Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
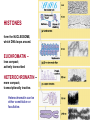
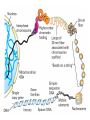
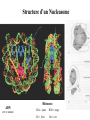
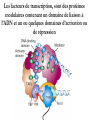
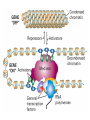
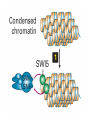
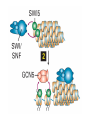
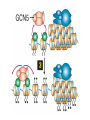
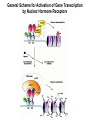
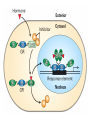
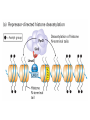
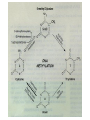
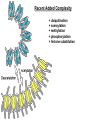
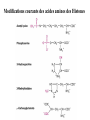
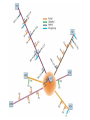
The Human Genome ~ 32,000 Genes ?? 200,000 Proteins a) alternative mRNA splicing b) post-translational modifications Régulation Transcriptionnel Pourquoi est-ce que l’expression génique est régulée ? Pourquoi la transcription est-elle le mode primaire de la régulation de l’expression d’un gène? Quel est le principal moyen de régulation de la transcription ? Comment est-ce que ceci est réalisé ? Le but principal du control de l’expression des gènes chez les organismes pluricellulaires est l’exécution des décisions précises au bon moment et dans les cellules appropriés au cours du développement et de la différentiation cellulaire. Regulation Can Be at Several Different Levels GENOME TRANSCRIPTOME Nucleus Gene Expression: Multiple, Spatially and Temporally Distinct Steps Carried out by Distinct Cellular Machinery DNA Pol II transcription PROTEOME Cytoplasm Primary RNA transcript Protein Nuclear processing Capping Splicing Polyadenylation Translation Mature mRNA Mature mRNA Export Degradation Post-Translational Modifications Degradation Gene expression is regulated at several levels : 1) Transcriptional – the most common way of controlling levels of a transcript (and therefore also the protein it encodes) 2) Post- transcriptional – regulation of aspects of the RNA after it is transcribed, for example, alternative splicing, polyadenylation, capping, transport out of the nucleus, and half- life 3) Translational – factors that affect the efficiency with which an mRNA is translated, such as mRNA capping, tRNA abundance, and modification of initiation factors (eIF- 2phosphorylation for example) 4) Post- translational – regulation at the protein level, mechanisms include protein stability,Covalent modification of the protein (Phosphorylation, Acetylation, Methylation, Ubiquitination, Sumoylation…), protein localization and protein degradation Le control de la transcription est le principal moyen de régulation de l’expression de gènes DNA Condensation 2 mètres d’ADN par cellule dans 2 x 10-9 cm3 (ou millilitres) 5 x 1010 kilomètres d’ADN par être humain Euchromatin open, transcriptionally active Heterochromatin condensed transcriptionally silent HISTONES form the NUCLEOSOME, which DNA loops around. EUCHROMATIN – less compact; actively transcribed HETEROCHROMATIN – more compact; transcriptionally inactive. Heterochromatin can be either constitutive or facultative. Levels of Chromatin Assembly Inactive Solenoid 30 nM structure Dnase I insensitive Active Beads-on-a-string structure Dnase I sensitive 10-fold Open Nucleosome-free Dnase I hypersensitive 100-fold Chromatin Structure • Core particle - 146-147 bp • Wrapped twice around histone octamer • H2A, H2B, H3, H4 • Linker DNA ~15 - 55 bp • H1 histone - linker associated Modèle d’empaquetage de la chromatine Structure d’un Nucleasome ADN vert et marron Histones : H2A = jaune H2B = rouge H3 = bleu H4 = vert Components of the Transcriptional Machinery • RNA Polymerase • Basal transcription factors • Sequence-specific transcription factors • Co-activators/repressors bridging the basal machinery and the other sequence-specific regulators • Enzymes that covalently modify core histones and other chromatin components • Enzymes that alter chromatin structure in an ATP-dependent manner Basal Gene Expression Pre-initiation complex assembly • Consensus cis-acting elements TATA box Initiator Element CCAAT box • General transcription factors TATA Box binding Protein (TBP) TBP Associated Factors (TAFs) • Mode of assembly Holoenzyme Sequential assembly Basal, Very Low Level mRNA Expression i) Detectable with RT-PCR ii) No protein production TAFs TBP Gene X -35 Basal Transcription Regulated Transcription 1. Cis-acting sequences 2. Transcriptional activators/co-activators 2,000 + transcription factors Combinatorial complexity Transcription Factors 1. Proteins (and RNA’s?) that regulate (positively or negatively) transcription initiation 2. Can act via sequence-specific DNA-protein interactions, or via protein-protein interactions 3. Many transcription factors bind to cis-acting regulatory DNA sequences (regulatory elements in DNA) associated with genes 4. Includes: general transcription factors, upstream activators, enhancer-binding proteins, cell-type-specific factors, co-activators, etc. Les facteurs de transcription, qui stimulent ou répriment la transcription, se fixent à des éléments régulateurs proches de du promoteur et des amplificateurs Les facteurs de transcription, sont des protéines modulaires contenant un domaine de liaison à l’ADN et