Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Extracellular matrix molecules of perineuronal nets –
Studies on structure and function in synapse formation
and synaptic activity
Dissertation to obtain the degree
Doctor rerum naturalium (Dr. rer. nat.)
at the Faculty of Biology and Biotechnology
International Graduate School of Biosciences
Ruhr-University Bochum
Department of Cell Morphology and Molecular Neurobiology
submitted by
Maren Geißler
1st supervisor: Prof. Dr. Andreas Faissner
2nd supervisor: Prof. Dr. Dr. Dr. Hanns Hatt
Bochum, February 2012
Extrazelluläre Matrix Moleküle perineuronaler Netze Strukturelle und funktionelle Untersuchungen zur
Synapsenbildung und synaptischer Aktivität
Dissertation zur Erlangung des Grades
eines Doktors (Dr. rer. nat.) der Naturwissenschaften
der Fakultät für Biologie und Biotechnologie
an der Internationalen Graduiertenschule Biowissenschaften
der Ruhr-Universität Bochum
Lehrstuhl für Zellmorphologie und Molekulare Neurobiologie
angefertigt von
Maren Geißler
Referent: Prof. Dr. Andreas Faissner
Korreferent: Prof. Dr. Dr. Dr. Hanns Hatt
Bochum, im Februar 2012
"Wenn das Gehirn des Menschen so einfach wäre, dass
wir es verstehen könnten, dann wären wir so dumm,
dass wir es trotzdem nicht verstehen könnten."
Jostein Gaarder, in „Sofies Welt“
Table of content
Table of content
Chapter 1 ............................................................................................................................. 1
General Introduction .......................................................................................................... 1
1.1 The Chemical Synapse ................................................................................................................................ 3
1.1.1 Synaptogenesis ...................................................................................................................................... 6
1.1.2 Synapse Maturation and Pruning................................................................................................... 12
1.1.4 Synaptic plasticity ............................................................................................................................... 14
1.2 Astrocytes...................................................................................................................................................... 15
1.2.1 Astrocytes - a changing image ....................................................................................................... 16
1.2.3 Neuron-glia interaction - the tripartite synapse ....................................................................... 16
1.3 The extracellular matrix ........................................................................................................................... 19
1.3.1 The composition of the brain´s ECM............................................................................................. 19
1.3.2 The “tetrapartite Synapse” .............................................................................................................. 26
1.3.3. The quadruple knock-out mouse .................................................................................................. 27
1.3.4 Perineuronal nets ................................................................................................................................ 28
1.5. References .................................................................................................................................................... 33
Chapter 2 ........................................................................................................................... 46
Objectives.......................................................................................................................... 46
Chapter 3 ........................................................................................................................... 48
Primary hippocampal neurons, which lack four crucial extracellular matrix
molecules, display abnormalities of synaptic structure and function and severe
deficits in perineuronal net formation ........................................................................... 48
3.1 Abstract .......................................................................................................................................................... 48
3.2 Introduction ................................................................................................................................................... 49
3.3 Material and methods ................................................................................................................................ 52
3.3.1 Ethical standards and animal housing ......................................................................................... 52
I
Table of content
3.3.2 Immunological reagents ................................................................................................................... 52
3.3.3 Cell culture ............................................................................................................................................ 53
3.3.4 Electrophysiology................................................................................................................................ 54
3.3.5 Immunocytochemistry....................................................................................................................... 55
3.3.6 Western Blotting.................................................................................................................................. 56
3.3.7 Microscopy ............................................................................................................................................ 56
3.3.8 Quantifications and statistical analyses ...................................................................................... 57
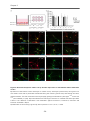
3.4 Results ............................................................................................................................................................ 58
3.4.1 Primary quadruple knock-out neurons and the extracellular matrix expression
pattern ............................................................................................................................................................... 58
3.4.2 Reduced frequency of mPSCs in patch clamp recordings .................................................... 61
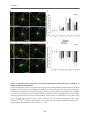
3.4.3 Synapse formation in the indirect neuron-astrocyte co-culture assay ............................ 64
3.4.4 Quantitative protein analysis of GAD 65, GAD 67 and vGlut ................................................ 67
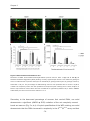
3.4.5 PNN formation in primary hippocampal neurons lacking four matrix components .....69
3.4.6 Synapse formation on PNN wearing neurons............................................................................ 71
3.5 Discussion ...................................................................................................................................................... 74
3.6 Acknowledgements .................................................................................................................................... 78
3.7 References ..................................................................................................................................................... 79
Chapter 4 ........................................................................................................................... 86
A new indirect co-culture set up of mouse hippocampal neurons and astrocytes on
microelectrode arrays ..................................................................................................... 86
4.1 Abstract .......................................................................................................................................................... 86
4.2 Introduction ................................................................................................................................................... 87
4.3 Material and methods ................................................................................................................................ 89
4.3.1 Ethical standards ................................................................................................................................ 89
4.3.2 Animal housing .................................................................................................................................... 89
4.3.3 Cell culture ............................................................................................................................................ 89
4.3.4 Microelectrode Array recordings ................................................................................................... 91
II
Table of content
4.3.5 Immuncytochemistry ......................................................................................................................... 93
4.3.6 Microscopy ............................................................................................................................................ 94
4.3.7 Plating efficiency and cell death .................................................................................................... 94
4.3.8 Statistics ................................................................................................................................................ 95
4.4 Results ............................................................................................................................................................ 95
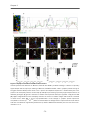
4.4.1 The indirect neuron astrocyte co-culture set up ...................................................................... 95
4.4.2 Spontaneous activity ....................................................................................................................... 100
4.4.3 Bursting behavior ............................................................................................................................. 103
4.4.4 Bicuculline treatment ..................................................................................................................... 104
4.5 Discussion ................................................................................................................................................... 106
4.6 Acknowledgements ................................................................................................................................. 109
4.7 References .................................................................................................................................................. 110
Chapter 5 ......................................................................................................................... 113
5.1 Comprehensive Discussion and 0utlook........................................................................................... 113
5.2 References .................................................................................................................................................. 124
5.2 Summary ..................................................................................................................................................... 128
5.3 Zusammenfassung .................................................................................................................................. 131
5.4 List of Abbreviations................................................................................................................................ 135
Chapter 6 ......................................................................................................................... 138
Appendix ......................................................................................................................... 138
6.1 Erklärung..................................................................................................................................................... 138
6.2 Curriculum Vitae ....................................................................................................................................... 139
6.3 Publications and benchmark of contribution .................................................................................. 141
6.4 Conference participations and poster abstracts ........................................................................... 143
6.5 Danksagung................................................................................................................................................ 144
III
Chapter 1
General introduction
Chapter 1
General Introduction
The vertebrate central nervous system (CNS) is one of the most complex organs
originated during evolution. The fascinating structural and functional complexity of the
brain is not completely understood so far and there are still plenty of developmental
and functional questions to be addressed and solved in the future.
The modern neuroscience looks back to a long history and a big technological and time
consuming effort is made to unravel the secrets of the vertebrate brain. Probably, one
of the most important steps in the accumulation of our today´s knowledge was the
postulation of the “neuron doctrine”, achieved by Heinrich Wilhelm Waldeyer. He was
cited by Camillo Golgi in the Nobel Lecture he gave in December 1906:
"The nervous system is made up of innumerable nerve units (neurons), which are
anatomically and genetically independent of each other....”
[The neuron doctrine - theory and facts, Camillo Golgi, Nobel Lecture December 11, 1906]
With this postulate, Heinrich Wilhelm Waldeyer, Camillo Golgi, Santiago Felipe Ramón y
Cajal and other research fellows from this decade revolutionized the former concept of
the brain. Until that time, the brain was assumed as being an undefined reticular mass,
thought to present an exception of the cell theory. With the neuron doctrine, a novel
picture of the anatomy and the physiology of the brain emerged:
“…The transmission of nerve impulses is conducted from the protoplasmic extensions
and the cell body towards the nerve extension; consequently, each nerve cell possesses a
receiving apparatus constituted by the body and the protoplasmic processes, a
conducting apparatus - the nerve process - and a transmitting or discharging organ…”
[The neuron doctrine - theory and facts, Camillo Golgi, Nobel Lecture December 11, 1906]
-1-
Chapter 1
General introduction
At these times, the neuroscience itself and the methods used were still in their infancy.
Nevertheless, the stainings drawn by the aforementioned pioneers of neuroscience bear
astonishing similarities to the today´s cellular and anatomical views of modern
neuroscience (see Fig. 1).
Figure 1: Drawings from the pioneers in neuroscience Camillo Golgi and Santiago Ramón y Cajal
A) Drawing of the neural circuitry of the rodent hippocampus by Santiago Ramón y Cajal. B: Drawing of a
cell from the granular layer of the cerebellum by Camillo Golgi; C: Astrocyte with processes connecting
the vasculature.
Taken from: “The neuron doctrine - theory and facts” (Camillo Golgi, Nobel Lecture December 11, 1906)
Years before the first detailed description of neurons as defined units of the CNS
emerged, glia cells where described for the first time by Rudolf Virchow in 1858 (see
Fig. 1C). At this stage, glia cells where assumed as kind of glue, holding the
protagonistic neurons of the brain together. The concept of the glia cells was probably
one of the most evolving pictures in the last decades, and the knowledge is still
growing. Once considered as being merely connective tissue, glia cells, especially
astrocytes are today known to be indispensable for neuronal survival and
communication. The neuronal picture has also rapidly matured and changed years after
the aforementioned neuronal postulate, due to the invention and the continuing
development in staining techniques and microscopy. Therefore, the progression in the
-2-
Chapter 1
General introduction
human knowledge about the CNS increased rapidly, and terms such as synapse,
perineuronal net, astrocytes, and extracellular matrix became manifested in the
modern neuroscience.
1.1 The Chemical Synapse
The chemical synapse is the pivotal communication element between neurons in the
CNS. Chemical synapses are the main type of synapses formed in the CNS. As the
name implies, chemical synapses are capable of converting an incoming electrical
signal (action potential) into a chemical signal (neurotransmitter release), which
becomes retranslated into an electrical signal (changes of the membrane potential) by
the input receiving cell. What was broken down here to a few words requires multiple
very complex and delicate processes, whose perturbation can lead to the breakdown of
the whole system and is mirrored in a couple of developmental diseases such as
schizophrenia or autism.
The synaptic transmission pathway is highly conserved throughout evolution and is
uniformly found from simple invertebrates to the much more complex human brain
(Kandel 2001; Ryan and Grant 2009). The chemical synapse implies anatomically three
specialized cell compartments. i) The presynaptic bouton, representing a small axonal
varicosity, enriched with neurotransmitter filled clear-centered vesicles. ii) The active
zone, within this presynaptic compartment, which is equipped with a unique set of
proteins enabling the fusion, the exocytosis and the recycling of these vesicles. iii) The
postsynaptic counterpart, directly facing the active zone and harboring a complex and
electron-dense network of specialized signal receiving and signal transducing proteins,
collectively named as postsynaptic density (PSD) (Palay 1956).
These structures become visible in electron microscopical images of glutamatergic
synapses, formed in vitro (see Fig.2) and in vivo. A synaptic cleft is formed between the
pre- and the postsynapse and defined cleft spanning proteins hold the active zone and
the PSD in register (Waites, Craig et al. 2005). Furthermore, the synaptic cleft is known
to harbor carbohydrates and extracellular matrix (ECM) molecules (Dityatev and
Schachner 2003) influencing the synaptic transmission (see chapter 1.3.2).
-3-
Chapter 1
General introduction
Both the pre- and the postsynaptic molecular scaffolds, represent very complex
cellular machineries with myriads of strictly regulated and organized proteins (Collins,
Husi et al. 2006) and only the orchestrated cellular events within these functional
compartments result in proper synaptic signaling:
An incoming action potential induces the fusion of synaptic vesicles with the
presynaptic membrane via local calcium elevations through the activation of voltagegated Ca2+ channels. The subsequent flood of the synaptic cleft with defined
transmitters and their following binding to postsynaptic receptors leads to the opening
of ligand-gated ion channels in the postsynaptic cell.
Figure 2: Electron-microscopical images, showing the ultrastructure of an excitatory glutamatergic
synapse.
Shown are two synapses formed between hippocampal neurons after 15 days in vitro (DIV). A A synapse
formed on a dendritic spine (SP). Arrowheads indicate the docked vesicles facing the postsynaptic
density. B Detailed view of a presynapse with synaptic vesicles (SVs) and vesicles docked to the active
zone (AZ). Stars indicate the electron dense protein assembly at the postsynaptic density (Waites, Craig
et al. 2005).
The most prominent excitatory transmitter is glutamate, while glycin and gammaaminobutyric acid (GABA) are the main inhibitory neurotransmitters. There exists also a
bunch of modulatory neurotransmitters like serotonin, acetylcholine, noradrenalin and
different
neuropeptides.
The
transmitter-induced
opening
of
the
respective
postsynaptic ion channels and the subsequent change of the ion homeostasis leads to a
change of the postsynaptic membrane potential, thereby in- or decreasing the
probability for a new action potential to be elicited.
-4-
Chapter 1
General introduction
The postsynaptic membrane comprises various ion channels, kinases, phosphatases,
signaling molecules and a diversity of receptors (see Fig. 3). Prominent postsynaptic
ligand-gated ion channels are the N-Methyl-D-aspartate (NMDA) and the alpha-amino3-hydroxyl-5-methyl-4-isoxazole-propionate (AMPA) receptors, both are necessary for
proper synaptic signaling. Nevertheless, NDMA receptor expressing and AMPA receptor
lacking, non-functional synapses can be found in CNS and occur frequently during
development. These synapses are called silent synapses (Faber, Lin et al. 1991;
Kerchner and Nicoll 2008) and become activated by the delayed insertion of AMPA
receptors, which become recruited to the membrane during development. These silent
synapses my present early tools for experience dependent synaptic plasticity (Kerchner
and Nicoll 2008).
Figure 3: The chemical synapse
Schematic drawing of a glutamatergic synapse with the presynaptic bouton, containing the synaptic
2+
vesicles, voltage-gated Ca channels and the exocytotic release machinery (not shown). The synaptic
cleft is flooded with neurotransmitter and harbors extracellular matrix molecules binding to a variety of
postsynaptic receptors and thereby influencing the postsynaptic response. Further, the inserted
glutamate receptors of the NMDA and AMPA type are indicated as being anchored in the postsynaptic
density (PSD) (Dityatev and Schachner 2003).
The postsynaptic receptors are anchored and coordinated within the membrane via
hundreds of different PSD proteins (Collins, Husi et al. 2006) (see Fig. 3). Prominent
PSD proteins are scaffold proteins like the membrane-associated guanylate kinase
-5-
Chapter 1
General introduction
(MAGUK) proteins as PSD95, multiple ankyrin repeat domains (Shank) family members,
guanylate kinase-associated protein (GKAP)-family members and the glutamate
receptor interacting proteins (GRIPs) (Feng and Zhang 2009). Most of the scaffolding
proteins found in the PSD contain PDZ domains, which are well suited for proteinprotein interactions and bind to receptors with weak affinities, enabling rapid changes
and plastic adaptations within the postsynaptic machinery (for review see Feng and
Zhang 2009).
The pre-, and especially the postsynapse are subject to plastic changes and due to its
fundamental importance for proper neuronal signaling and for the whole brain´s
function, it is necessary to understand the formation and malleability of synapses in
development and disease.
1.1.1 Synaptogenesis
In the human brain, one trillion synapses are assumed to be formed between
approximately hundred billion nerve cells. This network of enormous complexity has to
be build up and coordinated strictly, in order to obtain accurate neuronal function.
During embryogenesis neurons are born in the ventricular (VZ) and subventricular zone
(SVZ) through the division of precursor cells and this process is followed by the
migration to their final destination (Temple 2001). Along the way neurons are guided
by radial glia cells and gradual expression of extrinsic signaling molecules (Rakic and
Sidman 1970). In the following steps, axons reach out, from growth cones searching for
an appropriate synaptic partner, and start to establish connections with neighboring or
more remote neurons. Much knowledge about the formation of synapses has
accumulated from studies at the neuromuscular junction (Hall and Sanes 1993), but
the CNS synaptogenesis is a very complex process not completely understood so far.
A bunch of proteins is thought to be involved in the synaptic assembly, leading to a
functional synapse with a proper presynaptic transmitter release machinery and an
efficient postsynaptic recognition through the expression of the cognate receptors (for
review see Garner, Zhai et al. 2002).
-6-
Chapter 1
General introduction
The initial formation of synaptic connections is intensively studied in hippocampal
neurons (for review see Verderio, Coco et al. 1999), and the observed steps of
synaptogenesis in vitro are thought to resemble the processes occurring in vivo. Thus,
cultured hippocampal neurons represent a versatile tool to study the different steps of
synapse formation (Basarsky, Parpura et al. 1994; Verderio, Coco et al. 1999; Pyka,
Wetzel et al. 2011).
1.1.1.1 Initial Cell-Cell Contact
First, two cells have to find and face each other. These can be juxtaposed axonal or
dendritic membranes of two neighboring neurons, forming filopodia, or growth cones,
established during axonal pathfinding, searching for an appropriate partner over longer
distances (Vaughn 1989). One theory, how neurons encounter the correct partner, is
based on the idea of Roger Sperry, who postulated the “lock-key” theory (Sperry 1963),
meaning that two cells can assemble an initial cell-cell contact, if the expression of a
matching pair of a membrane bound ligand and a receptor is given. A prominent
subclass of proteins involved in cell-cell recognition is represented by the cell adhesion
molecules (CAMs). Examples of neuronal surface CAMs, thought to be involved in this
initial contact between two synapse-forming cells are the neurexins, binding to
neuroligins (Craig and Kang 2007), the ephrin-EphB complex (Kayser, Nolt et al. 2008;
Akaneya, Sohya et al. 2010), integrins (Einheber, Schnapp et al. 1996), family members
of the Ig superfamily, protocadherins and the
classical cadherins (Arikkath and
Reichardt 2008) facing a rich repertoire of alternatively spliced cadherin-related
neuronal receptors (CNRs). N-cadherin, as one of the most intensively studied family
members, becomes expressed when early synaptogenesis starts. N-cadherin is
selectively expressed at the nascent synapse and becomes clustered to the active
zones in matured synapses around 14 DIV (Elste and Benson 2006). Therefore, the
expression of cadherins seems to play a dual role in synapse formation as well as in
synapse function. The perturbation of the cadherin/catenin binding via the expression of
a dominant-negative construct leads to failures in synaptic structure and function
(Togashi, Abe et al. 2002). Nevertheless, N-cadherin is suggested to be supportive in
initial contact formation and stabilization between two synapse forming cells, rather
-7-
Chapter 1
General introduction
than being crucial for synapse formation per se (Waites, Craig et al. 2005). Ephrins and
Eph receptor are involved in many steps during the development of neuronal networks
(Wilkinson 2001). Thus, Eph receptor signals play crucial roles in the first steps of
axonal pathfinding and also in later stages during the morphological alteration of
filopodia to mature-shaped spines and clustering of NMDA receptors (Dalva, Takasu et
al. 2000; Wilkinson 2001). Ephs signal to multiple downstream effector molecules
reforming the cytoskeleton, such as focal adhesion kinases (Moeller, Shi et al. 2006)
and Rho family GTPases (Penzes, Beeser et al. 2003). Especially ephrinA5 and EphA5
are assumed to be crucially involved in early as well as late stages of synaptogenesis
(Akaneya, Sohya et al. 2010). Neuroligin is a further strong candidate to be crucially
involved in first steps in synapse formation (Scheiffele, Fan et al. 2000).
