kinase in the immediate and late actions of the EGF receptor. Nature 1987;328:820-823. 9 Threadgill DW, Dlugosz AA, Hansen LA et al. Targeted disruption of mouse EGF receptor: effect of genetic background on mutant phenotype. Science 1995;269:230-234. 26 Noonberg SB, Benz CC. Tyrosine kinase inhibitors targeted to the epidermal growth factor receptor subfamily: role as anticancer agents. Drugs 2000;59:753-767. 10 Sibilia M, Wagner EF. Strain-dependent epithelial defects in mice lacking the EGF receptor. Science 1995;269:234-238. 27 Arteaga CL, Johnson DH. Tyrosine kinase inhibitorsZD1839 (Iressa). Curr Opin Oncol 2001;13:491-498. 11 Troyer KL, Lee DC. Regulation of mouse mammary gland development and tumorigenesis by the ERBB signaling network. J Mammary Gland Biol Neoplasia 2001;6:7-21. 28 Ciardiello F, Caputo R, Bianco R et al. Antitumor effect and potentiation of cytotoxic drugs activity in human cancer cells by ZD-1839 (Iressa), an epidermal growth factor receptor-selective tyrosine kinase inhibitor. Clin Cancer Res 2000;6:2053-2063. 12 Salomon DS, Brandt R, Ciardiello F et al. Epidermal growth factor-related peptides and their receptors in human malignancies. Crit Rev Oncol Hematol 1995;19:183-232. 13 Ozanne B, Richards CS, Hendler F et al. Over-expression of the EGF receptor is a hallmark of squamous cell carcinomas. J Pathol 1986;149:9-14. 14 Tang CK, Gong XQ, Moscatello DK et al. Epidermal growth factor receptor vIII enhances tumorigenicity in human breast cancer. Cancer Res 2000;60:3081-3087. 15 Arteaga CL. Epidermal growth factor receptor dependence in human tumors: more than just expression? The Oncologist 2002;7(suppl 4):31-39. 29 Janmaat ML, Kruyt FA, Rodriguez JA et al. Response to epidermal growth factor receptor inhibitors in non-small cell lung cancer cells: limited antiproliferative effects and absence of apoptosis associated with persistent activity of extracellular signal-regulated kinase or Akt kinase pathways. Clin Cancer Res 2003;9:2316-2326. 30 Sirotnak FM, Zakowski MF, Miller VA et al. Efficacy of cytotoxic agents against human tumor xenografts is markedly enhanced by coadministration of ZD1839 (Iressa), an inhibitor of EGFR tyrosine kinase. Clin Cancer Res 2000;6:4885-4892. 16 Sauter G, Maeda T, Waldman FM et al. Patterns of epidermal growth factor receptor amplification in malignant gliomas. Am J Pathol 1996;148:1047-1053. 31 Magne N, Fischel JL, Dubreuil A et al. Influence of epidermal growth factor receptor (EGFR), p53 and intrinsic MAP kinase pathway status of tumour cells on the antiproliferative effect of ZD1839 (“Iressa”). Br J Cancer 2002;86:1518-1523. 17 Moscatello DK, Holgado-Madruga M, Godwin AK et al. Frequent expression of a mutant epidermal growth factor 32 Moasser MM, Basso A, Averbuch SD et al. The tyrosine kinase inhibitor ZD1839 (“Iressa”) inhibits HER2-driven Janmaat, Giaccone signaling and suppresses the growth of HER2-overexpressing tumor cells. Cancer Res 2001;61:7184-7188. 33 Anderson NG, Ahmad T, Chan K et al. ZD1839 (Iressa), a novel epidermal growth factor receptor (EGFR) tyrosine kinase inhibitor, potently inhibits the growth of EGFR-positive cancer cell lines with or without erbB2 overexpression. Int J Cancer 2001;94:774-782. 34 Arteaga CL, Ramsey TT, Shawver LK et al. Unliganded epidermal growth factor receptor dimerization induced by direct interaction of quinazolines with the ATP binding site. J Biol Chem 1997;272:23247-23254. 35 Lichtner RB, Menrad A, Sommer A et al. Signaling-inactive epidermal growth factor receptor/ligand complexes in intact carcinoma cells by quinazoline tyrosine kinase inhibitors. Cancer Res 2001;61:5790-5795. 36 Moulder SL, Yakes FM, Muthuswamy SK et al. Epidermal growth factor receptor (HER1) tyrosine kinase inhibitor ZD1839 (Iressa) inhibits HER2/neu (erbB2)-overexpressing breast cancer cells in vitro and in vivo. Cancer Res 2001;61:8887-8895. 