Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
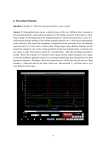
Quantitative Methods in Neuroscience/NB 545 Project Summary Purpose Random Walk is a relatively simple model for the excitatory and inhibitory steps that lead to a neuron firing. In the model, for each time period dt, the cell receives an input, determined randomly to be excitatory or inhibitory. If the input is excitatory, the cell's membrane potential increases (becomes more positive), while if the input is inhibitory, the membrane potential decreases (becomes more negative). Because the sign of the input is random, the membrane potential is randomly displaced upward or downward for each time step. A threshold is such that whenever the membrane potential reaches that point, the cell fires an action potential. Afterward, the membrane potential is reset to its starting value. In the random walk model, only a single factor, the membrane potential, determines whether or not the cell will fire an action potential. However, in a real cell, many other factors will influence the process. One such factor is the stochastic nature of the opening and closing of the voltage-gated sodium channels responsible for generating the rising phase of the action potential. The purpose of this project is to explore the effect of this stochastic process on the firing of an action potential in the random walk model. Part 1: Single Channel I will begin by examining the simplest case I can think of: a cell which has a single channel. For the sake of this simple model, the neuron will fire an action potential if the channel is open, and will not fire if the channel is closed. The opening and closing of the channel is stochastic, and I will think of the process as the flipping of a weighted coin, where heads means the channel will be open for that dt, and tails means it will be closed. Figure 1A single channel opens and closes randomly Of course, the channels we are interested in do not simply open and close as a function of time alone. They are voltage dependent. This voltage dependence can be simulated as the flip of a weighted coin, the weighting of which depends on the voltage. In the Random Walk script, we now have an extra step for each dt. We decide whether to take a step up or down, then decide whether or not to open or close the channel based on the new membrane potential. If the channel opens, we get a spike, and the membrane potential resets. If we assign a probability P(open) = 1 when at the level we called threshold earlier, and P(open) = 0 whenever we are below that level, we recover the same sort of random walk as the original. However, we can also assign a voltage dependence that varies over time according to some equation.. The question of what equation to use to determine voltage dependent probabilities of opening is problematic. We would like something that approaches 1 in the positive direction, and 0 in the negative direction, and hopefully will move between them over some interval that is interesting in terms of the model (between 0 and 10 in this case, since we have defined the resting potential as 0 and exB threshold as 10). Equations of the form y= give sigmoidal curves, approaching 0 and 1. 1+exB We can try various values of B until the curve fits the region of interest: 6.5 turns out to be pretty good. We can now run the RandomWalk simulation with a single channel. Note that although the channel is more likely to open near the former threshold line than at lower potentials, it frequently fires at other membrane potentials. Figure 2 Stochastic opening of a channel during a RandomWalk. a)path of random walk, with spikes shown in red. Red horizontal line represents original threshold. b)Histogram of interspike intervals. c) Histogram of level at which spikes occured (apparent threshold). Note variability of opening time. Part 2: Adding Another Channel The next thing we would like to do is move away from the excessively simplified model of only one channel. The addition of a second channel is a good time to think about another complexity of real voltage-gated sodium channels. These channels do not actually have simple binary, open/closed states: they can also be in an inactivated state. Let's assume that the channels never go directly from closed to inactivated, but that they always follow the sequence closed --> open --> inactive. Further, we know that there is a time course of inactivation, so let's assume that once a channel inactivates, it stays inactivated for a period of time, say, 2dt. How does this affect our model? I will add this into the code from this point on. Adding a second channel requires that we start thinking about how a population of channels can control the cell's firing rate. For example, do we still need just one channel open to give a spike, or do we need both? Let's look at both cases. Both channels required to give a spike: If we decide that both channels are required to generate a spike, then the probability of getting a spike is just the probability of both channels opening on the same dt. Anytime both open, we get a spike and reset to the resting potential. Figure 3. RandomWalk with two channels, in which both channels must open to get a spike. One channel open results in a spike: If instead we decide that only one of our two channels needs to be open to get a spike, then the probability of a spike at a given time is the probability that either channel is open at that time. Figure 4. RandomWalk with two channels, in which only one must open to get a spike. Note frequency of low apparent thresholds Note the differences in the potential at time of firing histograms. In the second case, the spikes always came at potentials below our original threshold, frequently much lower, because the probability of either channel being open is much greater. Part 3: More Complex Cases Of course, most cells are going to need more than two channels to generate an action potential, so we'd like to add many more to our model. We can construct a matrix where the rows represent channels, and each column holds the open/closed/inactive states of each channel. For the first dt, we assume all of the channels are closed. At the first dt, we take a step up or down, then evaluate the voltage dependent probability equation to obtain the probability that the channels will open. Then, we again generate random numbers to determine the states of the channels. We repeat this for each dt, and at each time point, count the number of channels that are in the open state. When it exceeds the limit we set, we achieve an action potential. We can adjust the number of channels, as well as the proportion of available channels necessary to get a spike. On making such changes over many trials, a very clear trend becomes apparent. If the number of channels required for an action potential is much less than the number available, the apparent threshold (the potential at which the cell most often spikes) becomes much more variable, and decreases significantly. This is because as the proportion required decreases, the likelihood of getting enough open channels increases. Figure 5. RandomWalk with 20 channels, 19 of which must open to get a spike Figure 6. RandomWalk with 20 channels, 9 of which must open to get a spike