There exist a couple of priming molecules released during development, which are
thought to have synaptogenic activity, thus inducing cell-cell contacts with following
formation of a synapse. Amongst others these are molecules like Wnt, the fibroblast
growth factor (FGF) (Scheiffele 2003; Umemori, Linhoff et al. 2004) and neurotrophic
factors as brain derived neurotrophic factor (BDNF) (Alsina, Vu et al. 2001).
Once two cells have found each other, navigated by the expression of extracellular
guidance cues (which can be attractive or repellent) and the aforementioned cell
adhesion molecules, the future synapse starts to differentiate.
1.1.1.2 Presynaptic Assembly
Interestingly, neurons are assumed to be intrinsically biased to form synaptic contacts.
Thus, neurons growing on microisland of glia cells start to form cell own contacts,
called autapses, rather than remaining without contacts (Bekkers and Stevens 1991).
A diversity of proteins is involved in the assembly of the presynapse. For example,
neuroligin was shown to induce presynaptic differentiation (Scheiffele, Fan et al. 2000),
to be involved in active zone formation (Dean, Scholl et al. 2003) and to be a crucial
component in synaptic assembly in general (Lee, Dean et al. 2010; Sun, Xing et al.
2011).The synaptic cell adhesion molecule SynCAM is a further member, crucially
involved in the differentiation of the presynaptic side, and it is known to be expressed in
both synaptic partners (Scheiffele 2003). Synapsin I is an important regulator of
-8-
Chapter 1
General introduction
synapse formation, which adjusts the synapse number in response to extracellular
signals (Perlini, Botti et al. 2011).
For the differentiation of the presynapse, proteins of the presynaptic machinery have to
be transported to the presumptive site of synaptic contact. It is commonly accepted
that packages of presynaptic proteins are recruited via vesicular trafficking (see Fig. 4)
(Ahmari, Buchanan et al. 2000; Garner, Zhai et al. 2002; Ziv and Garner 2004;
McAllister 2007).
Figure 4: Synaptogenesis at CNS glutamatergic synapses
The first step in synapse formation involves the activation of cell adhesion molecules (CAMs), such as
cadherins or neuroligin/neurexin. (b) Pleomorphic vesicular clouds become clustered pre- and
postsynaptically at sites of cell–cell contact. Vesicles could carry the precursors of the active zone (c)
The electron-dense dense core of the 80 nm vesicles suggests that these might also deliver
synaptogenic factors that could help drive postsynaptic differentiation. (d) Differentiation of the
postsynaptic cell appears to occur by the sequential in situ recruitment of PSD scaffolding molecules
followed by glutamate receptors and PSD signaling molecules. Whether vesicular trafficking also plays a
role in delivering PSD scaffold proteins is not known. The time points (t) represent the approximate time
course of these processes in minutes after axo–dendritic contact (Garner, Zhai et al. 2002).
It was shown that there exist different types of protein-carrying vesicles for the
construction of the presynaptic release machinery and the active zone (Ahmari,
Buchanan et al. 2000; Tao-Cheng 2007; Bury and Sabo 2011). One distinguishes the
piccolo transport vesicles (PTV) and the synaptic vesicle protein transport vesicles
-9-
Chapter 1
General introduction
(STV) (Zhai, Vardinon-Friedman et al. 2001; Shapira, Zhai et al. 2003; Sabo, Gomes et al.
2006). Both types of vesicles can be classified electron-microscopically: PTVs are
80nm dense core vesicles, while vesicles, carrying proteins for the synaptic vesicles are
small clear-centered vesicles (Ahmari, Buchanan et al. 2000; Zhai, Vardinon-Friedman
et al. 2001).The 80nm dense core PTVs were shown to carry proteins important for the
assembly of the active zone (e.g. Bassoon and piccolo), proteins for the exocytotic
fusion machinery as well as synaptogenic factors, initializing the postsynaptic
differentiation upon release (Zhai, Vardinon-Friedman et al. 2001; Tao-Cheng 2007).
The clear STVs are packed with many SV-associated proteins as voltage-dependent
Ca2+ channels, synaptic vesicle protein 2, synapsin I and amphiphysin (Ahmari,
Buchanan et al. 2000) and other proteins crucial for the exocytosis and the recycling of
synaptic vesicles. STVs were shown to split and fuse occasionally during the transport
and recruitment processes (Ahmari, Buchanan et al. 2000; Bresler, Shapira et al. 2004).
Recently, it was shown, that both types of vesicles are transported in a coordinated
fashion and that both vesicles occur frequently at the nascent synapse (Bury and Sabo
2011). Thus, the presynaptic machinery seems to become aggregated within a defined
time-frame and in a pre-assembled manner (McAllister 2007).
There exist a few publications describing the formation of neurotransmitter filled
vesicles before active exocytosis into the synaptic cleft could be observed (Hannah,
Schmidt et al. 1999; Ahmari, Buchanan et al. 2000; Bury and Sabo 2011). Prior to
synaptogenesis, the non-regulated fusion of these vesicles at non-synaptic sites leads
to detectable levels of transmitter along growth cones (Hannah, Schmidt et al. 1999;
Sudhof 2000). In the nascent synapse “ready-to-use” packages exists, which comprised
preassembled components of the presynaptic machinery, which can function as
rudimentary synaptic specialization (Huttner, Ohashi et al. 1995; Ahmari, Buchanan et
al. 2000; Tao-Cheng 2007). Synapse formation is not completely dependent on
transmitter release (Verhage, Maia et al. 2000), but it is suggested that presynaptic
differentiation precedes postsynaptic differentiation (Friedman, Bresler et al. 2000). In
contrast to that, former studies did also suggest that the future postsynaptic
compartment can be involved in the first contact initiation and can actively search for a
presynaptic partner (Nimchinsky, Sabatini et al. 2002). Further, the target neuron may
- 10 -
Chapter 1
General introduction
induce the presynapse to become competent for synapse formation via protein
interactions or acting directly on the axon (Ullian, Christopherson et al. 2004; Umemori,
Linhoff et al. 2004)
1.1.1.3 Postsynaptic Specialization:
The aforementioned spontaneous release of neurotransmitters prior to the
establishment of a functional synapse is one mechanism that can trigger the induction
of the postsynaptic assembly and stabilizes the site of contact (Lohmann, Finski et al.
2005). It was shown that the postsynaptic neuron starts to express receptors, capable
of transmitter detection preceding synapse formation (Haydon, Cohan et al. 1985;
Spencer, Lukowiak et al. 2000). These events of transmitter release and detection
between two neurons are in addition assumed to be crucial for attracting the respective
synaptic partner and repelling the wrong one (Spencer, Lukowiak et al. 2000) as well as
for timing synapse formation (Lovell, McMahon et al. 2002). After matching the correct
synaptic partner, the postsynaptic assembly starts in a sequential manner. Scaffolding
proteins for the formation of the postsynaptic density (PSD) were shown to be recruited
within a couple of minutes followed by the recruitment of glutamate receptors and PSD
signaling molecules (Bresler, Ramati et al. 2001). Comparable to the trafficking of
presynaptic proteins, it was shown that NMDA receptors do also travel in distinct
vesicles (Washbourne, Bennett et al. 2002; Washbourne, Liu et al. 2004). These
vesicles seem to carry further devices of the postsynapse, such as AMPA receptors and
scaffolding proteins (Washbourne, Bennett et al. 2002). Interestingly, owing to these
NMDA packages, the immature neuron is already able to detect glutamate during the
transport process (Washbourne, Liu et al. 2004; McAllister 2007), accounting for the
induction of postsynaptic assembly following presynaptic transmitter release.
A couple of complex molecules are known to be involved in forming the postsynaptic
compartment. One of the most prominent proteins is the neural activity related protein
(Narp). Narp is involved in the insertion of AMPA receptors in the postsynaptic
membrane (O'Brien, Xu et al. 1999). Further, ephrinB was shown to interact directly
with the NR1 subunit of NMDA receptors and to promote clustering of NMDA subunits
(Dalva, Takasu et al. 2000). Ephrins, in general, where shown to be crucially involved in
- 11 -
Chapter 1
General introduction
spine maturation (Penzes, Beeser et al. 2003). Neurexin induces postsynaptic
differentiation and local clustering of NMDA receptors and PSD-95 in concert with the
binding partner neuroligin (Graf, Zhang et al. 2004).
Thus, the postsynapse assembles in a sequential and well organized manner.
1.1.2 Synapse Maturation and Pruning
After the initial formation of a synaptic connection, which can occur within hours, the
synapse undergoes a prolonged period of synaptic maturation (Ahmari, Buchanan et al.
2000). This time is characterized by an enlargement of the side of contact, an
enhancement of pre- and postsynaptic proteins stabilizing the sides of synaptic contact,
an increase of transmitter content and a late and very characteristic step: the formation
of dendritic spines (Yuste and Bonhoeffer 2004; McAllister 2007).
Initial synaptic contacts are formed on dendritic shafts or filopodia, which become
further specialized during neuronal development and build mature dendritic synaptic
spines. Spines come in different flavors (thin, stubby, branched and mushroom) and are
indispensable for the function and the plastic properties of the CNS (Calabrese, Wilson
et al. 2006). Accordingly, dendritic spines are thought to increase the neuronal surface
and to compartmentalize the dendritic shaft, thereby modifying the synaptic input and
allowing plastic adaption to occur (for review see Yuste 2011). Dendritic spine
formation requires extensive remodeling of the actin cytoskeleton, induced by different
molecules such as cadherins, ephrins, extracellular matrix molecules and syndecan
activating a variety of Rho- and Ras family GTPases (Hering and Sheng 2001). Once, a
synaptic connection has to be maintained, spine formation was shown to become
immediately induced and NMDA receptor activation was shown to be crucially involved
in shaping synaptic spine morphology regulating the actin turnover (Okabe, Urushido et
al. 2001; Fukazawa, Saitoh et al. 2003). The postponed AMPA receptor activation is
involved in maintaining the dendritic spine, reducing spine motility and increasing spine
size and density (McKinney, Capogna et al. 1999; Fischer, Kaech et al. 2000).
- 12 -
Chapter 1
General introduction
The described induction and maturation of spines already points to a crucial and
probably most important factor for shaping the CNS: neuronal activity. This was already
postulated by Donald Hebb by his claim
“Cells that fire together, wire together” (Hebb 1949).
Consequently, synapses that receive converging synaptic input become strengthened,
while less used synapses are removed. In addition to the correct, long lasting synaptic
contacts, it is known that neurons form numerous contacts, which are eliminated
afterwards. The developing brain contains far more synapses than the fully matured.
For this reason, synaptic pruning is also implicated in the formation of neuronal circuits
(Lichtman and Colman 2000; Hashimoto and Kano 2003). Neuronal activity and the
proper release of neurotransmitters are fundamental steps therein. This was
underlined by studies, in- or decreasing the sensory input to the visual or
somatosensory system, directly resulting in respective changes in the synapse number
(LeVay, Wiesel et al. 1980; Knott, Quairiaux et al. 2002; Trachtenberg, Chen et al. 2002).
Accordingly, deprivation of sensory input leads to a severe reduction in synapses, while
increasing external stimulation leads to the extensive formation of new ones.
But what are the cellular and molecular substrates of the observed phenomena of
rapidly forming and degrading synaptic contacts?
Neurotrophins are released upon intense neuronal activity and it was shown that they
are crucially involved in the maturation and differentiation of neurons (for review see
Vicario-Abejon, Owens et al. 2002). Neurotrophins occur locally at sites of synaptic
contact, thereby initiating or stabilizing synaptic contacts and forcing their maturation
(Goodman, Valverde et al. 1996; Haubensak, Narz et al. 1998). Prominent neurotrophins
are the BDNF, nerve growth factor (NGF) as well as the neurotrophins NT-3, NT-4 and
NT-5. Neurotrophins bind to two transmembrane receptors namely the Trk
(tropomyosin receptor kinase) and the p75 receptors, which can induce intracellular
signaling cascades enhancing the formation of synapses (Vicario-Abejon, Owens et al.
2002). Further, the insertion of AMPA receptors is assumed to occur in a second wave
- 13 -
Chapter 1
General introduction
and enables former silent synapses for effective synaptic transmission as well as
stabilizing existing synaptic connection (Song and Huganir 2002; Malenka 2003).
In parallel, synapses can become eliminated and synaptic proteins become degraded
very fast in response to diminished neuronal input. This activity-dependent fast
turnover of synaptic proteins was shown to occur mostly via ubiquitination of pre- and
postsynaptic proteins (Ehlers 2003). Along these lines, Ehlers et al. 2003 demonstrated
that the postsynaptic proteins ProSAP/Shank, GKAP and AKAP79/150 (A-kinase
anchoring protein) become ubiquitinated upon decreased synaptic activity. Further,
there exist hints in the literature that the presynaptic machinery is also subject to
ubiquitination (Speese, Trotta et al. 2003).
1.1.4 Synaptic plasticity
In the field of neuroscience, the term “Plasticity” roughly describes the ability of the
CNS to adapt to the changing requirements a species is subjected to. This can be found
on the level of huge projections and representations, such as after brain damage
(Geissler, Dinse et al. 2011) or in the hippocampus during learning (Abraham 2008). On
the cellular level the term “synaptic plasticity” describes the context-dependent
strengthening, weakening, forming, elimination, or refinement of synapses.
The crucial factor here, as well as during developmental synaptic maturation and
synapse pruning (see chapter 1.1.3), is the intensity with which the site of contact is
used. Thus, neuronal activity tunes and shapes the intensity of synaptic contacts. On
the other hand, the neuronal circuits can respond via short- or long term changes in
synaptic efficiency. The most prominent types of synaptic plasticity are long term
potentiation (LTP) and long term depression (LTD). During LTP, a brief coordinated
increase in the neuronal input leads to a long-lasting increase in synaptic strength,
while LTD mirrors the long-lasting decrease in synaptic efficiency (Bliss and
Collingridge 1993). These phenomena where intensively studied in the hippocampus,
but can probably be found in the vast majority of CNS neurons (for review see
Collingridge, Isaac et al. 2004). Further, presynaptic mechanisms, via which neuronal
circuits can adjust synaptic signaling, are mediated via changes in the release
- 14 -
Chapter 1
General introduction
probability of vesicular neurotransmitters, changes in the quantal size of these vesicles,
modifications of the “ready releasable pool” of vesicles and adjustment of the docking
areas for synaptic vesicles. After the release of neurotransmitter, the chemical signal
transduction can be subject to further alteration, and in recent years evidence for the
involvement of astrocytes and extracellular matrix (ECM) molecules in synaptic
transmission emerged. On the next level, postsynaptic modifications as receptor
expression, receptor turnover and general receptor composition of the PSD as well as a
myriad of intracellular specifications allows additional modifications in the synaptic
efficacy.
1.2 Astrocytes
Neurons are no single-players, instead they tightly interact with glia cells, the second
cell class in the brain. The family of glia cells comprises different cell types, namely
oligodendrocytes, microglia, and astrocytes. Oligodendrocytes are the myelin forming
cells in the CNS, enwrapping central axons. Microglia represent the immune system of
the brain, scavenging the brain from cell fragments, molecular debris and infectious
material. Nevertheless, Astrocytes are probably the most multitalented cells in the
family of glia cells. The spectrum of responsibilities reaches from the blood-brainbarrier formation over the uptake and the recycling of glutamate up to the release of
neurotrophins and the active participation in neuronal transmission. That is why
astrocytes are today accepted as indispensable neuronal partners, actively
participating and dynamically influencing the synaptic language. Beyond that, a bunch
of publications have accumulated showing that astrocytes themselves are excitable
cells, exchanging currents via calcium waves and the release of glio-transmitters.
Oligodendrocytes and Microglia, as well as the recently discussed fourth family
member, the NG2 cells (Nishiyama, Komitova et al. 2009) are not addressed in the
present work and the focus will be placed on astrocytes.
- 15 -
Chapter 1
General introduction
1.2.1 Astrocytes - a changing image
The picture of astrocytes dramatically changed during the past twenty five years and is
still evolving (for review see Volterra and Meldolesi 2005; Faissner, Pyka et al. 2010).
Initially speculated as being a kind of glue, sticking the protagonistic neurons together,
astrocytes are today known to be crucial for promoting neuronal survival and signal
transmission. This functional variety is further increased by a huge diverseness in the
astrocytic cell population. Astrocytes can be distinguished among one another by the
expression of unique sets of genes and by triggering different currents, varying between
different brain regions and confirming the heterogeneity of this cell population (Bachoo,
Kim et al. 2004; Grass, Pawlowski et al. 2004; Wallraff, Odermatt et al. 2004). Beside
the intimate relationship with neurons (see chapter 1.2.3), astrocytes are involved in a
lot of other processes in the brain. Thus, they build bridges between the vasculature
and neurons, they can guide other non-neuronal cells to their prescribed destination,
and they play an active role in protecting intact tissue after brain damage (Tsai, Frost et
al. 2002; Babcock, Kuziel et al. 2003).
Nevertheless, the interaction with neurons is probably the most enigmatic one and will
be introduced further.
1.2.3 Neuron-glia interaction - the tripartite synapse
It is generally accepted, that astrocytes are indispensable for neuronal survival,
supporting the development of neurons by the release of different glial factors (for
review see Faissner, Pyka et al. 2011). In line with this, primary hippocampal neurons,
growing in defined medium, die within a few days without added astroglia and they can
not be replaced by other supporting cells, such as fibroblasts (Pyka, Busse et al. 2011).