584 46 Mandal M, Adam L, Wang RB et al. Inhibition of p21-activated kinase 1, directional cell motility and invasion of growth-factor-activated human cancer cells by the selective epidermal growth factor receptor tyrosine kinase inhibitor (EGFR-TKI) ZD1839 (´Iressa´). Proc Am Assoc Cancer Res 2002;43:786a. 47 Moyer JD, Barbacci EG, Iwata KK et al. Induction of apoptosis and cell cycle arrest by CP-358,774, an inhibitor of epidermal growth factor receptor tyrosine kinase. Cancer Res 1997;57:4838-4848. 48 Pollack VA, Savage DM, Baker DA et al. Inhibition of epidermal growth factor receptor-associated tyrosine phosphorylation in human carcinomas with CP-358,774: dynamics of receptor inhibition in situ and antitumor effects in athymic mice. J Pharmacol Exp Ther 1999;291:739-748. 49 Ng SS, Tsao MS, Nicklee T et al. Effects of the epidermal growth factor receptor inhibitor OSI-774, Tarceva, on downstream signaling pathways and apoptosis in human pancreatic adenocarcinoma. Mol Cancer Ther 2002;1:777-783. 37 Busse D, Doughty RS, Ramsey TT et al. Reversible G(1) arrest induced by inhibition of the epidermal growth factor receptor tyrosine kinase requires up-regulation of p27(KIP1) independent of MAPK activity. J Biol Chem 2000;275:6987-6995. 50 Rowinsky EK, Hammond L, Siu L et al. Dose-schedule-finding pharmacokinetic (PK). Biologic, and functional imaging studies of OSI-774, a selective epidermal growth factor receptor (EGFR) tyrosine kinase (TK) inhibitor. Proc Am Soc Clin Oncol 2001;20:5a. 38 Magne N, Fischel JL, Dubreuil A et al. Sequence-dependent effects of ZD1839 (‘Iressa’) in combination with cytotoxic treatment in human head and neck cancer. Br J Cancer 2002;86:819-827. 51 Hidalgo M, Malik S, Rowinsky EK. Inhibition of the epidermal growth factor receptor (EGFR) by OSI-774, a specific EGFR inhibitor in malignant and normal tissues of cancer patients. Proc Am Soc Clin Oncol 2001;20:281a. 39 Williams KJ, Telfer BA, Stratford IJ et al. ZD1839 (‘Iressa’), a specific oral epidermal growth factor receptor-tyrosine kinase inhibitor, potentiates radiotherapy in a human colorectal cancer xenograft model. Br J Cancer 2002;86:1157-1161. 52 Elsayed YA, Sausville EA. Selected novel anticancer treatments targeting cell signaling proteins. The Oncologist 2001;6:517-537. 40 Huang SM, Li J, Armstrong EA et al. Modulation of radiation response and tumor-induced angiogenesis after epidermal growth factor receptor inhibition by ZD1839 (Iressa). Cancer Res 2002;62:4300-4306. 41 Normanno N, Campiglio M, De LA et al. Cooperative inhibitory effect of ZD1839 (Iressa) in combination with trastuzumab (Herceptin) on human breast cancer cell growth. Ann Oncol 2002;13:65-72. 53 Lal A, Glazer CA, Martinson HM et al. Mutant epidermal growth factor receptor up-regulates molecular effectors of tumor invasion. Cancer Res 2002;62:3335-3339. 54 Gieseg MA, de Bock C, Ferguson LR et al. Evidence for epidermal growth factor receptor-enhanced chemosensitivity in combinations of cisplatin and the new irreversible tyrosine kinase inhibitor CI-1033. Anticancer Drugs 2001;12:683-690. 42 Moulder SL, Arteaga CL. A phase I/II trial of trastuzumab and gefitinib in patients with metastatic breast cancer that overexpresses HER2/neu (ErbB-2). Clin Breast Cancer 2003;4:142-145. 55 Erlichman C, Boerner SA, Hallgren CG et al. The HER tyrosine kinase inhibitor CI1033 enhances cytotoxicity of 7ethyl-10-hydroxycamptothecin and topotecan by inhibiting breast cancer resistance protein-mediated drug efflux. Cancer Res 2001;61:739-748. 43 Ciardiello F, Caputo R, Bianco R et al. Inhibition of growth factor production and angiogenesis in human cancer cells by ZD1839 (Iressa), a selective epidermal growth factor receptor tyrosine kinase inhibitor. Clin Cancer Res 2001;7:1459-1465. 