un ou quelques domaines d’activation ou de répression Le rôle principal des activateurs et des répresseurs de la transcription eucaryotes consiste à se fixer à des complexes à sous unités multiples que régulent la transcription soit en modulant la structure de la chromatine (effet indirect) soit en interagissant avec la polymérase II et les facteurs généraux de la transcription (effet direct) Co-activator protein General transcription factors TBP Gene X Transcriptional activators binding to promoter region TATA -35 Regulated Transcription Les activités de nombreux facteurs de la transcription sont régulées directement par son interaction avec des hormones et indirectement par la fixation de protéines et de peptides extracellulaires à des récepteurs présentes à la surface des cellules. Ces protéines constituent donc des cibles moléculaires pour des médicaments agissant comme agonistes ou antagonistes afin de réguler l’activité cellulaire à travers l’expression génique. Les récepteurs nucléaires constituent une superfamille de facteurs transcriptionnels dimériques à doigt à zinc C4 qui fixent des hormones liposolubles et interagissent avec des éléments spécifiques de réponses dans l’ADN. La liaison des hormones aux récepteurs nucléaires induit des changements conformationnels qui modifient leur interaction avec d’autres protéines Les récepteurs nucléaires hétérodimériques (récepteurs de l’acide rétinoïque, de la vitamine D ou de l’hormone thyroïdienne) sont présents uniquement dans le noyau. En l’absence d’hormone ils répriment la transcription de gènes cibles, une fois leur ligands fixés ils activent la transcription. General Scheme for Activation of Gene Transcription by Nuclear Hormone Receptors Les récepteurs des hormones stéroïdes sont des récepteurs nucléaires homodimériques. En l’absence d’hormone, ils sont piégés dans le cytoplasme par des protéines inhibitrices. Une fois associés a leur ligands, ils peuvent subir une translocation vers le noyau et activer la transcription des gènes cibles. Epigenetics “Any heritable changes in gene expression (influencing on gene function) that occur without a change in DNA sequence” Such changes cannot be attributed to changes in DNA sequence (mutations) but they can be as permanents as mutations (difficult to reverse) Major mechanisms of epigenetic changes Three important factors that play clear roles in transcriptional regulation are known: – DNA METHYLATION – A subset of cytosine (C) residues could be modified by methylation. – HISTONE ACETYLATION Histones can be modified by acetylation. - IMPRINTING (non-coding RNAs involved) Histone acetylation and gene expression • HISTONES in transcriptionally active genes are often ACETYLATED Acetylation of lysine residues in histones : – Reduces positive charge, weakens the interaction with DNA. – Makes DNA more accessible to RNA polymerase II • Enzymes that ACETYLATE HISTONES are recruited to actively transcribed genes. • Enzymes that remove acetyl groups from histones are recruited to methylated DNA. There are additional types of histone modification as well, such as methylation of the histones. Histone Acetyltransferase (HAT) Complexes P300/CBP PCAF TAFII 250 • Conserved lysines in the N-terminal tails of histones • Post-translational - Reversible • Localized - ~2 nucleosomes • Can also acetylate other proteins involved in transcription Histone Deacetylation Complexes (HDAC) Histone acetylation² Histone acetyltransferase Histone deacetylase Hypoacetylation Strong internucleosomal interactions: Acetylation has two functions: Neutralize the positive charge on the lysine residues Destabilize interactions between histone tails and structural proteins Hyperacetylation (Yellow) Weak internucleosomal interactions: histone tails do not constrain DNA, which is accessible to transcription factors Mitosis does not erase acetylation, but merely distributes histones, between the daughter chromosomes. Specific acetyltransferases (red) end up distributed between the daughter chromosomes, too. Once segregated, an acetyltransferase would acetylate the adjacent nucleosomes (yellow) and thereby spread over the entire chromatin domain. DNA Methylation • Genes that are transcriptionally inactive are often METHYLATED. – In eukaryotes, cytosine residues are modified by methylation. • Typically, the sites of methylation are CpG dinucleotides (vertebrates); CpG islands in front of genes are mostly unmethylated DNA Methylation • 5-methylcytosine 5’-CpG-3’ • CpG only 10% of predicted frequency in eukaryotic genomes • Deaminated methylcytosine - to - thymine • ~70% of remaining CpGs are methylated often in repeat sequences irreversibly repressed state Mitosis erases methylation only temporarily Cytosine methyl-transferases of vertebrates have preference for hemi-methylated targets Because of that newly synthetized DNA strand will receive same methylation pattern as parental strand DNMT1 enzyme = maintain methylation DNMTs coordinate transcriptional repression with histone deacetylases (HDACs) and methyl-CpG binding proteins (MBDs) DNMT-mediated gene silencing Transcriptional Repression by Methylation Binding to methylated CpGs by Methyl CpG binding proteins (MeCP2/MBD1,2,3) Mutations of MeCP2 in humans - Rett Syndrome (similar neurologic phenotype in KO mice) Changes in DNA methylation during mammalian development DNA methylation and histone acetylation can be maintained through replication This allows the packing of chromatin to be passed on - just like a gene sequence. – However, differences in chromatin packing are not as stable as gene sequences. • Heritable but potentially reversible changes in gene expression are called EPIGENETIC phenomena – Vertebrates use these differences in chromatin packing to IMPRINT certain patterns of gene regulation. – Some genes show MATERNAL IMPRINTING while other show PATERNAL IMPRINTING. • The alleles of some genes that are inherited from the relevant parent are methylated, and therefore are not expressed. Recent Added Complexity + ubiquitination + sumoylation + methylation + phosphorylation + histone substitution Modifications courants des acides amines des Histones Histone Substitutions H2AZ H2AX H3.3 Variations of nucleosome stability The Increasingly Complex Code Location specific Quantitative and Qualitative Epigenetic Modification Chromatin Remodeling Complexes SWI/SNF Family • Facilitates gene activation by assisting transcription machinery to gain access to targets in chromatin. • Multi-subunit complex - ~10 units • Very low abundance ~150 complexes/cell • Molecular weight - 2 MDa • Destabilize nucleosomes/ disrupt DNA histone contacts • ATP- dependent activity Chromatin Remodeling Complexes SWI/SNF Family Related to a Helicase superfamily of proteins RAD16/RAD54/ERCC6 - DNA Recombination/Repair SWI2/SNF2/brahma - Transcription ATR-X Syndrome Mutations of Helicase - XH2 Alpha Thalassemia + Mental Retardation Chromatin Modification Chromatin Remodeling SNF/SWI Transcription Factor Modification Acetylation Phosphorylation DNA Methylation CpG dinucleotides MeCP2 Histone Substitution H2AZ H2Ax H3.3 Histone Modification Acetylation Ubiquitination Sumoylation Methylation Phosphorylation Solid evidence demonstrates that in addition to genetic alteration, aberrant epigenetic regulation, such as silencing of tumor suppressors, is used by cancer cells to escape growth and death control mechanisms EPIGENETIC MODIFICATIONS AND CANCER Epigenetic modifications and particularly the methylation of cytosines 5’ of guanine residues (CpGs) in gene promoter regions is an essential regulatory mechanism for normal cell development. DNA methylation can inactivate tumor suppressor genes by inducing C>T transitions in somatic and germline cells and by altering gene transcription. On the other hand, hypomethylation of specific sequences may reactivate the expression of potential oncogenes. Thus, aberrant hyper- and hypomethylation are considered crucial steps leading to cancer development. Thus, compounds able to influence the epigenetic status of a cell have promise for cancer treatment. DNA metyl transferase inhibitors (DNMT-Is) as Decitabineand Azacitadine have been already largely tested in cancers. (One of the most recent results obtained 30–60% response rates in leukemias) Histone deacetylase inhibitors (HDAC-Is) as SAHA, valproic acid, MS-275, Depsipeptide and phenylbutyrate, exhibit impressive anti-tumor activity potentiated by little toxicity in vitro, ex vivo and in vivo models DNMT-I HDAC-I Epigenetic Therapy: Restoration of Gene Function Silenced Gene Function Evidence of restoration of function by epigenetic therapy ER Estrogen Receptor Restored expression from estrogen sensitive genes RARB2 Retinoic acid receptor Restored growth suppression by retinoic acid MLH1 Mismatch repair Correction of mismatch repair defect P16/INK4a cell-cycle regulation Restoration of RB checkpoint P14/ARF Restored normal cellular distribution of MDM2 MDM2 inhibition Hypomethylating Cytosine Analogues An important component of the actions spectrum of HDAC-Is is the induction of the cyclin-dependent kinase inhibitor p21WAF1/CIP1, Very recent data point to an exciting potential of HDAC-Is that may explain the selective anti-cancer activity of these compounds in AML models : the induction of TRAIL/Apo2L and its death receptors. (activation of the TRAIL death pathway is well known to be more toxic for tumor cells than for normal cells.) HDAC Inhibitors Clinical Trials • • • • • Butyrate, Phenylbutyrate Depsipeptide SAHA MS-275 Valproic acid Studies on the molecular basis that modulate epigenetic events during tumorigenesis, and their effect on differentiation and apoptosis pathways, alone or in combination with other drugs, will provide new tools to fight cancer. Régulation Transcriptionnel Pourquoi est-ce que l ’expression génique est régulée ? Pour faire les protéines appropriées dans les bonnes cellules au bon moment. Pourquoi la transcription est-elle le mode primaire de la régulation de l’expression d’un gène? Pour obtenir un meilleur control avec une efficacité maximale. Quel est le principal moyen de régulation de la transcription ? « Turning on-and-off » l’amorçage de la transcription. Comment est-ce que ceci est réalisé ? Par des protéines que se lient à l'ADN et qui augmentent ou répriment la liaison de la ARN polymérase au promoteur Par des modifications de la chromatine