Astrocytes release a couple of neurotrophins and other supporting molecules the
neurons can benefit from (Cahoy, Emery et al. 2008). Beside the survival supporting
properties, the astrocyte-derived molecules seem to foster especially the structural
formation and the functional activation of synapses. Some of the responsible factors
were identified in the past years and many impressive studies demonstrated, that cocultured astrocytes or astrocyte conditioned medium (ACM) are indispensable for
- 16 -
Chapter 1
General introduction
proper synapse formation (Beattie, Stellwagen et al. 2002; Slezak and Pfrieger 2003;
Pyka, Wetzel et al. 2011). As astrocyte-released synaptogenic factors, thrombospondin
(TSP) (Christopherson, Ullian et al. 2005), cholesterol and apolipoprotein E (Mauch,
Nagler et al. 2001) were identified. Nevertheless, retinal ganglion cells, growing in TSP
enriched medium, remain silent and AMPA receptor responsiveness is missing
(Christopherson, Ullian et al. 2005), suggesting that a couple of further synaptogenic
factors has to be tagged in the future.
The spatial proximity of neurons and astrocytes, especially at the site of synaptic
contact led to the concept of the tripartite synapse (Araque, Parpura et al. 1999) (see
Fig. 5). Astrocytes are organized in territories in vivo and one cell can contact and
tightly enwrap thousands of synapses (Bushong, Martone et al. 2002). Astrocytes can
exert their influence especially due to the close synaptic proximity, e.g. via the
expression of defined protocadherins (Garrett and Weiner 2009) or ephrins (Carmona,
Murai et al. 2009). The intimate relationship between astrocytic processes and the
neuronal synapse is not static rather it is plastic, thus modulating synaptic
transmission (Hirrlinger, Hulsmann et al. 2004). This underlines that the model of the
tripartite synapse is also crucial for many aspects of synaptic plasticity.
The release of extracellular matrix (ECM) molecules from astrocytes seems to play a
further crucial role for neuronal development and synapse formation. The probably
most prominent and intensively studied example is the ECM at the neuromuscular
junction (NMJ) (Sanes and Lichtman 1999; Singhal and Martin 2011). At this well
suited synapse model, the distinct distribution of ECM molecules dictates the clustering
of acetylcholine and synapses. It is known that ECM molecules such as agrin and
perlecan are significantly involved in nearly all aspects of synaptogenesis, synapse
stability, and synaptic transmission at this peripheral synapse (Singhal and Martin
2011). There accumulated evidence that the ECM does also play a fundamental role at
the central synapse (Frischknecht, Heine et al. 2009; Pyka, Wetzel et al. 2011). In line
with that conclusion, approaches extending the model of the tripartite synapse to a
tetrapartite model emerged. They include the presynapse, the postsynapse, the
astrocytic process and the synaptic extracellular matrix (Dityatev and Rusakov 2011).
- 17 -
Chapter 1
General introduction
Figure 5: Model of the tripartite synapse
The neuronal pre- and postsynaptic side of contact is closely enwrapped by astrocytic processes.
- 18 -
Chapter 1
General introduction
1.3 The extracellular matrix
In general, the extracellular matrix (ECM) is a connective macromolecular assembly,
giving rise to the shape of a tissue and organizing the cells within it. ECM molecules are
found in tissues across the organisms. With regard to the huge diversity of tissues,
reaching from solid bones and teeth to the resilient tendons and the transparent
cornea, the ECM has to come in quite different flavors.
The ECM is a meshwork of macromolecules produced by the cells themselves, and is
therefore tightly associated with their surfaces and does exactly fit to the requirements
in the respective developmental context (Dityatev, Seidenbecher et al. 2010; Faissner,
Pyka et al. 2010). The ECM presents an important substrate for cell-cell
communication and is well suited for presenting signaling molecules for guiding cells.
Thus, the composition of matrix molecules is extremely variable in a time- and spacedependent manner.
The ECM in general is mainly made of fibrous proteins such as collagen and elastin,
proteoglycans attached to glycosaminoglycans (GAGs), and glycoproteins like
fibronectin and tenascins (Faissner 1993; Bandtlow and Zimmermann 2000). The
defined composition and the relative amounts of these molecules within the matrix give
rise to the texture of a tissue and fit exactly to the defined requirements. Thus, the
unique ECM of the CNS has to fulfill special demands.
1.3.1 The composition of the brain´s ECM
The exact composition of the ECM in the nervous system highly depends on time and
space and therefore changes dramatically during development. In contrast to the
collagen-rich peripheral connective tissues, the ECM of the CNS is mainly composed of
glucosaminoglycans, proteoglycans and glycoproteins, while fibrous proteins such as
collagen and elastin are nearly absent (Asher, Perides et al. 1991).
- 19 -
Chapter 1
General introduction
1.3.1.1 Glycosaminoglycans (GAGs)
GAGs are polysaccharides, occurring frequently in the ECM of the central nervous
system. They consist of long, unbranched repeating (≈ 20 – 200) disaccharide units (for
review see Bandtlow and Zimmermann 2000). In general, GAGs are classified with
respect to their disaccharide composition. Accordingly, one can distinguish
Chondroitinsulfate (CS), Heparansulfate (HS), Keratansulfate (KS), Dermatansulfate
(DS) (see Fig. 6) and Hyaluronan (HA). The disaccharide units can be subject to a
diversity of modifications, such as carboxylation or sulfation (Bulow and Hobert 2006).
The unique composition of disaccharides and the remarkable amount of posttranslational modifications makes GAGs to one of the most information-dense
biological molecules (Turnbull, Powell et al. 2001).
Figure 6: The different disaccharide units found in glycosaminoglycans (GAGs). Molecular structure
of the disaccharide units that form the GAG chains: Chondroitinsulfate (CS), Heparansulfate (HS),
Keratansulfate (KS), Dermatansulfate (DS). Modifications are colored (from Bulow and Hobert 2006).
Due to the molecular structure, GAGs are suitable for binding a diversity of signaling
molecules and they play a multifarious role in the brains ECM.
Hyaluronan is an exception within this list of GAGs, because it is not bound to a protein,
it is non-sulfated and it is made of identical disaccharide units (Toole 2001). Hyaluronan
is synthesized within the plasma membrane, forms the backbone of the extracellular
- 20 -
Chapter 1
General introduction
matrix and is a crucial component in perineuronal nets (PNNs) (see chapter 1.3.2). In
contrast to hyaluronan, most of the aforementioned GAGs are covalently attached to a
core protein, a proteoglycan (see chapter 1.3.1.2).
1.3.1.2 Proteoglycans
GAGs bind to proteoglycans through a serine residue and a specific carbohydrate
tetrasaccharide linker region. Proteoglycans can be divided into different subclasses
with respect to the GAGs they bind to. Thus, one distinguishes Chrondroitin sulfate
proteoglycans (CSPGs), Heparan sulfate proteoglycans (HSPGs), Keratan sulfate
proteoglycans (KSPGs) and Dermatan sulfate proteoglycans (DSPGs).
CSPGs are the most abundant proteoglycans in the CNS and were shown to be
crucially involved in a diversity of developmental processes, regeneration and synaptic
plasticity (Carulli, Laabs et al. 2005; Pyka, Wetzel et al. 2011). The most prominent
family within the CSPGs are the lecticans, which comprise neurocan, aggrecan,
brevican, and versican (Yamaguchi 2000) (see Fig. 7). While versican was identified for
the first time to be produced by fibroblasts (Zimmermann and Ruoslahti 1989), and
aggrecan was already known to be abundantly expressed in cartilage (Doege, Sasaki et
al. 1987), neurocan and brevican represent ECM molecules exclusively expressed in the
CNS (Rauch, Karthikeyan et al. 1992; Yamada, Watanabe et al. 1994). All of the four
lecticans display unique expression patterns during development (Milev, Maurel et al.
1998). In agreement with this notion, the appearance of brevican and aggrecan
increases from embryonic day (E) 14 until postnatal day (P) 100, while neurocan shows
the opposite expression patterns, with highest rate from embryonic stages until P2-P6,
where after it progressively decreases with increasing age. Versican shows a unique
isoform specific expression pattern (Milev, Maurel et al. 1998). This distinct timedependent transcriptional control already hints at the unique role each molecule exerts
during development. Lecticans are known to be involved in migration, axon guidance,
cell adhesion, synapse formation and synaptic plasticity (Faissner, Clement et al. 1994;
Dityatev, Schachner et al. 2010) and one can assume that the whole functional
spectrum of the individual lecticans has not been completely unraveled so far.
Manipulations of the aggrecan and versican gene lead to lethal mouse mutants
- 21 -
Chapter 1
General introduction
(Watanabe, Kimata et al. 1994; Mjaatvedt, Yamamura et al. 1998), due to heart deficits
and respiratory failure, while mice with mutations in the brevican or neurocan gene
suffer only mild deficits (Zhou, Brakebusch et al. 2001; Brakebusch, Seidenbecher et al.
2002; Bekku, Rauch et al. 2009). However, these mild deficits can help to unravel the
function of these proteins. Along these lines, the genetical depletion of neurocan was
shown to lead to changes in the late phases of LTP (Brakebusch, Seidenbecher et al.
2002), underlining the functional importance of these molecules in synaptic plasticity.
Figure 7: The Lectican family of the CSPGs
Molecular composition of the four lecticans: aggrecan, versican, neurocan, and brevican with the
respective splicevariants (Bandtlow and Zimmermann 2000).
CSPGs are abundantly expressed in growth barriers and guide axons to their
appropriate targets. Most of the CSPGs are highly negatively charged and therefore
exert in most cases repellent and inhibitory properties in their environment, such as
inhibiting neurite outgrowth (Friedlander, Milev et al. 1994) or synapse formation (Pyka,
Wetzel et al. 2011). The expression of CSPGs shows a remarkable
increased
expression after central and peripheral lesion (Kwok, Dick et al. 2011), which is most
- 22 -
Chapter 1
General introduction
prominent in the glial scar (Shen, Li et al. 2008). The formation of this inhibitory growth
barrier protects the adjacent tissue against further damage, but the side effects are
diminished
regenerative
capacities
(Carulli,
Laabs
et
al.
2005).
Therefore,
pharmacological agents such Hyaluronidase or ChrondroitinaseABC (ChABC) have
been used in order to restore the regenerative capacities (Bradbury, Moon et al. 2002;
Garcia-Alias, Barkhuysen et al. 2009; Wang, Ichiyama et al. 2011).
Nevertheless, CSPGs with neurite outgrowth promoting properties were also described
in the past (Faissner, Clement et al. 1994).
The lecticans show a characteristic molecular structure: They consist of a central
domain, which carries the respective sugar chains, the N-terminal globular domain,
which can bind to hyaluronan and the C-terminal globular domain containing a C-type
lectin domain flanked by EGF- and complement regulatory protein (CRP)-like domains
(Iozzo 1998; Yamaguchi 2000) (see Fig. 7). This C-type domain can bind e.g. to
glycoproteins such as tenascin-R and is important for interaction between different
matrix molecules (Aspberg, Binkert et al. 1995), forming together with hyaluronan a
macromolecular meshwork.
1.3.1.2 Glycoprotein
Glycoproteins are oligosaccharide side chain bearing proteins. Common examples of
glycoproteins are laminin, fibronectin and the tenascins. Fibronectin, a ~ 440 kDa
protein, is a prominent glycoprotein indispensable for proper development of different
organs, such as heart and vasculature. Thus, the fibronectin knock-out mouse is lethal
during embryogenesis (Watt and Hodivala 1994). Fibronectin is subject to alternative
splicing and occurs in different isoforms. A repeating motif within the huge molecule
are the fibronectin type III repeats (FNIII), which are also found in a variety of ECM
molecules as well as in tenascin-C (Van Obberghen-Schilling, Tucker et al. 2011).
Tenascin-C is composed of 14.5 EGF repeats and a maximum of 17 (in humans, 14 in
mice) FN Type III domains (see Fig. 8). A fraction of FN Type III domains is alternatively
spliced, leading to a huge amount of different isoforms.
Theoretically, this alternative splicing can lead to 26 different tenascin-C molecules in
the mouse and 29 in humans. In the mouse CNS, 27 different isoforms are described so
- 23 -
Chapter 1
General introduction
far (Joester and Faissner 2001; von Holst, Egbers et al. 2007). Six tenascin-C
molecules are assembled to a hexabrachion at the cystein-rich N-terminus. Tenascin-C
is a crucial component for central nervous system development and was in the recent
years also shown to be important for synaptic plasticity. For example, the knock-out of
tenascin-C leads to reduced LTP in the hippocampal CA1 region (Evers, Salmen et al.
2002), functional and structural abnormalities in cortical development (Irintchev,
Rollenhagen et al. 2005) and changes in the migratory behavior of oligodendrocyte
precursor cells (OPCs) along the optic nerve (Garcion, Faissner et al. 2001). In general,
tenascin-C is a protein crucially involved in CNS development (Garcion, Halilagic et al.
2004; Czopka, Von Holst et al. 2009; Czopka, von Holst et al. 2010; Karus, Denecke et
al. 2011).
Figure 8: Modular structure of tenascin-C
The molecular composition of tenascin-C is shown. The red FN Type III domains are alternatively spliced
and give rise to a variety of tenascin-C isoforms. The binding partners with respective binding sides are
indicated (taken from Van Obberghen-Schilling, Tucker et al. 2011).
Tenascin-R, the second family member of the tenascin glycoproteins is also abundantly
expressed in the central nervous system. While tenascin-C is mainly expressed in early
phases of development and decreases with age, tenascin-R is a crucial component of
the mature ECM and is part of PNNs, which appear around a subpopulation of neurons
at the end of the critical period (see chapter 1.3.4). In addition, tenascin-R is enriched at
- 24 -
Chapter 1
General introduction
the nodes of Ranvier and is essential for proper axonal conduction velocities (Weber,
Bartsch et al. 1999).
Tenascin-R contains also EGF-repeats and the FN type III domains (see Fig. 9), which
are subject to alternative splicing, even if the number of alternatively isoforms is
smaller as for tenascin-C and limited to two variants (160 and 180 kDa) so far.
Figure 9: modular domain structure of tenascin-R
Tenascin-R 4.5 EGF repeats and 9 FN type III repeats. The R1 domain is alternatively spliced (taken from
Dityatev and Schachner 2003).
Tenascin-R and tenascin-C are carrier of the human natural killer-1 carbohydrate
(HNK-1) (Saghatelyan, Gorissen et al. 2000). Modifications within the tenascin-R gene,
as well as in the synthesis of the HNK-1 motif where shown to result in changes in
hippocampal CA1 LTP (Saghatelyan, Gorissen et al. 2000; Yamamoto, Oka et al. 2002),
suggesting again the crucial involvement of ECM molecules in synaptic plasticity.
- 25 -
Chapter 1
General introduction
1.3.2 The “tetrapartite Synapse”
In the previous chapters the rough composition of the central nervous system ECM was
depicted and the involvement of some of these molecules in synaptic formation,
maturation and plasticity was already pointed out. In recent years, the picture of the
“tripartite synapse” was expanded to a “tetrapartite synapse” including beyond the
astrocyte also the ECM to the classical bipolar view of the chemical synapse. Today we
know that the ECM and the modification of the ECM can strongly affect the synaptic
transmission machinery. A recently published review by Dityatev and Rusakov 2011,
(see Fig. 10) summarized the today´s knowledge about the interactions between the
four partners interacting at the site of neuronal contact.
Figure 10: Model of the tetrapartite synapse:
The intense interaction between the four different partners (presynapse, postsynapse, glia and ECM) are
depicted (taken from Dityatev and Rusakov 2011).
- 26 -
Chapter 1
General introduction
1.3.3. The quadruple knock-out mouse
In order to unravel the complete functional spectrum of extracellular matrix molecules
during neuronal network development and maintenance, a couple of mice, carrying
mutations in the respective genes were generated. As mentioned above, some of these
mutants are lethal due to severe developmental failures of the heart and the
respiratory system (Watanabe, Kimata et al. 1994; Mjaatvedt, Yamamura et al. 1998) ,
while others show no gross alterations (Evers, Salmen et al. 2002; Cybulska-Klosowicz,
Zakrzewska et al. 2004; Irintchev, Rollenhagen et al. 2005; Sykova, Vorisek et al. 2005;
Bekku, Rauch et al. 2009; Faissner, Pyka et al. 2010).
Despite their mild phenotypes, these mouse mutants are considered to be a versatile
tool in understanding the involvement of ECM molecules in brain formation and
function. Some of the results suggest that the ECM is very flexible and that the lack of
ECM components can be compensated by other members. For instance, the quadruple
knock-out mouse (Rauch, Zhou et al. 2005), deficient for tenascin-C, tenascin-R,
neurocan, and brevican was shown to up regulate fibulin-1 and fibulin-2, which are
normally not predominantly expressed in the CNS.
Figure 11: Structural organization of the ECM in the quadruple knock-out mouse
Left: The molecular interaction of the four matrix molecules: tenascin-C, tenascin-R, brevican and
neurocan with hyaluronan are depicted. Right: The alternative matrix made of fibulin-1 and fibulin-2
interacting with versican and aggrecan (taken from Rauch, Zhou et al. 2005).
- 27 -
Chapter 1
General introduction
Interestingly, both fibulins have been shown to interact with aggrecan and versican
(Aspberg, Adam et al. 1999; Olin, Morgelin et al. 2001). Nevertheless, the only existing
study about the quadruple mutant by Rauch et al. 2009 revealed no gross
morphological changes, but a slight difference in the hyaluronan immunoreactivity and
a change in the density of perineuronal nets.
We argue that despite the maintenance of the broad structural organization of the ECM,
the quadruple knock-out mice can be valued a versatile tool to gain insight into the role
of the matrix during development.
1.3.4 Perineuronal nets
Perineuronal nets (PNNs) represent a specialized form of the brain ECM. PNNs are
lattice-like aggregates of ECM molecules that accumulate around a subpopulation of
neurons, tightly enwrapping the soma and dendrites. Different ECM molecules such as
CSPGs and hyaluronan have been shown to be part of PNNs (Asher, Perides et al. 1991;
Bruckner, Hartig et al. 1996), but the detailed composition is highly variable in a timeand region dependent manner (Lander, Kind et al. 1997). First described by Golgi and
colleagues, PNNs are in the main focus of today´s research (Pizzorusso, Medini et al.
2002; Balmer, Carels et al. 2009; Gogolla, Caroni et al. 2009), but they look back to a
long history (see Fig. 12). One reason for the remarkable interest in PNNs emerging in
the recent years may be rooted in the observation, that the formation of PNNs is a
peculiarity of the matured brain, while it is rarely found in the infantile, still plastic
brain. Further, there is growing evidence that the formation of this netlike ECM is a
prerequisite for the maintenance and stabilization of synaptic connections, thus storing
information in the adult brain (Balmer, Carels et al. 2009; Gogolla, Caroni et al. 2009).
1.3.4.1 PNN composition and formation
Already in the first half of the last century it was noticed, that the appearing net-like
structures, accumulating around some neurons, are composed of hyaluronic acid,
glycosaminoglycans and chondroitin sulfates (Glegg and Pearce 1956; Feigin 1980).