56 Rao GS, Murray S, Ethier SP. Radiosensitization of human breast cancer cells by a novel ErbB family receptor tyrosine kinase inhibitor. Int J Radiat Oncol Biol Phys 2000;48:15191528. 44 Hirata A, Ogawa S, Kometani T et al. ZD1839 (Iressa) induces antiangiogenic effects through inhibition of epidermal growth factor receptor tyrosine kinase. Cancer Res 2002;62:2554-2560. 57 Nelson JM, Fry DW. Akt, MAPK (Erk1/2), and p38 act in concert to promote apoptosis in response to ErbB receptor family inhibition. J Biol Chem 2001;276:14842-14847. 45 Wakeling AE, Guy SP, Woodburn JR et al. ZD1839 (Iressa): an orally active inhibitor of epidermal growth factor signaling with potential for cancer therapy. Cancer Res 2002;62:57495754. 58 Schuetz JD, Leggus M, Sampath J et al. Potent BCRP inhibition by the Erbβ1 inhibitor ZD1839 (‘IRESSA’) dramatically enhances oral bioavailability of topotecan and irinotecan (CPT-11) in mice. Proc Am Assoc Cancer Res 2002;43:1351a. 585 59 Motoyama AB, Hynes NE, Lane HA. The efficacy of ErbB receptor-targeted anticancer therapeutics is influenced by the availability of epidermal growth factor-related peptides. Cancer Res 2002;62:3151-3158. 60 Mellinghoff IK, Tran C, Sawyers CL. Growth inhibitory effects of the dual ErbB1/ErbB2 tyrosine kinase inhibitor PKI-166 on human prostate cancer xenografts. Cancer Res 2002;62:5254-5259. 61 Baker CH, Solorzano CC, Fidler IJ. Blockade of vascular endothelial growth factor receptor and epidermal growth factor receptor signaling for therapy of metastatic human pancreatic cancer. Cancer Res 2002;62:1996-2003. 62 Brandt R, Wong AM, Hynes NE. Mammary glands reconstituted with Neu/ErbB2 transformed HC11 cells provide a novel orthotopic tumor model for testing anti-cancer agents. Oncogene 2001;20:5459-5465. 63 Solorzano CC, Baker CH, Tsan R et al. Optimization for the blockade of epidermal growth factor receptor signaling for therapy of human pancreatic carcinoma. Clin Cancer Res 2001;7:2563-2572. 64 Bruns CJ, Solorzano CC, Harbison MT et al. Blockade of the epidermal growth factor receptor signaling by a novel tyrosine kinase inhibitor leads to apoptosis of endothelial cells and therapy of human pancreatic carcinoma. Cancer Res 2000;60:2926-2935. 65 Xia W, Mullin RJ, Keith BR et al. Anti-tumor activity of GW572016: a dual tyrosine kinase inhibitor blocks EGF activation of EGFR/erbB2 and downstream Erk1/2 and AKT pathways. Oncogene 2002;21:6255-6263. 66 Mullin RJ, Keith BR, Murray PI et al. Combination therapy with the dual EGFR-ERBB-2 tyrosine kinase inhibitor GW572016. Proc Am Soc Clin Oncol 2003;22:970a. 67 Wissner A, Brawner Floyd MB, Rabindran SK et al. Syntheses and EGFR and HER-2 kinase inhibitory activities of 4-anilinoquinoline-3-carbonitriles: analogues of three important 4-anilinoquinazolines currently undergoing clinical evaluation as therapeutic antitumor agents. Bioorg Med Chem Lett 2002;12:2893-2897. 68 Natarajan M, Roldan FA, Sherrif H et al. EKB-569 attenuates radiation triggered survival response in squamous cell carcinoma. Proc Am Soc Clin Oncol 2003;22:993a. 69 Herbst RS, Maddox AM, Rothenberg ML et al. Selective oral epidermal growth factor receptor tyrosine kinase inhibitor ZD1839 is generally well-tolerated and has activity in non-small-cell lung cancer and other solid tumors: results of a phase I trial. J Clin Oncol 2002;20:3815-3825. 70 Baselga J, Rischin D, Ranson M et al. Phase I safety, pharmacokinetic, and pharmacodynamic trial of ZD1839, a selective oral epidermal growth factor receptor tyrosine kinase inhibitor, in patients with five selected solid tumor types. J Clin Oncol 2002;20:4292-4302. 71 Ranson M, Hammond LA, Ferry D et al. ZD1839, a selective oral epidermal growth factor receptor-tyrosine kinase inhibitor, is well tolerated and active in patients with solid, malignant tumors: results of a phase I trial. J Clin Oncol 2002;20:2240-2250. Small-Molecule EGFR-TKIs 72 Albanell J, Rojo F, Averbuch S et al. Pharmacodynamic studies of the epidermal growth factor receptor inhibitor ZD1839 in skin from cancer patients: histopathologic and molecular consequences of receptor inhibition. J Clin Oncol 2002;20:110-124. 73 Negoro S, Nakagawa K, Fukuoka M et al. Final results of a phase I intermittent dose-escalation trial of ZD1839 (‘Iressa’) in Japanese patients with various solid tumours. Proc Am Soc Clin Oncol 2001;20:1292a. 74 Fukuoka M, Yano S, Giaccone G et al. Multi-institutional randomized phase II trial of gefitinib for previously treated patients with advanced non-small-cell lung cancer. J Clin Oncol 2003;21:2237-2246. 75 Kris MG, Natale RB, Herbst RS et al. A phase II trial of ZD1839 (‘Iressa’) in advanced non-small cell lung cancer (NSCLC) patients who failed platinum- and docetaxel-based regimens (IDEAL 2). Proc Am Soc Clin Oncol 2002;21:1166a. 76 Baselga J, Yano S, Giaccone G et al. Initial results from a phase II trial of ZD1839 (IRESSA) as second- and third-line monotherapy for patients with advanced non-small-cell lung cancer (IDEAL 1). Clin Cancer Res 2001;7(suppl):630a. 77 Cohen EE, Rosen F, Stadler WM et al. Phase II trial of ZD1839 in recurrent or metastatic squamous cell carcinoma of the head and neck. J Clin Oncol 2003;21:1980-1987. 78 Baselga J, Albanell J, Ruiz A et al. Phase II and tumor pharmacodynamic study of gefinitib (ZD1839) in patients with advanced breast cancer. Proc Am Soc Clin Oncol 2003;22:24a. 79 Johnson DH, Herbst R, Giaccone G et al. ZD1839 (´Iressa´) in combination with paclitaxel & carboplatin in chemotherapy-naive patients with advanced non-small-cell lung cancer (NSCLC): results from a phase III clinical trial (INTACT 2). Ann Oncol 2003;13:127. 80 Giaccone G, Johnson DH, Manegold C et al. A phase III clinical trial of ZD1839 (´Iressa´) in combination with gemcitabine and cisplatin in chemotherapy-naive patients with advanced non-small-cell lung cancer (INTACT 1). Ann Oncol 2003;13:2. 81 Cappuzzo F, Gregorc V, Rossi E et al. Gefitinib in pretreated non-small-cell lung cancer (NSCLC): analysis of efficacy and correlation with HER2 and epidermal growth factor receptor expression in locally advanced or metastatic NSCLC. J Clin Oncol 2003;21:2658-2663. 82 Bailey LR, Kris M, Wolf M et al. Tumor EGFR membrane staining is not clinically relevant for predicting response in patients receiving gefitinib (‘Iressa’, ZD1839) monotherapy for pretreated advanced non-small-cell lung cancer: IDEAL 1 and 2. Proc Am Assoc Cancer Res 2003;44:LB-170a. 83 Zakowski MF, Miller VA, Kris MG et al. Bronchioalveolar histology and smoking history predict response to gefitinib. Proc Am Soc Clin Oncol 2003;22:2524a. 84 Hidalgo M, Siu LL, Nemunaitis J et al. Phase I and pharmacologic study of OSI-774, an epidermal growth factor receptor tyrosine kinase inhibitor, in patients with advanced solid malignancies. J Clin Oncol 2001;19:3267-3279. 85 Perez-Soler R, Chachoua A, Huberman M et al. A phase II trial of the epidermal growth factor receptor (EGFR) Janmaat, Giaccone 586 tyrosine kinase inhibitor OSI-774, following platinum-based chemotherapy in patients (pts) with advanced, EGFRexpressing, non-small cell lung cancer (NSCLC). Proc Am Soc Clin Oncol 2001;20:1235a. 96 Adams VR, Bence AK, Anderson EB et al. A phase I pharmacokinetic/pharmacodynamic study evaluating multiple doses of oral GW572016 in healthy subjects. Proc Am Soc Clin Oncol 2003;22:374a. 86 Finkler N, Gordon A, Crozier M et al. Phase 2 evaluation of OSI-774, a potent oral antagonist of the EGFR-TK in patients with advanced ovarian carcinoma. Proc Am Soc Clin Oncol 2001;20:831a. 97 DeSimone PA, Bence AK, Anderson EB et al. A phase I study to investigate the safety, tolerability, and pharmacokinetics