Nevertheless, the crucial molecular components for proper net formation, the PNN
- 28 -
Chapter 1
General introduction
organization and the detailed steps necessary for PNN recruitment are still subject of
debate in the current literature (Giamanco, Morawski et al. 2010; Kwok, Carulli et al.
2010; Bekku, Saito et al. 2011).
Figure12: First drawings of PNNs by Camillo Golgi and colleagues
Perineuronal nets accumulating around subpopulations of neurons were already recognized by the
th
pioneers in neuroscience during the 19 century.
A: Nerve cell from the anterior horn of cat spinal cord with enlarged details in (a) and (b) B: Two cerebral
cells from the adult cat C: Nerve cell derived from the anterior horns of the dog spinal cord D: Cell with
Golgi’s net and a diffuse net (anterior horn of the spinal cord of a calf embryo). E: cortical cell of an adult
dog F: Alterations within the perineuronal net of a human cortical cell, derived from a patient with
paralytic dementia. G: Cell derived from the nucleus of the vagus (medulla oblongata) of Lacerta muralis.
Taken from, and original sources given in Celio, Spreafico et al. 1998.
The today´s knowledge about the composition of perineuronal nets mainly lists
hyaluronan, CSPGs (aggrecan, neurocan, versican, brevican and phosphacan), tenascinR, and the link proteins brain-specific link protein 2 (Bral2) and cartilage link protein 1
(Crtl1). There exits evidence, that hyaluronan, aggrecan and Crtl1 are essential and
sufficient to induce proper net formation (Kwok, Carulli et al. 2010) and recent data
suggest that the expression of neuron-derived link proteins is a crucial factor for the
- 29 -
Chapter 1
General introduction
initiation of the PNN construction (Bekku, Su et al. 2003; Carulli, Pizzorusso et al. 2010;
Bekku, Saito et al. 2011). Thus, mice lacking either Crtl-1 or Bral-2 show attenuated
PNNs (Carulli, Pizzorusso et al. 2010; Bekku, Saito et al. 2011). Bral-2 was shown to be
especially involved in the recruitment of brevican (Bekku, Saito et al. 2011), while Crtl-1
seems to play a fundamental role in initiating PNN formation (Carulli, Pizzorusso et al.
2010).
Nevertheless, there is a huge heterogeneity of the detailed PNN composition, depending
on the CNS subregion. Thus, it was shown, that the expression of the different aggrecan
isoforms is highly variable between different PNNs in the cerebral cortex (Matthews,
Kelly et al. 2002; Virgintino, Perissinotto et al. 2009), while the functional consequences
of different PNN compositions are still not clear.
Neurons and glia cells contribute to the synthesis of PNN molecules (Carulli, Rhodes et
al. 2006), but neurons themselves seem to be the main source and therefore the
coordinators of the special PNN composition (Matthews, Kelly et al. 2002).
It was noticed, that the phenomenon of bearing a PNN seems to be a characteristic
feature of only certain subpopulations of neurons. PNNs where described to locate
especially around parvalbumin-positive interneurons (Dityatev, Bruckner et al. 2007;
Balmer, Carels et al. 2009), but it was found, that PNNs can additionally accumulate
around excitatory neurons. In line with this, PNNs were identified around cortical
pyramidal cells (Wegner, Hartig et al. 2003), especially in the visual cortex (Alpar,
Gartner et al. 2006). Different publications hint to a similarity between PNN wearing
neurons in the expression of the potassium channel Kv3.1b, in cortex as well as in
spinal cord (Hartig, Singer et al. 2001; Wegner, Hartig et al. 2003; Vitellaro-Zuccarello,
Bosisio et al. 2007). However, the subtype determining mechanisms and the overall
similarity that PNN bearing neurons share have still to be unraveled, in order to
understand the functional relevance of these ECM accumulations.
1.3.4.2 PNN function
The structure, at a first glance thought to be an artifact derived from the coagulation of
the pericellular fluid during the staining procedure (for review see Celio, Spreafico et al.
1998), is today known to be crucially involved in the formation and the plastic
- 30 -
Chapter 1
General introduction
properties of the maturing brain. The formation of PNN coincides with the end of the
critical period (Pizzorusso, Medini et al. 2002; Balmer, Carels et al. 2009; Gogolla,
Caroni et al. 2009). This observation and the possibility to delay the closure of this
window of enhanced plasticity via enzymatic removal of PNNs, led to the idea that
PNNs are crucially involved in restricting plasticity in the matured brain. In line with
this, it could be shown that the injection of ChABC can restore ocular dominance
plasticity in the adult cat visual cortex (Pizzorusso, Medini et al. 2002) and that Crtl-1
deficient mice with attenuated PNNs retain juvenile levels of ocular dominance
plasticity (Carulli, Pizzorusso et al. 2010). Beside these observations related to the
visual system it could be shown, that storing fear memory and song learning in birds is
also related to PNN formation (Balmer, Carels et al. 2009; Gogolla, Caroni et al. 2009).
Different studies described the formation of PNNs in an activity-dependent manner
(Lander, Kind et al. 1997; Dityatev, Bruckner et al. 2007). Thus, the blocking of actionpotentials, transmitter-release, Ca2+-permeable AMPA receptors or L-type voltagegated Ca2+-channels in vitro led to a significant reduction in PNN wearing cell (Dityatev,
Bruckner et al. 2007). Nevertheless, the knowledge about the functional correlate is
still fragmentary. Recently, it was shown, that the axon guidance molecule semaphorin
3A (Sema3A) (Schwarting, Kostek et al. 2000) is localized in PNNs (De Wit, De Winter et
al. 2005 and unpublished data from personal correspondence with James Fawcett,
Cambridge). Sema3A is known to be crucially involved in axon guidance, therefore
supporting the idea, that PNNs control the formation of synaptic contacts. In line with
that, the expression of Sema3A might be one of the neuronal mechanisms to repel
axons and to restrict plasticity.
In addition, a few additional functions of PNNs are discussed in the literature. PNNs can
accumulate growth factors around certain neurons, act as a buffering system for ions,
protect against oxidative stress and function as a microenvironment for highly active
neurons (Blumcke, Weruaga et al. 1994; Hartig, Derouiche et al. 1999). In addition,
PNNs were shown to restrict lateral mobility of AMPA receptors, referring to the
involvement of PNNs in synaptic plasticity (Frischknecht, Heine et al. 2009).
- 31 -
Chapter 1
General introduction
In summary, the research of the recent years highly contributed to the existing
knowledge about the role of the ECM. As this introduction pointed out, there exist a
couple of complex interactions in the ECM related neuron-glia relationship. The
emerging role of the ECM in synapse development as well as in synaptic plasticity
raises a lot of further questions, which were partially addressed in the present work.
- 32 -
Chapter 1
References
1.5. References
Abraham, W. C. (2008). "Metaplasticity: tuning synapses and networks for plasticity." Nat Rev
Neurosci 9(5): 387.
Ahmari, S. E., J. Buchanan, et al. (2000). "Assembly of presynaptic active zones from
cytoplasmic transport packets." Nat Neurosci 3(5): 445-451.
Akaneya, Y., K. Sohya, et al. (2010). "Ephrin-A5 and EphA5 interaction induces synaptogenesis
during early hippocampal development." PLoS One 5(8): e12486.
Alpar, A., U. Gartner, et al. (2006). "Distribution of pyramidal cells associated with perineuronal
nets in the neocortex of rat." Brain Res 1120(1): 13-22.
Alsina, B., T. Vu, et al. (2001). "Visualizing synapse formation in arborizing optic axons in vivo:
dynamics and modulation by BDNF." Nat Neurosci 4(11): 1093-1101.
Araque, A., V. Parpura, et al. (1999). "Tripartite synapses: glia, the unacknowledged partner."
Trends Neurosci 22(5): 208-215.
Arikkath, J. and L. F. Reichardt (2008). "Cadherins and catenins at synapses: roles in
synaptogenesis and synaptic plasticity." Trends Neurosci 31(9): 487-494.
Asher, R., G. Perides, et al. (1991). "Extracellular matrix of central nervous system white
matter: demonstration of an hyaluronate-protein complex." J Neurosci Res 28(3): 410421.
Aspberg, A., S. Adam, et al. (1999). "Fibulin-1 is a ligand for the C-type lectin domains of
aggrecan and versican." J Biol Chem 274(29): 20444-20449.
Aspberg, A., C. Binkert, et al. (1995). "The versican C-type lectin domain recognizes the
adhesion protein tenascin-R." Proc Natl Acad Sci U S A 92(23): 10590-10594.
Babcock, A. A., W. A. Kuziel, et al. (2003). "Chemokine expression by glial cells directs
leukocytes to sites of axonal injury in the CNS." J Neurosci 23(21): 7922-7930.
Bachoo, R. M., R. S. Kim, et al. (2004). "Molecular diversity of astrocytes with implications for
neurological disorders." Proc Natl Acad Sci U S A 101(22): 8384-8389.
Balmer, T. S., V. M. Carels, et al. (2009). "Modulation of perineuronal nets and parvalbumin with
developmental song learning." J Neurosci 29(41): 12878-12885.
Bandtlow, C. E. and D. R. Zimmermann (2000). "Proteoglycans in the developing brain: new
conceptual insights for old proteins." Physiol Rev 80(4): 1267-1290.
Basarsky, T. A., V. Parpura, et al. (1994). "Hippocampal synaptogenesis in cell culture:
developmental time course of synapse formation, calcium influx, and synaptic protein
distribution." J Neurosci 14(11 Pt 1): 6402-6411.
- 33 -
Chapter 1
References
Beattie, E. C., D. Stellwagen, et al. (2002). "Control of synaptic strength by glial TNFalpha."
Science 295(5563): 2282-2285.
Bekkers, J. M. and C. F. Stevens (1991). "Excitatory and inhibitory autaptic currents in isolated
hippocampal neurons maintained in cell culture." Proc Natl Acad Sci U S A 88(17):
7834-7838.
Bekku, Y., U. Rauch, et al. (2009). "Brevican distinctively assembles extracellular components
at the large diameter nodes of Ranvier in the CNS." J Neurochem 108(5): 1266-1276.
Bekku, Y., M. Saito, et al. (2011). "Bral2 is indispensable for the proper localization of brevican
and the structural integrity of the perineuronal net in the brain stem and cerebellum." J
Comp Neurol.
Bekku, Y., W. D. Su, et al. (2003). "Molecular cloning of Bral2, a novel brain-specific link protein,
and immunohistochemical colocalization with brevican in perineuronal nets." Mol Cell
Neurosci 24(1): 148-159.
Bliss, T. V. and G. L. Collingridge (1993). "A synaptic model of memory: long-term potentiation
in the hippocampus." Nature 361(6407): 31-39.
Blumcke, I., E. Weruaga, et al. (1994). "Discrete reduction patterns of parvalbumin and calbindin
D-28k immunoreactivity in the dorsal lateral geniculate nucleus and the striate cortex
of adult macaque monkeys after monocular enucleation." Vis Neurosci 11(1): 1-11.
Bradbury, E. J., L. D. Moon, et al. (2002). "Chondroitinase ABC promotes functional recovery
after spinal cord injury." Nature 416(6881): 636-640.
Brakebusch, C., C. I. Seidenbecher, et al. (2002). "Brevican-deficient mice display impaired
hippocampal CA1 long-term potentiation but show no obvious deficits in learning and
memory." Mol Cell Biol 22(21): 7417-7427.
Bresler, T., Y. Ramati, et al. (2001). "The dynamics of SAP90/PSD-95 recruitment to new
synaptic junctions." Mol Cell Neurosci 18(2): 149-167.
Bresler, T., M. Shapira, et al. (2004). "Postsynaptic density assembly is fundamentally different
from presynaptic active zone assembly." J Neurosci 24(6): 1507-1520.
Bruckner, G., W. Hartig, et al. (1996). "Extracellular matrix organization in various regions of rat
brain grey matter." J Neurocytol 25(5): 333-346.
Bulow, H. E. and O. Hobert (2006). "The molecular diversity of glycosaminoglycans shapes
animal development." Annu Rev Cell Dev Biol 22: 375-407.
Bury, L. A. and S. L. Sabo (2011). "Coordinated trafficking of synaptic vesicle and active zone
proteins prior to synapse formation." Neural Dev 6: 24.
- 34 -
Chapter 1
References
Bushong, E. A., M. E. Martone, et al. (2002). "Protoplasmic astrocytes in CA1 stratum radiatum
occupy separate anatomical domains." J Neurosci 22(1): 183-192.
Cahoy, J. D., B. Emery, et al. (2008). "A transcriptome database for astrocytes, neurons, and
oligodendrocytes: a new resource for understanding brain development and function." J
Neurosci 28(1): 264-278.
Calabrese, B., M. S. Wilson, et al. (2006). "Development and regulation of dendritic spine
synapses." Physiology (Bethesda) 21: 38-47.
Carmona, M. A., K. K. Murai, et al. (2009). "Glial ephrin-A3 regulates hippocampal dendritic
spine morphology and glutamate transport." Proc Natl Acad Sci U S A 106(30): 1252412529.
Carulli, D., T. Laabs, et al. (2005). "Chondroitin sulfate proteoglycans in neural development and
regeneration." Curr Opin Neurobiol 15(1): 116-120.
Carulli, D., T. Pizzorusso, et al. (2010). "Animals lacking link protein have attenuated
perineuronal nets and persistent plasticity." Brain 133(Pt 8): 2331-2347.
Carulli, D., K. E. Rhodes, et al. (2006). "Composition of perineuronal nets in the adult rat
cerebellum and the cellular origin of their components." J Comp Neurol 494(4): 559577.
Celio, M. R., R. Spreafico, et al. (1998). "Perineuronal nets: past and present." Trends Neurosci
21(12): 510-515.
Christopherson, K. S., E. M. Ullian, et al. (2005). "Thrombospondins are astrocyte-secreted
proteins that promote CNS synaptogenesis." Cell 120(3): 421-433.
Collingridge, G. L., J. T. Isaac, et al. (2004). "Receptor trafficking and synaptic plasticity." Nat
Rev Neurosci 5(12): 952-962.
Collins, M. O., H. Husi, et al. (2006). "Molecular characterization and comparison of the
components and multiprotein complexes in the postsynaptic proteome." J Neurochem
97 Suppl 1: 16-23.
Craig, A. M. and Y. Kang (2007). "Neurexin-neuroligin signaling in synapse development." Curr
Opin Neurobiol 17(1): 43-52.
Cybulska-Klosowicz, A., R. Zakrzewska, et al. (2004). "Reduced plasticity of cortical whisker
representation in adult tenascin-C-deficient mice after vibrissectomy." Eur J Neurosci
20(6): 1538-1544.
Czopka, T., A. von Holst, et al. (2010). "Regulatory mechanisms that mediate tenascin Cdependent inhibition of oligodendrocyte precursor differentiation." J Neurosci 30(37):
12310-12322.
- 35 -
Chapter 1
References
Czopka, T., A. Von Holst, et al. (2009). "Tenascin C and tenascin R similarly prevent the
formation of myelin membranes in a RhoA-dependent manner, but antagonistically
regulate the expression of myelin basic protein via a separate pathway." Glia 57(16):
1790-1801.
Dalva, M. B., M. A. Takasu, et al. (2000). "EphB receptors interact with NMDA receptors and
regulate excitatory synapse formation." Cell 103(6): 945-956.
Dean, C., F. G. Scholl, et al. (2003). "Neurexin mediates the assembly of presynaptic terminals."
Nat Neurosci 6(7): 708-716.
Dityatev, A., G. Bruckner, et al. (2007). "Activity-dependent formation and functions of
chondroitin sulfate-rich extracellular matrix of perineuronal nets." Dev Neurobiol 67(5):
570-588.
Dityatev, A. and D. A. Rusakov (2011). "Molecular signals of plasticity at the tetrapartite
synapse." Curr Opin Neurobiol 21(2): 353-359.
Dityatev, A. and M. Schachner (2003). "Extracellular matrix molecules and synaptic plasticity."
Nat Rev Neurosci 4(6): 456-468.
Dityatev, A., M. Schachner, et al. (2010). "The dual role of the extracellular matrix in synaptic
plasticity and homeostasis." Nat Rev Neurosci 11(11): 735-746.
Doege, K., M. Sasaki, et al. (1987). "Complete primary structure of the rat cartilage
proteoglycan core protein deduced from cDNA clones." J Biol Chem 262(36): 1775717767.
Ehlers, M. D. (2003). "Activity level controls postsynaptic composition and signaling via the
ubiquitin-proteasome system." Nat Neurosci 6(3): 231-242.
Einheber, S., L. M. Schnapp, et al. (1996). "Regional and ultrastructural distribution of the alpha
8 integrin subunit in developing and adult rat brain suggests a role in synaptic function."
J Comp Neurol 370(1): 105-134.
Elste, A. M. and D. L. Benson (2006). "Structural basis for developmentally regulated changes
in cadherin function at synapses." J Comp Neurol 495(3): 324-335.
Evers, M. R., B. Salmen, et al. (2002). "Impairment of L-type Ca2+ channel-dependent forms of
hippocampal synaptic plasticity in mice deficient in the extracellular matrix glycoprotein
tenascin-C." J Neurosci 22(16): 7177-7194.
Faissner, A., A. Clement, et al. (1994). "Isolation of a neural chondroitin sulfate proteoglycan
with neurite outgrowth promoting properties." J Cell Biol 126(3): 783-799.
- 36 -
Chapter 1
References
Faissner, A., M. Pyka, et al. (2010). "Contributions of astrocytes to synapse formation and
maturation - Potential functions of the perisynaptic extracellular matrix." Brain Res Rev
63(1-2): 26-38.
Feigin, I. (1980). "The mucopolysaccharides of the ground substance of the human brain." J
Neuropathol Exp Neurol 39(1): 1-12.
Feng, W. and M. Zhang (2009). "Organization and dynamics of PDZ-domain-related
supramodules in the postsynaptic density." Nat Rev Neurosci 10(2): 87-99.
Fischer, M., S. Kaech, et al. (2000). "Glutamate receptors regulate actin-based plasticity in
dendritic spines." Nat Neurosci 3(9): 887-894.
Friedlander, D. R., P. Milev, et al. (1994). "The neuronal chondroitin sulfate proteoglycan
neurocan binds to the neural cell adhesion molecules Ng-CAM/L1/NILE and N-CAM,
and inhibits neuronal adhesion and neurite outgrowth." J Cell Biol 125(3): 669-680.
Friedman, H. V., T. Bresler, et al. (2000). "Assembly of new individual excitatory synapses: time
course and temporal order of synaptic molecule recruitment." Neuron 27(1): 57-69.
Frischknecht, R., M. Heine, et al. (2009). "Brain extracellular matrix affects AMPA receptor
lateral mobility and short-term synaptic plasticity." Nat Neurosci 12(7): 897-904.
Fukazawa, Y., Y. Saitoh, et al. (2003). "Hippocampal LTP is accompanied by enhanced F-actin
content within the dendritic spine that is essential for late LTP maintenance in vivo."
Neuron 38(3): 447-460.
Garcia-Alias, G., S. Barkhuysen, et al. (2009). "Chondroitinase ABC treatment opens a window
of opportunity for task-specific rehabilitation." Nat Neurosci 12(9): 1145-1151.
Garcion, E., A. Faissner, et al. (2001). "Knockout mice reveal a contribution of the extracellular
matrix molecule tenascin-C to neural precursor proliferation and migration."
Development 128(13): 2485-2496.
Garcion, E., A. Halilagic, et al. (2004). "Generation of an environmental niche for neural stem
cell development by the extracellular matrix molecule tenascin C." Development
131(14): 3423-3432.
Garner, C. C., R. G. Zhai, et al. (2002). "Molecular mechanisms of CNS synaptogenesis." Trends
Neurosci 25(5): 243-251.
Garrett, A. M. and J. A. Weiner (2009). "Control of CNS synapse development by {gamma}protocadherin-mediated astrocyte-neuron contact." J Neurosci 29(38): 11723-11731.
Geissler, M., H. R. Dinse, et al. (2011). "Human umbilical cord blood cells restore brain damage
induced changes in rat somatosensory cortex." PLoS One 6(6): e20194.
- 37 -
Chapter 1
References
Giamanco, K. A., M. Morawski, et al. (2010). "Perineuronal net formation and structure in
aggrecan knockout mice." Neuroscience 170(4): 1314-1327.
Glegg, R. E. and R. H. Pearce (1956). "Chemical extraction of metachromatic and periodic acidSchiff positive carbohydrates from cerebral tissue." J Comp Neurol 106(2): 291-297.
Gogolla, N., P. Caroni, et al. (2009). "Perineuronal nets protect fear memories from erasure."
Science 325(5945): 1258-1261.
Goodman, L. J., J. Valverde, et al. (1996). "Regulated release and polarized localization of brainderived neurotrophic factor in hippocampal neurons." Mol Cell Neurosci 7(3): 222-238.
Graf, E. R., X. Zhang, et al. (2004). "Neurexins induce differentiation of GABA and glutamate
postsynaptic specializations via neuroligins." Cell 119(7): 1013-1026.
Grass, D., P. G. Pawlowski, et al. (2004). "Diversity of functional astroglial properties in the
respiratory network." J Neurosci 24(6): 1358-1365.
Hall, Z. W. and J. R. Sanes (1993). "Synaptic structure and development: the neuromuscular
junction." Cell 72 Suppl: 99-121.
Hannah, M. J., A. A. Schmidt, et al. (1999). "Synaptic vesicle biogenesis." Annu Rev Cell Dev Biol
15: 733-798.
Hartig, W., A. Derouiche, et al. (1999). "Cortical neurons immunoreactive for the potassium
channel Kv3.1b subunit are predominantly surrounded by perineuronal nets presumed
as a buffering system for cations." Brain Res 842(1): 15-29.
Hartig, W., A. Singer, et al. (2001). "Perineuronal nets in the rat medial nucleus of the trapezoid
body surround neurons immunoreactive for various amino acids, calcium-binding
proteins and the potassium channel subunit Kv3.1b." Brain Res 899(1-2): 123-133.
Hashimoto, K. and M. Kano (2003). "Functional differentiation of multiple climbing fiber inputs
during synapse elimination in the developing cerebellum." Neuron 38(5): 785-796.
Haubensak, W., F. Narz, et al. (1998). "BDNF-GFP containing secretory granules are localized in
the vicinity of synaptic junctions of cultured cortical neurons." Journal of Cell Science
111: 1483-1493.
Haydon, P. G., C. S. Cohan, et al. (1985). "Neuron-specific growth cone properties as seen in
identified neurons of Helisoma." J Neurosci Res 13(1-2): 135-147.
Hebb, D. O. (1949). "The Organization of Behavior; a Neuropsychological Theory." Wiley, New
York, 1949)
(reprinted by Lawrence Erlbaum Associates, 2002).
Hering, H. and M. Sheng (2001). "Dendritic spines: structure, dynamics and regulation." Nat Rev
Neurosci 2(12): 880-888.
- 38 -
Chapter 1
References
Hirrlinger, J., S. Hulsmann, et al. (2004). "Astroglial processes show spontaneous motility at
active synaptic terminals in situ." Eur J Neurosci 20(8): 2235-2239.
Huttner, W. B., M. Ohashi, et al. (1995). "Biogenesis of neurosecretory vesicles." Cold Spring
Harb Symp Quant Biol 60: 315-327.
Iozzo, R. V. (1998). "Matrix proteoglycans: from molecular design to cellular function." Annu Rev
Biochem 67: 609-652.
Irintchev, A., A. Rollenhagen, et al. (2005). "Structural and functional aberrations in the cerebral
cortex of tenascin-C deficient mice." Cereb Cortex 15(7): 950-962.
Joester, A. and A. Faissner (2001). "The structure and function of tenascins in the nervous
system." Matrix Biol 20(1): 13-22.
Kandel, E. R. (2001). "The molecular biology of memory storage: a dialog between genes and
synapses." Biosci Rep 21(5): 565-611.
Karus, M., B. Denecke, et al. (2011). "The extracellular matrix molecule tenascin C modulates
expression levels and territories of key patterning genes during spinal cord astrocyte
specification." Development 138(24): 5321-5331.
Kayser, M. S., M. J. Nolt, et al. (2008). "EphB receptors couple dendritic filopodia motility to
synapse formation." Neuron 59(1): 56-69.
Kerchner, G. A. and R. A. Nicoll (2008). "Silent synapses and the emergence of a postsynaptic
mechanism for LTP." Nat Rev Neurosci 9(11): 813-825.
Knott, G. W., C. Quairiaux, et al. (2002). "Formation of dendritic spines with GABAergic synapses
induced by whisker stimulation in adult mice." Neuron 34(2): 265-273.
Kwok, J. C., D. Carulli, et al. (2010). "In vitro modeling of perineuronal nets: hyaluronan
synthase and link protein are necessary for their formation and integrity." J Neurochem
114(5): 1447-1459.
Kwok, J. C., G. Dick, et al. (2011). "Extracellular matrix and perineuronal nets in CNS repair."
Dev Neurobiol.
Lander, C., P. Kind, et al. (1997). "A family of activity-dependent neuronal cell-surface
chondroitin sulfate proteoglycans in cat visual cortex." J Neurosci 17(6): 1928-1939.
Lee, H., C. Dean, et al. (2010). "Alternative splicing of neuroligin regulates the rate of
presynaptic differentiation." J Neurosci 30(34): 11435-11446.
LeVay, S., T. N. Wiesel, et al. (1980). "The development of ocular dominance columns in normal
and visually deprived monkeys." J Comp Neurol 191(1): 1-51.
Lichtman, J. W. and H. Colman (2000). "Synapse elimination and indelible memory." Neuron
25(2): 269-278.
- 39 -
Chapter 1
References
Lohmann, C., A. Finski, et al. (2005). "Local calcium transients regulate the spontaneous
motility of dendritic filopodia." Nat Neurosci 8(3): 305-312.
Lovell, P., B. McMahon, et al. (2002). "Synaptic precedence during synapse formation between
reciprocally connected neurons involves transmitter-receptor interactions and AA
metabolites." J Neurophysiol 88(3): 1328-1338.
Malenka, R. C. (2003). "Synaptic plasticity and AMPA receptor trafficking." Ann N Y Acad Sci
1003: 1-11.
Matthews, R. T., G. M. Kelly, et al. (2002). "Aggrecan glycoforms contribute to the molecular
heterogeneity of perineuronal nets." J Neurosci 22(17): 7536-7547.
Mauch, D. H., K. Nagler, et al. (2001). "CNS synaptogenesis promoted by glia-derived
cholesterol." Science 294(5545): 1354-1357.
McAllister, A. K. (2007). "Dynamic aspects of CNS synapse formation." Annu Rev Neurosci 30:
425-450.
McKinney, R. A., M. Capogna, et al. (1999). "Miniature synaptic events maintain dendritic spines
via AMPA receptor activation." Nat Neurosci 2(1): 44-49.
Milev, P., P. Maurel, et al. (1998). "Differential regulation of expression of hyaluronan-binding
proteoglycans in developing brain: aggrecan, versican, neurocan, and brevican."
Biochem Biophys Res Commun 247(2): 207-212.
Mjaatvedt, C. H., H. Yamamura, et al. (1998). "The Cspg2 gene, disrupted in the hdf mutant, is
required for right cardiac chamber and endocardial cushion formation." Dev Biol 202(1):
56-66.
Moeller, M. L., Y. Shi, et al. (2006). "EphB receptors regulate dendritic spine morphogenesis
through the recruitment/phosphorylation of focal adhesion kinase and RhoA activation."
J Biol Chem 281(3): 1587-1598.
Nimchinsky, E. A., B. L. Sabatini, et al. (2002). "Structure and function of dendritic spines." Annu
Rev Physiol 64: 313-353.
Nishiyama, A., M. Komitova, et al. (2009). "Polydendrocytes (NG2 cells): multifunctional cells
with lineage plasticity." Nat Rev Neurosci 10(1): 9-22.
O'Brien, R. J., D. Xu, et al. (1999). "Synaptic clustering of AMPA receptors by the extracellular
immediate-early gene product Narp." Neuron 23(2): 309-323.
Okabe, S., T. Urushido, et al. (2001). "Rapid redistribution of the postsynaptic density protein
PSD-Zip45 (Homer 1c) and its differential regulation by NMDA receptors and calcium
channels." J Neurosci 21(24): 9561-9571.
- 40 -
Chapter 1
References
Olin, A. I., M. Morgelin, et al. (2001). "The proteoglycans aggrecan and Versican form networks
with fibulin-2 through their lectin domain binding." J Biol Chem 276(2): 1253-1261.
Palay, S. L. (1956). "Synapses in the central nervous system." J Biophys Biochem Cytol 2(4
Suppl): 193-202.
Penzes, P., A. Beeser, et al. (2003). "Rapid induction of dendritic spine morphogenesis by transsynaptic ephrinB-EphB receptor activation of the Rho-GEF kalirin." Neuron 37(2): 263274.
Perlini, L. E., F. Botti, et al. (2011). "Effects of phosphorylation and neuronal activity on the
control of synapse formation by synapsin I." J Cell Sci 124(Pt 21): 3643-3653.
Pizzorusso, T., P. Medini, et al. (2002). "Reactivation of ocular dominance plasticity in the adult
visual cortex." Science 298(5596): 1248-1251.
Pyka, M., C. Busse, et al. (2011). "Astrocytes are crucial for survival and maturation of
embryonic hippocampal neurons in a neuron-glia cell-insert coculture assay." Synapse
65(1): 41-53.
Pyka, M., C. Wetzel, et al. (2011). "Chondroitin sulfate proteoglycans regulate astrocytedependent synaptogenesis and modulate synaptic activity in primary embryonic
hippocampal neurons." Eur J Neurosci 33(12): 2187-2202.
Rakic, P. and R. L. Sidman (1970). "Histogenesis of cortical layers in human cerebellum,
particularly the lamina dissecans." J Comp Neurol 139(4): 473-500.
Rauch, U., L. Karthikeyan, et al. (1992). "Cloning and primary structure of neurocan, a
developmentally regulated, aggregating chondroitin sulfate proteoglycan of brain." J
Biol Chem 267(27): 19536-19547.
Rauch, U., X. H. Zhou, et al. (2005). "Extracellular matrix alterations in brains lacking four of its
components." Biochem Biophys Res Commun 328(2): 608-617.
Ryan, T. J. and S. G. Grant (2009). "The origin and evolution of synapses." Nat Rev Neurosci
10(10): 701-712.
Sabo, S. L., R. A. Gomes, et al. (2006). "Formation of presynaptic terminals at predefined sites
along axons." J Neurosci 26(42): 10813-10825.
Saghatelyan, A. K., S. Gorissen, et al. (2000). "The extracellular matrix molecule tenascin-R and
its HNK-1 carbohydrate modulate perisomatic inhibition and long-term potentiation in
the CA1 region of the hippocampus." Eur J Neurosci 12(9): 3331-3342.
Sanes, J. R. and J. W. Lichtman (1999). "Development of the vertebrate neuromuscular
junction." Annu Rev Neurosci 22: 389-442.
- 41 -
Chapter 1
References
Scheiffele, P. (2003). "Cell-cell signaling during synapse formation in the CNS." Annu Rev
Neurosci 26: 485-508.
Scheiffele, P., J. Fan, et al. (2000). "Neuroligin expressed in nonneuronal cells triggers
presynaptic development in contacting axons." Cell 101(6): 657-669.
Schwarting, G. A., C. Kostek, et al. (2000). "Semaphorin 3A is required for guidance of olfactory
axons in mice." J Neurosci 20(20): 7691-7697.
Shapira, M., R. G. Zhai, et al. (2003). "Unitary assembly of presynaptic active zones from
Piccolo-Bassoon transport vesicles." Neuron 38(2): 237-252.
Shen, L. H., Y. Li, et al. (2008). "Down-regulation of neurocan expression in reactive astrocytes
promotes axonal regeneration and facilitates the neurorestorative effects of bone
marrow stromal cells in the ischemic rat brain." Glia 56(16): 1747-1754.
Singhal, N. and P. T. Martin (2011). "Role of extracellular matrix proteins and their receptors in
the development of the vertebrate neuromuscular junction." Dev Neurobiol 71(11): 9821005.
Slezak, M. and F. W. Pfrieger (2003). "New roles for astrocytes: regulation of CNS
synaptogenesis." Trends Neurosci 26(10): 531-535.
Song, I. and R. L. Huganir (2002). "Regulation of AMPA receptors during synaptic plasticity."
Trends Neurosci 25(11): 578-588.
Speese, S. D., N. Trotta, et al. (2003). "The ubiquitin proteasome system acutely regulates
presynaptic protein turnover and synaptic efficacy." Curr Biol 13(11): 899-910.
Spencer, G. E., K. Lukowiak, et al. (2000). "Transmitter-receptor interactions between growth
cones of identified Lymnaea neurons determine target cell selection in vitro." J Neurosci
20(21): 8077-8086.
Sperry, R. W. (1963). "Chemoaffinity in the Orderly Growth of Nerve Fiber Patterns and
Connections." Proc Natl Acad Sci U S A 50: 703-710.
Sudhof, T. C. (2000). "The synaptic vesicle cycle revisited." Neuron 28(2): 317-320.
Sun, M., G. Xing, et al. (2011). "Neuroligin 2 is required for synapse development and function at
the Drosophila neuromuscular junction." J Neurosci 31(2): 687-699.
Sykova, E., I. Vorisek, et al. (2005). "Reduced extracellular space in the brain of tenascin-R- and
HNK-1-sulphotransferase deficient mice." Eur J Neurosci 22(8): 1873-1880.
Tao-Cheng, J. H. (2007). "Ultrastructural localization of active zone and synaptic vesicle
proteins in a preassembled multi-vesicle transport aggregate." Neuroscience 150(3):
575-584.
Temple, S. (2001). "The development of neural stem cells." Nature 414(6859): 112-117.
- 42 -
Chapter 1
References
Togashi, H., K. Abe, et al. (2002). "Cadherin regulates dendritic spine morphogenesis." Neuron
35(1): 77-89.
Toole, B. P. (2001). "Hyaluronan in morphogenesis." Semin Cell Dev Biol 12(2): 79-87.
Trachtenberg, J. T., B. E. Chen, et al. (2002). "Long-term in vivo imaging of experiencedependent synaptic plasticity in adult cortex." Nature 420(6917): 788-794.
Tsai, H. H., E. Frost, et al. (2002). "The chemokine receptor CXCR2 controls positioning of
oligodendrocyte precursors in developing spinal cord by arresting their migration." Cell
110(3): 373-383.
Turnbull, J., A. Powell, et al. (2001). "Heparan sulfate: decoding a dynamic multifunctional cell
regulator." Trends Cell Biol 11(2): 75-82.
Ullian, E. M., K. S. Christopherson, et al. (2004). "Role for glia in synaptogenesis." Glia 47(3):
209-216.
Umemori, H., M. W. Linhoff, et al. (2004). "FGF22 and its close relatives are presynaptic
organizing molecules in the mammalian brain." Cell 118(2): 257-270.
Van Obberghen-Schilling, E., R. P. Tucker, et al. (2011). "Fibronectin and tenascin-C:
accomplices in vascular morphogenesis during development and tumor growth." Int J
Dev Biol 55(4-5): 511-525.
Vaughn, J. E. (1989). "Fine structure of synaptogenesis in the vertebrate central nervous
system." Synapse 3(3): 255-285.
Verderio, C., S. Coco, et al. (1999). "Synaptogenesis in hippocampal cultures." Cell Mol Life Sci
55(11): 1448-1462.
Verhage, M., A. S. Maia, et al. (2000). "Synaptic assembly of the brain in the absence of
neurotransmitter secretion." Science 287(5454): 864-869.
Vicario-Abejon, C., D. Owens, et al. (2002). "Role of neurotrophins in central synapse formation
and stabilization." Nat Rev Neurosci 3(12): 965-974.
Virgintino, D., D. Perissinotto, et al. (2009). "Differential distribution of aggrecan isoforms in
perineuronal nets of the human cerebral cortex." J Cell Mol Med 13(9B): 3151-3173.
Vitellaro-Zuccarello, L., P. Bosisio, et al. (2007). "Differential expression of several molecules of
the extracellular matrix in functionally and developmentally distinct regions of rat
spinal cord." Cell Tissue Res 327(3): 433-447.
Volterra, A. and J. Meldolesi (2005). "Astrocytes, from brain glue to communication elements:
the revolution continues." Nat Rev Neurosci 6(8): 626-640.
von Holst, A., U. Egbers, et al. (2007). "Neural stem/progenitor cells express 20 tenascin C
isoforms that are differentially regulated by Pax6." J Biol Chem 282(12): 9172-9181.
- 43 -
Chapter 1
References
Waites, C. L., A. M. Craig, et al. (2005). "Mechanisms of vertebrate synaptogenesis." Annu Rev
Neurosci 28: 251-274.
Wallraff, A., B. Odermatt, et al. (2004). "Distinct types of astroglial cells in the hippocampus
differ in gap junction coupling." Glia 48(1): 36-43.
Wang, D., R. M. Ichiyama, et al. (2011). "Chondroitinase combined with rehabilitation promotes
recovery of forelimb function in rats with chronic spinal cord injury." J Neurosci 31(25):
9332-9344.