of single oral escalating doses of GW572016 in healthy volunteers. Proc Am Soc Clin Oncol 2003;22:275a. 87 Senzer NN, Soulieres D, Siu LL et al. Phase 2 evaluation of OSI-774, a potent oral antagonist of the EGFR-TK in patients with advanced squamous cell carcinoma of the head and neck. Proc Am Soc Clin Oncol 2001;20:6a. 98 Burris HA, Taylor C, Jones S et al. A phase I study of GW572016 in patients with solid tumors. Proc Am Soc Clin Oncol 2003;22:994a. 88 Bulgaru AM, Mani S, Goel S et al. Erlotinib (Tarceva): a promising drug targeting epidermal growth factor receptor tyrosine kinase. Expert Rev Anticancer Ther 2003;3:269-279. 89 Ratain MJ, George CM, Janisch L et al. Phase I trial of erlotinib (OSI-774) in combination with gemcitabine (G) and cisplatin (P) in patients with advanced solid tumors. Proc Am Soc Clin Oncol 2002;21:2115a. 90 Forero L, Patnaik A, Hammond LA et al. Phase I, pharmacokinetic (PK) and biologic study of OSI-774, a selective epidermal growth factor receptor (EGFR) tyrosine kinase (TK) inhibitor in combination with paclitaxel and carboplatin. Proc Am Soc Clin Oncol 2002;21:1908a. 91 Forouzesh B, Hidalgo M, Takimoto C et al. Phase I, pharmacokinetic (PK), and biological studies of the epidermal growth factor-tyrosine kinase (EGFR-TK) inhibitor OSI-774 in combination with docetaxel. Proc Am Soc Clin Oncol 2002;21:81a. 92 Saltz L, Kies MS, Abbruzzese JL et al. The presence and intensity of the cetuximab-induced acne-like rash predicts increased survival in studies across multiple malignancies. Proc Am Soc Clin Oncol 2003;22:817a. 93 Clark GM, Pérez-Soler R, Siu LL et al. Rash severity is predictive of increased survival with erlotinib HCl. Proc Am Soc Clin Oncol 2003;22:786a. 99 Belanger M, Jones CM, Germond C et al. A phase II, openlabel, multicenter study of GW572016 in patients with metastatic colorectal cancer refractory to 5-FU in combination with irinotecan and/or oxaliplatin. Proc Am Soc Clin Oncol 2003;22:978a. 100 Kaplan EH, Jones CM, Berger MS. A phase II, open-label, multicenter study of GW572016 in patients with trastuzumab refractory metastatic breast cancer. Proc Am Soc Clin Oncol 2003;22:981a. 101 DeBono JS, Schwartz G, Monroe P et al. Phase I and pharmacokinetic (PK) study of oral GW572016, a potent reversible dual inhibitor of both erbB1 and erbB2 tyrosine kinase (TK), administered in combination with capecitabine. Proc Am Soc Clin Oncol 2003;22:901a. 102 Spector NL, Reafsky E, Hurwitz H et al. Safety, clinical efficacy, and biologic assessments from EGF10004: a randomized phase IB study of GW572016 for patients with metastatic carcinomas overexpressing EGFR or erbB2. Proc Am Soc Clin Oncol 2003;22:772a. 103 Hidalgo M, Erlichman C, Rowinsky EK et al. Phase 1 trial of EKB-569, an irreversible inhibitor of the epidermal growth factor receptor (EGFR), in patients with advanced solid tumors. Proc Am Soc Clin Oncol 2002;21:65a. 94 Zinner RG, Nemunaitis J, Donato NJ et al. A phase I clinical and biomarker study of the novel pan-erB tyrosine kinase inhibitor CI-1033, in patients with solid tumors. Clin Cancer Res 2001;7:566a. 104 Morgan JA, Bukowski RM, Xiong H et al. Preliminary report of a phase 1 study of EKB-569, an irreversible inhibitor of the epidermal growth factor receptor (EGFR), given in combination with gemcitabine to patients with advanced pancreatic cancer. Proc Am Soc Clin Oncol 2003;22:788a. 95 Nemunaitis J, Eiseman I, Cunningham A et al. A phase 1 trial of CI-1033, a pan-erbB tyrosine kinase inhibitor, given daily for 14 days every 3 weeks, in patients with advanced solid tumors. Proc Am Soc Clin Oncol 2003;22:974a. 105 Salazar R, Kohne CH, Tabernero J et al. A phase 1/2A openlabel study of EKB-569 in combination with CPT-11/5FU/LV (FOLFIRI) in patients with advanced colorectal cancer. Proc Am Soc Clin Oncol 2003;22:888a.