Washbourne, P., J. E. Bennett, et al. (2002). "Rapid recruitment of NMDA receptor transport
packets to nascent synapses." Nat Neurosci 5(8): 751-759.
Washbourne, P., X. B. Liu, et al. (2004). "Cycling of NMDA receptors during trafficking in neurons
before synapse formation." J Neurosci 24(38): 8253-8264.
Watanabe, H., K. Kimata, et al. (1994). "Mouse cartilage matrix deficiency (cmd) caused by a 7
bp deletion in the aggrecan gene." Nat Genet 7(2): 154-157.
Watt, F. M. and K. J. Hodivala (1994). "Cell adhesion. Fibronectin and integrin knockouts come
unstuck." Curr Biol 4(3): 270-272.
Weber, P., U. Bartsch, et al. (1999). "Mice deficient for tenascin-R display alterations of the
extracellular matrix and decreased axonal conduction velocities in the CNS." J Neurosci
19(11): 4245-4262.
Wegner, F., W. Hartig, et al. (2003). "Diffuse perineuronal nets and modified pyramidal cells
immunoreactive for glutamate and the GABA(A) receptor alpha1 subunit form a unique
entity in rat cerebral cortex." Exp Neurol 184(2): 705-714.
Wilkinson, D. G. (2001). "Multiple roles of EPH receptors and ephrins in neural development."
Nat Rev Neurosci 2(3): 155-164.
Yamada, H., K. Watanabe, et al. (1994). "Molecular cloning of brevican, a novel brain
proteoglycan of the aggrecan/versican family." J Biol Chem 269(13): 10119-10126.
Yamaguchi, Y. (2000). "Lecticans: organizers of the brain extracellular matrix." Cell Mol Life Sci
57(2): 276-289.
Yamamoto, S., S. Oka, et al. (2002). "Mice deficient in nervous system-specific carbohydrate
epitope HNK-1 exhibit impaired synaptic plasticity and spatial learning." J Biol Chem
277(30): 27227-27231.
Yuste, R. (2011). "Dendritic spines and distributed circuits." Neuron 71(5): 772-781.
Yuste, R. and T. Bonhoeffer (2004). "Genesis of dendritic spines: insights from ultrastructural
and imaging studies." Nat Rev Neurosci 5(1): 24-34.
- 44 -
Chapter 1
References
Zhai, R. G., H. Vardinon-Friedman, et al. (2001). "Assembling the presynaptic active zone: a
characterization of an active one precursor vesicle." Neuron 29(1): 131-143.
Zhou, X. H., C. Brakebusch, et al. (2001). "Neurocan is dispensable for brain development." Mol
Cell Biol 21(17): 5970-5978.
Zimmermann, D. R. and E. Ruoslahti (1989). "Multiple domains of the large fibroblast
proteoglycan, versican." EMBO J 8(10): 2975-2981.
Ziv, N. E. and C. C. Garner (2004). "Cellular and molecular mechanisms of presynaptic
assembly." Nat Rev Neurosci 5(5): 385-399.
- 45 -
Chapter 2
Objectives
Chapter 2
Objectives
The extracellular matrix, released by glia cells and neurons, is thought to be crucially
involved in the development of neuronal networks, especially in the formation and
maintenance of synapses. Perineuronal nets (PNNs) were recently suggested to be one
of the key ECM structures for synapse stabilization and maintenance. Several studies
used enzymatic matrix digestion via Chondroitinase ABC (ChABC) treatment in vivo and
in vitro and verified this hypothesis. However, the detailed mechanisms remain elusive.
A developmental genetic deficiency of major matrix components is thought to shed
light on these processes and would provide a strong tool for unraveling the cell-matrix
interaction at the CNS synapse. Therefore, we utilized a quadruple knock-out mouse,
lacking tenascin-C, tenascin-R, neurocan and brevican. This mutant is viable and fertile
without gross abnormalities, thereby representing a versatile approach to study the
consequences of the matrix deficiency in detail.
The present study comprises two main parts, structured in chapter 3 and chapter 4.
In chapter 3, a manuscript that is being prepared for submission, which presents the
main work of the thesis, is embedded. This project aimed to intensively characterize the
development of matrix deficient hippocampal neurons with regard to PNN- and synapse
formation and the consequences for the network formation and the synaptic
transmission. We have chosen an in vitro set up, where hippocampal neurons and
supporting astrocytes can communicate via the medium, without direct membrane
mediated contact. This allows for the long-term cultivation of primary hippocampal
neurons and the combinatorial use of wild type and mutant cells, as well as for the
separation of the glial and neuronal contribution to the matrix assembly. For analysis
we combined immunocytochemical approaches and electrophysiological whole-cell
patch clamp recordings.
Chapter 4 deals with the establishment of a new method for cultivating primary
hippocampal mouse neurons in indirect co-culture with astrocytes on multi electrode
- 46 -
Chapter 2
Objectives
arrays (MEAs). The aim of this part was to establish the MEA technology in our lab and
to transfer the indirect co-culture assay, utilized in chapter 3, to MEAs.
The use of primary mouse cells on the MEA allows now for the use of genetic mutants
and will provide new insights into the ECM dependent development on a network wide
level.
This work has recently been published in the Journal of Neuroscience Methods
(Geissler and Faissner, 2011).
- 47 -
Chapter 3
Chapter 3
Primary hippocampal neurons, which lack four crucial
extracellular matrix molecules, display abnormalities
of synaptic structure and function and severe deficits in
perineuronal net formation
Maren Geissler1, Ainhara Aguado2, Uwe Rauch3, Christian H. Wetzel2, Hanns Hatt2, Andreas
Faissner1
1 Department of Cell Morphology and Molecular Neurobiology, Ruhr-University, D-44780 Bochum,
Germany
2 Department of Cell Physiology, Ruhr-University, D-44780 Bochum, Germany
3 Department of Experimental Medical Science, Lund, Sweden
3.1 Abstract
The extracellular matrix (ECM) of the brain plays a diverse and crucial role during
development, maturation and regeneration of the central nervous system (CNS).
Nevertheless, there remain many open questions, especially with regard to the role of
the ECM at the CNS synapse. Here we show for the first time via a morphological and
electrophysiological characterization of primary hippocampal neurons lacking four
matrix molecules significant changes in synaptic transmission, in perineuronal net
(PNN) formation and maintenance, and in synapse formation and stabilization as a
consequence of ECM deficiency. This unique study yields new important findings
concerning the involvement of the ECM in synapse formation, maturation and function
and
provides
new
knowledge
concerning
the
bidirectional
neuron-astrocyte
communication. Further, we present first insights into the neuronal phenotype of PNN
deficiency, which results in diminished expression of synaptic proteins.
- 48 -
Chapter 3
3.2 Introduction
Astrocytes by far outnumber neurons in the brain and the research of the last decades
shed light on the complexity of neuron-glia interactions, which reaches far beyond the
structural support and the delivery of nutrients (Di Castro et al., 2011; Navarrete and
Araque, 2011; Perea et al., 2009; Porto-Pazos et al., 2011). Rather, astrocytes and
neurons form intimate contacts, especially at the central synapse, resulting in a
tripartite structure (Haydon, 2001; Perea et al., 2009; Slezak and Pfrieger, 2003), in
which the presynapse and the postsynaptic membrane are arranged closely to the
astrocytic processes. Owing to that close neighborhood, astrocytes are capable of
affecting synaptic strength and plasticity (Beattie et al., 2002; Theodosis et al., 2008).
Moreover, astrocytes have been shown to be indispensable for neuronal survival,
synapse formation and function (Beattie et al., 2002; Di Castro et al., 2011; Nagler et al.,
2001; Navarrete and Araque, 2011; Porto-Pazos et al., 2011; Pyka et al., 2011a; Pyka et
al., 2011b). Thus, neurons growing in defined medium without supporting glia cells,
show diminished synaptic activity (Boehler et al., 2007; Christopherson et al., 2005) or
higher rates of cell death (Pyka et al., 2011a). Beside the direct, membrane-mediated
contact astrocytes are known to secret soluble factors promoting the neuron´s viability
and the formation and stabilization of the synapse (Gomes et al., 2001; Lafon-Cazal et
al., 2003). Proposed secreted candidate molecules involved in these processes are
neurotrophins (Cahoy et al., 2008; Elmariah et al., 2005) and cholesterol (Mauch et al.,
2001). Furthermore, astrocytes are supposed to release molecules that assemble the
extracellular matrix (ECM) and thus exert further local impact on neuronal network
formation and maintenance (Carulli et al., 2005; Faissner et al., 2010; Frischknecht et
al., 2009; Pyka et al., 2011b). Thus, thrombospondins (Christopherson et al., 2005),
Chondroitin sulfate proteoglycans (CSPGs) (Pyka et al., 2011b) and the two
matricellular proteins Hevin and SPARC (Kucukdereli et al., 2011) are known to be
involved in the assembly of the synaptic machinery. In line with this, the enzymatic
digestion of astrocyte released ECM components with Chondroitinase ABC was shown
to significantly increase synaptic puncta formation in rat hippocampal neurons in vitro
(Pyka et al., 2011b). This observation indicates a context-dependent, regulatory,
- 49 -
Chapter 3
repulsive and inhibitory impact the ECM can exert, in contrast to its attractive and
synapse enhancing properties (Hartley et al., 1999; van den Pol and Spencer, 2000).
Prominent neural ECM molecules are the lecticans of the CSPG family neurocan,
brevican, versican and aggrecan, as well as the glycoproteins tenascin-C and tenascinR. Mutations in these genes are a versatile tool to investigate the matrix´ role in the
respective developmental processes. While the mutations of aggrecan and versican are
lethal (Mjaatvedt et al., 1998; Watanabe et al., 1994), the single knock-out mice of the
other genes show moderate phenotypes, revealing the proteins´ involvement in
processes like LTP-induction and synaptic plasticity. Thus, mice deficient for tenascin-C
show reduced LTP in the CA1 region (Evers et al., 2002) and diminished plasticity after
vibrissectomy (Cybulska-Klosowicz et al., 2004). Mutant mice lacking tenascin-R do
also exhibit impaired LTP (Bukalo et al., 2001; Saghatelyan et al., 2001) and show a
metaplastic increase in the threshold for the induction of LTP (Bukalo et al., 2007).
Interestingly, both tenascin mutants carry a phenotype in the inhibitory system of the
brain. A reduced perisomatic GABAergic inhibition (Saghatelyan et al., 2000) and
impaired GABA release at perisomatic inhibitory synapses (Nikonenko et al., 2003)
were described in the tenascin-R mutant, while a decreased density of parvalbuminpositive neurons and a low ratio of inhibitory to excitatory neurons in the cortex of
tenascin C deficient mice (Irintchev et al., 2005) was found. From experiments with
mice lacking one of the lecticans stems further evidence for the involvement of the
ECM in synaptic plasticity and neuronal development (Brakebusch et al., 2002), (for
review see Faissner et al., 2010). Nevertheless, the precise mechanisms that contribute
to the complex interplay between astrocytes and neurons in the context of the
extracellular matrix molecules have still to be unraveled.
A specialized form of the matrix that is tightly associated with the neuronal cell body
and proximal dendrites is the perineuronal net (PNN) (Bruckner et al., 2000; Carulli et
al., 2006; Celio et al., 1998). PNNs are defined net-like structures of condensed
extracellular matrix molecules that accumulate around a subpopulation of neurons
(Carulli et al., 2006; Dityatev et al., 2007; Galtrey et al., 2008). Parvalbumin-positive
interneurons (Wintergerst et al., 1996) as well as a minority of glutamatergic neurons
are enwrapped by these ECM structures (Wegner et al., 2003). Despite the first
- 50 -
Chapter 3
description decades ago in the 19th century by Golgi and colleagues (Celio et al., 1998),
PNN functions until recently remain enigmatic (Carulli et al., 2010; Gogolla et al., 2009;
Kwok et al., 2010; Kwok et al., 2011).These lattice-like ECM structures are supposed to
play an important role during the maturation and maintenance of the neuronal network
(Dityatev et al., 2007; Gogolla et al., 2009; Pizzorusso et al., 2002). In line with this,
PNNs appear in their mature form at the end of the critical periods (Gogolla et al., 2009;
Guimaraes et al., 1990; Pizzorusso et al., 2002), when plasticity has to be restricted and
the CNS structures need to be stabilized. Thus, the enzymatic digestion of PNNs leads
to prolonged phases of plasticity (Gogolla et al., 2009; Pizzorusso et al., 2002).
However, the consequences of altered PNN formation for the cellular level are still
unclear. Therefore, further experimental evidence is needed, in particular with regard to
approaches that exploit these structures as a point of intervention for regenerative
treatments after CNS injury (Kwok et al., 2011).
The disappearance of dense PNN structures in the cortex and hippocampus has been
described for the quadruple knock-out mice, which lack the four matrix molecules
tenascin-C, tenascin-R, neurocan and brevican (Rauch et al., 2005). This study
demonstrated the flexibility of the extracellular matrix as two matrix molecules
untypical for the CNS became up-regulated in response to the mutation, namely fibulin1 and fibulin-2 (Rauch et al., 2005). Beyond the reduced PNN expression and the
reorganization of the ECM, no gross alterations in this mutant have so far been
described. Therefore, this mutant represents a strong tool to obtain further insights into
matrix-dependent plasticity.
To this end, we investigated primary hippocampal neurons derived from E15.5
quadruple mutant mice in a defined neuron-astrocyte co-culture set up (Geissler and
Faissner, 2011; Pyka et al., 2011a) and performed a complex in vitro analysis on the
morphological and physiological level. We found a dramatic reduction in PNN number
and complexity in vitro, accompanied by a significant decrease in miniature
postsynaptic current frequency, changes in synapse formation, especially on PNN
coated neurons and a phenotype related to the inhibitory cell population.
Taken together, this study provides new important findings concerning the involvement
of the ECM in synapse development.
- 51 -
Chapter 3
3.3 Material and methods
3.3.1 Ethical standards and animal housing
All experiments were approved by the animal care and utilization committees from the
Ruhr University Bochum and were performed in accordance with the Society for
Neuroscience and the German guidelines.
Animals were reared under standardized housing conditions with 24 hour light/dark
cycle and regulated humidity. Food and water were available ad libitum.
A colony of the Quadruple knock-out mouse (Rauch, Zhou et al. 2005) is kept in the
animal house of the faculty for biology and biotechnology of the Ruhr University
Bochum.
3.3.2 Immunological reagents
The following listed antibodies were used in this study in the declared dilutions.
The following polyclonal antibodies (all rabbit) were used: anti-neurocan (1:200,
prepared by U. Rauch, Lund, Sweden (Haas, Rauch et al. 1999)) anti-brevican
(Seidenbecher, Richter et al. 1995) (1:300), for the detection of tenascin-C (batch KAF
14; (Faissner and Kruse 1990)) (1:200) was used, anti-bassoon (tom Dieck, SanmartiVila et al. 1998) (kind gift from E. Gundelfinger, 1 : 2000) and anti-neurofilament 200
(NF 200, Sigma-Aldrich, St Louis, MO, USA, 1 : 300).
The following mouse monoclonal antibodies were used: anti-postsynaptic density
protein 95 (PSD95, Calbiochem, La Jolla, CA, USA, 1 : 500), anti-β III tubulin (SigmaAldrich 1 : 200 for ICC, 1 : 750 in WB), anti-tenascin-R (clone 23-14 (Rathjen, Wolff et
al. 1991)) (1 : 200, kind gift from F.G. Rathjen), anti-Glutamic acid decarboxylase 65 and
67 (GAD 65/67, Stressgen, Ann Arbor, MI, USA, 1 : 1000) and anti-vesicular glutamate
transporter (vGLUT, Medimabs, Quebec, Canada 1 : 1000). For the detection of PNNs
we used Wisteria floribunda agglutinin (WFA, Sigma-Aldrich; 1 : 100).
All Secondary antibodies were derived from Dianova (Hamburg, Germany) and subclass
specific CY2, CY3 or CY5 anti-rabbit or anti-mouse as well as streptavidin coupled CY2
- 52 -
Chapter 3
or CY3 were used to reveal primary antibody or WFA binding. Cell nuclei were detected
via bisbenzimide (Hoechst 33528, Sigma-Aldrich, 1 : 100 000).
3.3.3 Cell culture
Neurons and astrocytes were cultivated in an indirect co-culture assay without direct
contact, but sharing the same, defined medium (Pyka, Busse et al. 2011). Both cell
types were derived as described previously (Goslin and Banker 1989; Kaech and Banker
2006; Michele and Faissner 2009) with minor modifications, indicated below.
For the preparation of astrocytes, postnatal mice (P1-P3, NMRI and quadruple knockout mice (Rauch, Zhou et al. 2005)) were decapitated, cortices were removed and
separated from the meninges and surrounding tissue. After enzymatic digestion of the
cortices with 0.1% w/v papain (Worthington, Lakewood, NJ, USA) in MEM (Invitrogen,
San Diego, CA, USA) for one hour, the tissue was triturated mechanically and
centrifugated. The resulting cell pellet was resuspended in 1 ml astrocyte medium
(DMEM (Invitrogen) with 10% v/v horse serum (Biochrom, Berlin, Germany) and 1% v/v
penicillin/streptomycin (Gibco, Karlsruhe, Germany)). The obtained single cell
suspension was added to 10 ml astrocyte medium in T-75 flasks (Nunc, Roskilde,
Denmark), pre-coated with 10 µg/ml Ploy-D-Lysine (Sigma-Aldrich) with a density of
four cortices per flask. The cells were grown for at least 7 DIV at 37°C with 6 % v/v CO2,
while a complete medium change was performed every second day. To get rid of
progenitor and oligodendrocyte precursor cells and to achieve a confluent layer of
astrocytes, flasks were shaken over night on an orbital shaker (New Brunswick, Edison,
NJ, USA) at 37°C and 250 rpm, followed by the addition of 20 μM cytosine-1-ß-D
arabinofuranosid (AraC, Sigma-Aldrich) for two days. 24-48 hours before the
preparation of neurons was undertaken (see below), astrocytes were replated in cell
culture inserts (BD Biosciences San Jose, CA, USA, pore size 0.4 µm) via enzymatic
digestion with trypsin (0.25% w/v trypsin with EDTA in MEM (both from Invitrogen)) at a
density of 25000 cells per insert.
Hippocampal neurons were derived from embryonic mice (E15.5) of time pregnant
NMRI and quadruple knock-out mice (Rauch, Zhou et al. 2005). Hippocampi were
dissected and separated from meninges and surrounding tissue before enzymatic
- 53 -
Chapter 3
digestion with 0.25% w/v trypsin (Invitrogen) in MEM (Invitrogen) for 15 minutes. After
mechanical trituration and centrifugation the resulting cell pellet was resuspended in
hippocampus medium, containing MEM (Invitrogen), 10 mM sodium pyruvate (SigmaAldrich), 0.1% w/v ovalbumin (Sigma-Aldrich) and 2% v/v B27 (Invitrogen)). Afterwards,
1200 cells were plated out at a low density of 3500 cells /cm2 on glass cover slips
(Thermo Scientific, Pittsburgh, PA, USA), coated with 15 µg/ ml Polyornithin (SigmaAldrich) in 24-well plates (BD Biosciences). Hippocampal neurons were cultivated at
37°C and 6 % v/v CO2 in a humidified incubator.
After an adherence time of two hours, the cell culture inserts with astrocytes were
added and the astrocyte medium was substituted by hippocampus medium.
Four different genotype combinations (wilde-type:
wt/wt
, knock-out:
ko/ko
) of neurons (N)
and astrocytes (A) were used in the study: both cells from wild-type: Awt/wt|Nwt/wt, both
cells from knock-out Ako/ko|Nko/ko, astrocytes from knock-out and neurons from wildtype Ako/ko|Nwt/wt and astrocytes from wild-type and neurons from knock-out
Awt/wt|Nko/ko.
3.3.4 Electrophysiology
Primary embryonic (E15.5) hippocampal neurons of the four cell culture combinations
were recorded in the whole-cell configuration using the patch-clamp technique.
Membrane voltage was controlled and currents were measured using an L/M-EPC7
patch-clamp amplifier (List, Darmstadt, Germany). Borosilicate glass (GB150EFT-10,
Science products, Hofheim, Germany) pipettes were pulled with a horizontal pipette
puller (DMZ Universal Puller, Zeitz Instruments, Munich, Germany). The patch pipettes
showed resistances ranging from 3 to 6 MΩ and hippocampal neurons were voltageclamped at -60 mV. Signals were filtered from 1.0 to 3.0 kHz. Series resistance and cell
capacitance were compensated prior to the recordings.
The patch pipettes were filled with an intracellular solution containing (in mM): 140
KCl, 2 MgCl2, 11 EGTA, 1 CaCl2, 2 HEPES (pH = 7.4, 290~310 mOsm). The standard
bath solution contained (in mM): 145 NaCl, 5 KCl, 2 CaCl2, 1 MgCl2, 10 HEPES (pH =
7.3, 290~310 mOsm) and all recordings were performed at room temperature (20 to 24
°C).
- 54 -
Chapter 3
For the measurements of miniature postsynaptic currents (mPSCs) pharmacological
isolation was required. Picrotoxin (150 μM) was added on bath solutions of all four cell
culture combinations to block GABAA receptor–mediated currents that allowed the
recording of miniature excitatory postsynaptic currents (mEPSCs). The AMPA and
Kainate receptor-mediated currents blocker 6,7-dinitroquinoxaline-2,3-dione (DNQX, 10
µM) and the voltage-gated sodium currents blocker tetrodotoxin (TTX, 1 µM) were
added on the bath solutions of all four cell combinations for the measurement of
miniature inhibitory postsynaptic currents (mIPSCs).
Acquisition of the postsynaptic events was performed by the Pulse Software (HEKA
Instruments). After conversion of the data in the right format (ABF Utility of
Minianalysis program, Synaptosoft), specific functional parameters of mEPSCs and
mIPSCs, i.e. rise and decay time (kinetic), amplitude, charge (which represents the area
under the peak of the postsynaptic event), and frequency were analyzed with the
Minianalysis programm (Synaptosoft, version 6.0.3). Only cells with consistent gigaseal
and series resistances throughout the experiment were included in the analysis.
3.3.5 Immunocytochemistry
For the immunofluorescent staining of neurons, the medium was carefully aspirated
and the cells were fixed with 4% w/v paraformaldehyde (PFA, Sigma-Aldrich) for 10
minute, rinsed with PBS and washed twice with PBS/T (PBS with 0.1 %⁄ v Triton X
-
100 (Sigma-Aldrich)). Primary antibodies were diluted in PBS/T and incubated for 30
minutes, followed by three times washing with PBS/A (PBS with 0.1 % w/v BSA from
Sigma-Aldrich). Secondary antibodies were diluted in PBS/A and incubated for 30
minutes. Thereafter, the cells were washed twice with PBS and once with Milli Q water
before covering the cells on microscope slides with Immumount (both from Thermo
Scientific).
For the immunocytochemical detection of synaptic puncta, the cells were fixed as
described above. The fixation of cells was followed by three washing steps with PBS,
15 minute incubation of 25 mM glycin (Sigma-Aldrich) and one hour blocking with a
blocking buffer containing 10% v/v horse serum (Biochrom, Berlin, Germany) and 0.1%
v/v Triton X-100 (Sigma-Aldrich). Primary antibodies in blocking buffer were incubated
- 55 -
Chapter 3
for 60 minutes, followed by three washing steps with PBS and secondary antibody
incubation in blocking buffer for 60 minutes. Afterwards, the cover slips were washed
three times with PBS and rinsed with Milli Q water before covering in Immumount on
microscope slides (both from Thermo Scientific).
3.3.6 Western Blotting
After a cultivation time of 14 and 21 DIV, neurons were washed twice with ice cold PBS
after the medium was carefully aspirated. Thereafter, the cells were collected on ice
with ice
cold
lysis
buffer
(150
mMTris/HCl pH 7.4, 150
mM NaCl, 5
ethylenediaminetetraacetic acid (EDTA), 5 mM EGTA, 1% v/v Triton X100, 0.1% w/v Nadeoxycholate and 0.1% v/v sodium dodecyl sulfate (SDS)). Protein lysates were cleared
via centrifugation at 4 °C and 16000 g for 20 minutes. The protein concentration of
probes was determined using a protein concentration kit (Pierce, Rockfort) and the
tubes were stored at -20 °C. For the SDS-PAGE, 20 µg of the proteins from each probe
were separated on a 7% polyacrylamide gel under reducing conditions. Afterwards,
protein bands were transferred to a polyvinylidene fluoride (PVDF) membrane (Roth,
Karlsruhe, Germany). Membranes were blocked by incubation with 5% w/v milk powder
in TBST. For immunodetection, the PVDF membranes were incubated overnight with
primary antibodies (see above) in 5% w/v milk powder in TBST. Next day, the
appropriate secondary antibodies (horseradish peroxidase (HRP)-conjugated) were
incubated
for
one
hour
and
signals
were
detected
using
enhanced
electrochemiluminescence (ECL) (Pierce, Rockford, IL, USA).
3.3.7 Microscopy
The immunostained neurons were examined using an Axioplan 2 microscope with UVepifluorescence (Zeiss, Göttingen, Germany). Images were photographed with a digital
camera (AxioCam MRm, Zeiss) and documented using the Axiovision 4.5 software
(Zeiss). Images of neurons immunostained for the detection of synaptic puncta were
examined using the confocal laser-scanning microscope LSM 510 meta (Zeiss). Gain
and threshold were not changed during the experiments. Z-stacks were taken with a
- 56 -
Chapter 3
0.25 µm interval and an overlay of the resulting single pictures was performed
afterwards.
3.3.8 Quantifications and statistical analyses
Quantification of the intensity of the emerged protein bands was carried out using
ImageJ (http://rsbweb.nih.gov/ij/) and results were normalized to the appropriate β III
tubulin bands.
For the detection of immunopositive and PNN bearing neurons, cells were randomly
chosen under the microscope and the percentage of immunopositive neurons from 100
Hoechst positive cells was counted.
The intensity of the PNN staining was quantified using a macro for pixel detection in
ImageJ. Synaptic puncta were counted using the puncta-analyzer plug-in from Barry
Wark (licensed under http://www.gnu.org/copyleft)) for ImageJ. We verified that this
semi-automatic analysis led to results that are similar to those we have obtained by
visual counting.
The Data is given in mean ± standard error of the mean (sem). We performed ANOVA
test with a significance level set to p ≤ 0.05. Following pairwise comparisons were done
with Students´ T-test, while Scheffé post test was used for multiple comparisons.
- 57 -
Chapter 3
3.4 Results
Primary hippocampal neurons were cultivated in indirect co-culture with astrocytes,
allowing for the molecular exchange between both cell types via the defined medium.
Thereby, we focused on the contribution of extracellular matrix molecules in neuronal
development, synapse- and PNN formation, using a quadruple matrix knock-out mouse
(Rauch et al., 2005).
3.4.1 Primary quadruple knock-out neurons and the extracellular
matrix expression pattern
We used primary hippocampal neurons and cortical astrocytes from the quadruple
knock-out mouse lacking the four matrix molecules tenascin-C, tenascin-R, neurocan
and brevican (Rauch et al., 2005). Neurons and astrocytes were combined with wildtype cells in the four possible combinations (see Fig. 1a). Neurons and Glia shared the
same defined medium, but had no direct, membrane-mediated contact, as indicated in
the scheme in Fig. 1a. Growing in this set-up, neurons developed complex networks and
survived for at least 21 DIV (Fig. 1 b-e). Neurons derived from mutant tissue appeared
morphologically normal on the first glimpse and via immunocytochemical staining
against βIII-tubulin or neurofilament (Fig. 1 b-e) no gross morphological differences
could be detected. Further, the survival and the cells´ viability was not altered (data not
shown). First, we investigated the expression of the four matrix molecules under
control conditions, where both cell types were derived from wild-type tissue. We found
a strong extracellular staining pattern of the antibodies against the four ECM proteins
tenascin-C, tenascin-R, neurocan and brevican (Fig. 1 b) after 14 DIV when neurons and
astrocytes were derived from wild-type tissue. Most of the immunoreactivity was
detectable around the soma and proximal dendrites and in some areas a substrate
staining was visible. The brevican expression pattern tended to be enhanced around one
individual neurite (Fig. 1 b, left lower image). Interestingly, the combination of wild-type
neurons with astrocytes derived from the knock-out cortices (Fig. 1 c) did not result in a
lack of one of the proteins detected in the control cultures.
- 58 -
Chapter 3
Figure1: Neuron-astrocyte Co-culture set up and the expression of extracellular matrix molecules
in vitro
a) Scheme of the indirect neuron-astrocyte co-culture set up. Astrocytes (red asterisk) were grown in a
cell culture insert with a permeable membrane facing the neurons (yellow dots) and sharing the same,
wt/wt
knock-out:
defined medium. The four used neuron-astrocyte (N/A) genotype combinations (wild-type:
ko/ko
) are indicated (1-4). b) - e) Immunofluorescent stainings of neurons co-cultured with astrocytes after
14DIV in the different combinations with antibodies against tenascin-C, tenascin-R, neurocan and
brevican. Scale Bar is 50µm.
Quantification of the staining is given in f), bars represent ± s.e.m., N = 3, n = 300.
- 59 -
Chapter 3
A strong staining of the four molecules around the neurons was detectable, despite the
absence of expression in the astrocyte monolayer, suggesting that neurons themselves
may be capable of matrix expression under particular conditions. Here, we found again
the tendency of brevican to accumulate around one selected process (Fig. 1 c, left
lower panel). In cultures where both cell types were derived from the mutant, no ECM
immunoreactivity whatsoever could be detected, confirming the absence of tenascin-C,
tenascin-R, neurocan and brevican.
In the rescue experiment where matrix-deficient neurons were co-cultivated with wildtype astrocytes, the distinct brevican expression pattern was also visible (Fig. 1d) and
the tenascin-C expression was still present, despite the lack of expression within the
neuronal monolayer. In contrast, we were no longer able to detect tenascin-R and
neurocan in primary knock-out neurons, in spite of the presence of wild-type astrocytes.
Therefore, astrocytes in the indirect co-culture appear to be capable of producing
tenascin-C and brevican, while tenascin-R and neurocan seem to be uniquely produced
in the neuronal layer. The quantification of the percentage of the immunopositive
neurons in the different neuron-astrocyte combinations (Fig. 1 f) revealed that the four
matrix molecules were expressed as follows: Awt/wt|Nwt/wt : tenascin-C: 42 ± 5 %,
tenascin-R: 40 ± 5 %, neurocan: 34 ± 2 %, and brevican accumulated around 35 ± 8 %.
Ako/ko|Nwt/wt: tenascin-C: 38 ± 4 %, tenascin-R: 40 ± 9 %, neurocan: 38 ± 2 %, brevican:
38 ± 6 %. If both cells where from knock-out (Ako/ko|Nko/ko), none of the neurons
expressed one of the mentioned matrix molecules, as indicated above. In knock-out
neurons, which were cultivated with wild-type astrocytes (Awt/wt|Nko/ko), we found 40 ± 2
% of neurons expressing tenascin-C and 44 ± 9 % expressing brevican, while neurocan
and tenascin-R were not detectable. The expression levels of the respective proteins in
the three combinations were not significantly different (ANOVA≥ p0.05). Thus, the
mutations did not result in a changed expression level of one of the other four
investigated matrix molecules in the wild-type cell compartment.
- 60 -
Chapter 3
3.4.2 Reduced frequency of mPSCs in patch clamp recordings
Recent studies have demonstrated that mice deficient in ECM molecules such as
tenascin-R (Bukalo et al., 2001; Saghatelyan et al., 2004) and tenascin-C show an
impairment of long-term potentiation (LTP) (Evers et al., 2002). In order to evaluate
whether the brevican/neurocan/tenascin-C/tenascin-R quadruple knock-out leads to
changes of neuronal physiology, we performed whole-cell patch-clamp recordings in
neurons grown for 14 and 21 DIV and analyzed all knock-out/wild-type combinations of
astrocyte-neuron culture conditions (Fig. 2A).
In the last years, the analysis of postsynaptic events in neurons has become a useful
tool to investigate the mechanisms involved in neurotransmission. In our study, the
miniature postsynaptic currents (mPSCs), inhibitory or excitatory postsynaptic events
that are elicited by the release of a single vesicle of transmitter in the absence of
presynaptic stimuli were recorded. The addition of specific neuroblockers (TTX, DNQX,
and PTX) allowed for the pharmacological isolation of excitatory or inhibitory
postsynaptic currents (mEPSCs or mIPSCs, respectively). We used astrocyte/neuron
cultures grown for 14 days in vitro (DIV 14) in the Awt/wt|Nwt/wt, Awt/wt|Nko/ko, Ako/ko|Nwt/wt,
and Ako/ko|Nko/ko combinations and observed that neither the kinetics of mPSCs, i.e. the
rise and decay time), nor the amplitudes of the recorded currents were significantly
different among groups (Data not shown).
Interestingly, analyzing the frequency of mPSC events, we found that neurons grown in
Ako/ko|Nko/ko combination showed a strongly reduced mEPSC frequency (0.06 ± 0.006
Hz, n = 13 neurons, p= 0.02), when compared with control cultures (0.2 ± 0.031 Hz, n =
15 neurons) (Fig. 2C). Similarly to the mEPSCs, we could not detect any differences in
kinetics or amplitude of mIPSCs recorded from neurons grown the various ko/wt
combinations (Fig. 2B) after 14 DIV. But, when analyzing the mIPSC frequency, we
observed that neurons grown in Ako/ko|Nko/ko (0.08 ± 0.002 Hz, n = 12 neurons, p= 0.02),
and also in Awt/wt|Nko/ko (0.1 ± 0.005 Hz, n = 30 neurons, p = 0.03) cultures showed a
significantly decreased frequency when compared to control neurons Awt/wt|Nwt/wt (0.2 ±
0.05 Hz, n = 14 neurons) (Fig. 2C).
- 61 -
Chapter 3
Figure 2: Whole-cell voltage-clamp recordings of miniature inhibitory and excitatory postsynaptic
currents (mIPSCs, mEPSCs) in the four astrocyte-neuron culture combinations after 14 and 21 days
in vitro (DIV). a) Primary hippocampal neuron and patch-clamp pipette recorded in a control culture after
14 DIV. Neurons were voltage-clamped at -60 mV. b) mIPSCs traces recorded from wild-type
wt/wt
wt/wt
ko/ko
ko/ko
|N
) and knock-out (A
|N
) cell culture combinations. For the pharmacological isolation,
(A
TTX (1µM) and DNQX (10 µM) were added to the bath solution. c) Mean frequencies of mIPSCs and
wt/wt
ko/ko
ko/ko
ko/ko
|N
and A
|N
culture
mEPSCs after 14 DIV were significantly decreased in the A
wt/wt
ko/ko
ko/ko
ko/ko
|N
and A
|N
combinations. d) After 21 DIV, mIPSC and mEPSC mean frequencies of the A
combinations showed a significant reduction when compared to control cultures. Data are represented
as mean ± s.e.m. and were considered significantly different at p≤ 0.05 in ANOVA test. The respective n
is indicated in brackets.
- 62 -
Chapter 3
Frischknecht and co-workers showed that the ECM matured at the end of the second
and beginning of the third week in culture, when brevican and hyaluronan were present
on neuronal surfaces (Frischknecht, Heine et al. 2009).
Figure S1: Whole-cell voltage-clamp recordings of miniature inhibitory and exictatory postsynaptic
currents (mIPSCs, mEPSCs) in the neuron-astrocyte cell combinations after 13 and 21 days in vitro
(DIV). a) mIPSCs and mEPScs mean amplitudes of knock-out cell combinations after 13 DIV were not
significantly affected when compared to wild type cells b) mIPSCs and mEPScs mean amplitudes of
knockout cell combinations after 21 DIV were not significantly affected when compared to wild type
cells. Data are represented as mean ± s.e.m. and were considered significantly different at p< 0.05 in
ANOVA test. The respective n is indicated in brackets.
This special feature, together with the fact that our cells remained stable after 21 DIV,
allowed us to perform further electrophysiological recordings. Following the same
standard whole-cell patch-clamp protocols we repeated our experiments with cultures
grown for 21 DIV. Similar to the results obtained in DIV 14 cultures, mPSC kinetics and
amplitude were not different between groups, but the frequency of mEPSCs were
affected: neurons grown in Ako/ko|Nko/ko (0.16 ± 0.04 Hz, n = 12 neurons, p = 0.02) but
- 63 -
Chapter 3
also in the Awt/wt|Nko/ko combination (0.2 ± 0.04, n = 15 neurons, p = 0.02) showed
reduced mEPSC frequencies compared to Awt/wt|Nwt/wt control cultures (0.4 ± 0.08
Hz, n = 19 neurons) (Fig. 2D). Again, mIPSC frequency of neurons recorded in the
Awt/wt|Nko/ko (0.06 ± 0.006 Hz, n = 15 neurons, p = 0.01) and Ako/ko|Nko/ko (0.04 ±
0.002 Hz, n= 13 neurons, p = 0.01) combinations showed a significant reduction
compared to control cultures (Fig. 2D).
Our data show that the frequency of mPSCs is a functional parameter affected in
neurons obtained from animals with a mutation of ECM components, while other
parameters were not affected.
3.4.3 Synapse formation in the indirect neuron-astrocyte co-culture
assay
To investigate whether the electrophysiological phenotype of a significantly decreased
mIPSCs and mEPSCs frequency in the knock-out neurons (see Fig. 2) was paralleled
with a reduced expression of synaptic proteins, we performed immunocytochemical
stainings, followed by semi-automatic read-out of synaptic puncta expression (Ippolito
and Eroglu, 2010). Therefore, immunodetection of Bassoon and PSD 95 in the four
different combinations of mutant and wild-type neurons after 14 and after 21 DIV was
performed (Fig. 3). Primary hippocampal neurons, grown in indirect co-culture with
astrocytes expressed synaptic proteins and built structural synapses after 7 days in
culture. The immunological detection of the two synaptic proteins revealed a punctate
staining pattern (Fig. 3a, c and higher magnification). Beside the detection of Bassoon
and PSD 95positive puncta, we could observe overlapping dots, indicating the colocalization of both proteins and referring to a structurally intact synapse. In the matrix
deficient combinations, we recognized a tendency towards an enhanced synaptic
puncta expression compared to the control situation (Awt/wt|Nwt/wt), after 14 DIV, while
comparing the mean percentages of in- and decrease in synaptic puncta expression
between the different conditions.
- 64 -
Chapter 3
Figure 3: Synaptic puncta expression in primary hippocampal neuron-astrocyte co-cultures of
different genotype combinations
Immuncytochemical stainings of primary hippocampal neurons with antibodies against Bassoon and PSD
95. Neurons were grown for 14 DIV (a) and 21 DIV (c) in the four different neuron-astrocyte combinations
as indicated (A: genotype astrocytes, N: genotype neurons). Areas of higher magnifications are indicated
in the red boxes and are shown in adjacent images. Scale Bar is 20 µm. Quantification of the punctate
staining (red puncta: bassoon, green puncta: PSD 95, yellow puncta: co-localization of both) after 14 (b)
and 21 DIV (d). Data represent mean values of percent de- and increase compared to the control
wt/wt
wt/wt
|N
) and error bars present ± s.e.m. Data were considered significantly different at p≤
situation (A
0.05 in ANOVA and Scheffe post hoc test. N = 3, n = 120.
- 65 -
Chapter 3
The following values were measured: In the Awt/wt|Nko/ko combination we found an
increased expression of Bassoon of about 16 ± 6 %, the PSD 95 expression was
increased about 6 ± 6 % and an enhanced co-localization of both proteins of about 12 ±
6 % compared to the control situation could be detected (ANOVA p > 0.05). A more
attenuated, yet not significant (ANOVA p > 0.05) increase in synaptic puncta expression
was observed in control neurons combined with mutant astrocytes (Ako/ko|Nwt/wt
Bassoon: 49 ± 7 %, PSD95: 30 ± 7 %, Co-localization: 48 ± 7 %). The same attenuated
increase was detectable in the combination where both cell types where derived from
the knock-out tissue (Ako/ko|Nko/ko, Bassoon: 32 ± 8 %, PSD95: 22 ± 8 %, Co-localization:
33 ± 8 %). This increased synaptic puncta expression was significantly detectable in
most of the single experiments (ANOVA p ≤ 0.05). The immuncytochemical stainings of
more mature cultures after 21 DIV are shown in Fig. 3 c. In all four combinations, the
total expression of synaptic proteins increased further with time in culture,
corroborating a proper development of all cultures.
However, we found a significant (ANOVA ≤p 0.05, Scheffé post hoc test p = 0.017)
decrease in the relative expression of PSD 95 to about -35 ± 7 % when both cell types
were depleted of the four matrix molecules (Ako/ko|Nko/ko) compared to the wild-type
situation (Awt/wt|N wt/wt). Furthermore, the co-localization of PSD 95 and Bassoon was
significantly (ANOVA p ≤ 0.05, Scheffé post hoc test p = 0.018) decreased by about -33
± 7 % in the Ako/ko|Nko/ko combination compared to the control cells. This decrease could
also be observed when primary neurons from the mutant mice where cultivated with
wild-type
astrocytes (Awt/wt|Nko/ko, Bassoon: -17 ± 7 %, PSD 95: -23 ± 7 %, Co-
localization: -20 ± 7 %) and the other way around (Ako/ko|Nwt/wt Bassoon: -16 ± 6 %, PSD
95: -19 ± 6 %, Co-localization: -16 ± 7 %), but the differences compared to the wild-type
combination (Awt/wt|Nwt/wt) where in both cases not significant (ANOVA p > 0.05).
Nevertheless, we found a significantly decreased total expression of synaptic puncta on
the single experiment level when knock-out and wild-type neurons were compared
(ANOVA p ≤ 0.05).
In summary, the four matrix molecules seem to exert a different impact in early and
late stages of synapse formation in vitro, not directly reflecting the physiological
phenotype.
- 66 -
Chapter 3
3.4.4 Quantitative protein analysis of GAD 65, GAD 67 and vGlut
The expression pattern of synaptic proteins after 21 DIV could directly reflect the
decreased frequency of mEPSCs, while after 14 DIV an indirect, secondary functional
deficit may be causing the observed electrophysiological phenotype. To verify whether
there is a change in the inhibitory and excitatory neuronal subpopulations, we checked
for the expression of the two Glutamate-decarboxylase (GAD) isoforms 65 and 67 (GAD
65/67) and the vesicular Glutamate transporter (vGlut) after 14 and after 21 DIV in the
four different neuron-astrocyte combinations (Fig. 4). Both proteins were detectable in
the indirect co-culture. The GAD 65/67 expression relative to βIII tubulin increased in all
four combinations from 14 to 21 DIV (Awt/wt|Nwt/wt 14 DIV: 0.36 ± 0.17, 21 DIV: 0.78 ±
0.53; Awt/wt|Nko/ko: 14 DIV: 1.15 ± 0.25, 21 DIV: 1.88 ± 0.22; Ako/ko|Nwt/wt14 DIV: 0.33 ±
0.12, 21 DIV: 0.51 ± 0.39; Ako/ko|Nko/ko 14 DIV: 0.88 ± 0.08, 21 DIV: 1.62 ± 0.64, Fig. 4a
and b), emphasizing again the vitality and the proper development of the neuronal
network. However, we detected significant (ANOVA p≤ 0.05) differences between the
four combinations and found a significantly increased GAD 65/67 expression after 14
DIV, when neurons were derived from the mutant, independently of the genotype of the
astrocyte layer added (Awt/wt|Nwt/wt 0.36 ± 0.17 vs. Awt/wt|Nko/ko: 1.15 ± 0.25; post hoc
test: p = 0.033 and (Ako/ko|Nwt/wt vs. Awt/wt|Nko/ko; post hoc test: p = 0.024, Fig. 4b). The
neurons grown in the Ako/ko|Nko/ko combination showed also an increased GAD 65/67
expression compared to the wild-type neurons (post hoc test: Ako/ko|Nko/ko 0.88 ± 0.08
vs. Awt/wt|Nwt/wt 0.36 ± 0.17; post hoc test: p = 0.234). After 21 DIV, there were still
strong differences in the GAD65/67 expression between the wild-type and the knockneurons detectable: Awt/wt|Nwt/wt: 0.78 ± 0.53; Awt/wt|Nko/ko: 1.15 ± 0.25; Ako/ko|Nko/ko 1.62
± 0.64, ANOVA p = 0.270, Fig. 4b. Overall, the mutant neurons displayed an increased
GAD65/67 expression compared to the wild-type neurons, independent of the added
astrocytes. The expression of the vesicular glutamate transporter vGlut was detectable
in all four cultured neuron-astrocyte combinations (Fig. 4 c). Overall, we found a strong
vGlut expression, that increased further with increasing time in culture (Awt/wt|Nwt/wt14
DIV: 1.04 ± 0.3, 21 DIV: 1.2 ± 0.5; Awt/wt|Nko/ko 14 DIV: 1.38 ± 0.33, 21 DIV: 2.05 ± 0.82;
Ako/ko|Nwt/wt14 DIV: 1.18 ± 0.2, 21 DIV: 1.64 ± 0.83; Ako/ko|Nko/ko 14 DIV: 1.39 ± 0.15, 21
DIV: 1.62 ± 0.59, Fig. 4c and d). Close inspection revealed, however, that the vGlut
- 67 -
Chapter 3
expression was not modified by the matrix removal in the mutants (ANOVA 14 DIV and
21 DIV p > 0.05). Thus, despite the changed synapse formation and the decreased
mEPSC frequency, the vGlut expression was unaffected.
Figure 4: Quantitative expression levels of vGlut and GAD 65 and 67 in primary hippocampal
neuron-astrocyte co-cultures
a) Immunodetection of GAD 65, GAD 67 and βIII-tubulin in Western Blots with protein lysates, derived
from neurons co-cultured with astrocytes for 14 and 21 DIV. b) Quantifications of the GAD 65 and 67
expressions were normalized to βIII-tubulin bands. c) Immunodetection of vGlut and βIII-tubulin after 14
and 21 DIV in the same culture. Quantification of the relative vGlut expression is shown in d). Different
genotypes are indicated as follows: A: genotype of astrocytes, N: genotype of neurons. Data represent
means ± s.e.m. and were considered significantly different at p≤ 0.05 in ANOVA and Scheffe post hoc
test. 14 DIV n=7, 21 DIV n=3.
- 68 -
Chapter 3
3.4.5 PNN formation in primary hippocampal neurons lacking four
matrix components
A subpopulation of neurons is known to recruit a defined set of matrix molecules to
form perineuronal nets (PNNs) (Celio et al., 1998). PNNs are a specialized form of the
extracellular matrix, which accumulate around maturing neurons. The restricted matrix
expression in the investigated quadruple knock-out mouse could affect the PNN
formation and may contribute secondarily to the observed changes of synaptic
physiology and morphology. For this reason, we investigated the PNN formation and
maintenance in the available in vitro assay.
Initial formation of PNNs around a small subpopulation of neurons in the indirect
neuron-astrocyte co-culture was recognized around 10 DIV by WFA (Wisteria floribunda
Agglutinin) staining (data not shown). The lattice-like matrix spanned the neuronal cell
body and apical dendrites, with an increasing extent and complexity over time in
culture. We quantified the formation of PNNs in the four different combinations after 14
and 21 DIV (Fig. 5 a, b) with WFA labeling. In the Awt/wt|Nwt/wt combination a bright WFA
fluorescent signal appeared around 12 ± 1 % of βIII-tubulin positive neurons after 14
DIV (Fig. 5a, c), and this picture was maintained for the following time in vitro ( 21 DIV:
11 ± 1 % of neurons). Despite the partial lack of matrix in the glia cell compartment, we
observed 10 ± 2 % of βIII-tubulin positive neurons surrounded by PNNs both after 14
DIV as well as after 21 DIV (Fig. 5c) in the Ako/ko|Nwt/wt culture. Thus, neurons were
capable of proper net formation, despite the lack of four matrix molecules in the
astrocyte compartment. The immuno-detection of PNNs after 14 DIV revealed a
remarkable (ANOVA p ≤ 0.05) different picture in the knock-out neurons: only 5 ± 1 % of
βIII-tubulin positive neurons were decorated by a WFA-positive staining pattern in the
Ako/ko|Nko/ko combination (post hoc test p = 0.019) and interestingly, this could not be
rescued by the addition of wild-type astrocytes (Awt/wt|Nko/ko, 4 ± 1 %, p = 0.009). After
21 DIV, the fraction of PNN wearing neurons further decreased significantly (ANOVA p ≤
0.05) to 2 ± 0 % (p = 0.05) in the Ako/ko|Nko/ko combination and to 3 ± 0 % in the
Awt/wt|Nko/ko situation (p = 0.06), compared to the Awt/wt|Nwt/wtcombination.
- 69 -
Chapter 3
Figure 5: Perineuronal net formation in vitro
Detection of PNNs, accumulated around βIII-tubulin positive neurons, after 14 (a) and 21 DIV (b) via
Wisteria Floribunda Agglutinin (WFA) binding. Primary hippocampal neurons from different genotypes
were co-cultured with respective astrocytes as indicated (A: genotype astrocytes, N: genotype neurons).
Scale Bar is 50 µm. The percentage of PNN bearing neurons was quantified and is shown in c) N=3,
n=300. The complexity of the nets was quantified via respective pixel counting and is shown in d), N= 3.
Data is represented as mean values and was considered as significantly different at p ≤ 0.05 in ANOVA
and Scheffe post hoc tests. Error bars indicate ± s.e.m.
Secondary to the decreased percentage of neurons that carried PNNs, we could
demonstrate a significant (ANOVA p≤ 0.05) reduction of the net complexity around
knock-out neurons (Fig. 5 a, b, d). Via pixel quantification of the WFA staining, we could
demonstrate that the PNNs increased in complexity in the Awt/wt|Nwt/wt assay and that
- 70 -
Chapter 3
the matrix network accumulated over a larger surface with increasing time in culture
(Awt/wt|Nwt/wt14 DIV: 10785 ± 1332 pixel; 21 DIV: 21117 ± 7558 pixel). In the co-culture
combination devoid of the four matrix molecules (Ako/ko|Nko/ko), this complexity was
significantly decreased and vanished further after 21 DIV (14 DIV: 3301 ± 680 pixel; p =
0.009; 21 DIV: 662 ± 197 pixel, p = 0.001). This phenotype could not be rescued by the
addition of wild-type astrocytes (Awt/wt|Nko/ko), where a significantly decreased
complexity after 14 (2834 ± 435 pixel, p = 0.002) and after 21 DIV (765 ± 435 pixel, p =
0.001) was again detectable. Interestingly, wild-type neurons co-cultured with knockout astrocytes showed a reduced elevation of the PNN complexity from 14 (8567 ±
1822 pixel) to 21 DIV (10281 ± 3691 pixel) in comparison to the Awt/wt|Nwt/wt
combination (p = 0.212), suggesting also an astrocytic contribution in PNN development
and maintenance.
In summary, the knock-out of tenascin-C, tenascin-R, brevican and neurocan resulted in
a strongly compromised PNN formation around primary hippocampal neurons, which
could not be rescued by the neighboring wild-type astrocytes.
3.4.6 Synapse formation on PNN wearing neurons
The reported results suggested that primary hippocampal neurons lacking four matrix
components a characterized by a failure of PNN formation and maintenance.
Furthermore, we could demonstrate that the expression of synaptic proteins is involved
in the phenotype of the quadruple matrix mutation. Thus, we decided to investigate the
synapse
formation
focusing
on
PNN-coated
neurons
and
performed
triple
immunocytochemical stainings with WFA and antibodies against Bassoon and PSD 95
(Fig. 6). This approach resulted in appropriate fluorescent stainings, where the
punctuate expression patterns of the synaptic proteins became visible directly flanked
by the WFA fluorescence. Interestingly, the appearance of synaptic proteins correlated
directly with the gaps in the lattice-like WFA staining (Fig. 6a, b). This was further
demonstrated in the color profile, where the fluorescent spectra of a representative
neurite are shown (Fig. 6b). The immunoreactivity of Bassoon and PSD 95 appeared
clearly framed by the PNNs. To investigate modifications in the expression of synaptic
- 71 -
Chapter 3
proteins on the PNN coated subpopulation of neurons we quantified the punctate
staining and the co-localization of both synaptic proteins via semi-automatic read-out
with ImageJ (Ippolito and Eroglu, 2010). Here we found, in contrast to the results
obtained with the randomly picked neurons, a significant reduction in synaptic protein
expression after 14 (ANOVA p ≤ 0.05) and after 21 DIV (ANOVA p ≤ 0.05), when neurons
originated from knock-out hippocampal tissue. The mean percentage of the de/increase in synaptic puncta expression after 14DIV was significantly reduced for
Bassoon (-46 ± 11%, p = 0.005), PSD 95 (-54 ± 11%, p = 0.009) as well as for the colocalization of both protein (-55 ± 13%, p = 0.026) when knock-out neurons were
combined with knock-out astrocytes (Ako/ko|Nko/ko). This effect could not be rescued by
the co-cultivation of astrocytes originated from wild-type mice (Awt/wt|Nko/ko) and we
found a significant decrease in synaptic puncta expression in that combination
(Bassoon: -51 ± 13%, p = 0.003, PSD 95: -45 ± 12%, p = 0.026, Co-localization: -57 ±
13%, p = 0.022). In contrast, the lack of the four matrix molecules in the astrocyte
compartment did neither result in a significantly decreased expression nor in decreased
co-localization of synaptic protein in wild-type neurons (Bassoon: 0 ± 7%, p = 1.0, PSD
95: -8 ± 12%, p = 0.923, Co-localization: -9 ± 9%, p = 0.932).
After 21 DIV this effect persisted, and the lack of the four ECM molecules resulted
again in a decreased synaptic protein expression in the Ako/ko|Nko/ko combination,
although the effect was not as pronounced as after 14 DIV (Bassoon: -34 ± 8%, p =
0.088, PSD 95: -51 ± 11%, p = 0.05, Co-localization: -39 ± 10%, p = 0.305). In the rescue
experiment where wild-type astrocytes were added, no salvage of this effect could be
observed and there was still a reduction in synaptic puncta expression (Bassoon: -31 ±
8%, p = 0.139, PSD 95: -43 ± 10%, p = 0.089, Co-localization: -37 ± 10%, p = 0.351),
while the lack of matrix molecules in astrocytes did not result in significant changes in
synapse formation between wild-type neurons (Bassoon: -7 ± 8%, p = 0.940, PSD 95: -8
± 10%, p = 0.958, Co-localization: 7 ± 18%, p = 0.986).
Thus, synapse formation on the subpopulation of PNN wearing neurons was severely
affected by the matrix knock-out. That could not be rescued by adding wild-type
astrocytes, while the lack of four matrix molecules in the astrocytic compartment did
not result in a changed expression of synaptic proteins in wild-type neurons.
- 72 -
Chapter 3
Figure 6: Synapse formation on PNN coated neurons
Immuncytochemical detection of Bassoon, PSD 95 and PNNs (via WFA binding) in cultures of primary
hippocampal neurons. a) Triple staining of Bassoon, PSD95 and WFA, where synaptic puncta emerge in
the gaps between WFA positive areas. This is precise and adjacent expression is underlined by the color
profile of an exemplary neurite, shown in b). c) and d) Triple staining of primary hippocampal neurons of
different genotypes (N) grown in indirect co-culture with respective astrocytes (A) after 14 (c) and 21 DIV
(d). Higher magnifications from exemplary neurites are shown in smaller images and the area is outlined
in white boxes. Scale Bar is 10 µm. e) Quantification of the change in synaptic puncta expression
wt/wt
wt/wt
|N
) after 14 DIV and after 21 DIV (f). Data is represented as mean values
compared to controls (A
and was considered as significantly different at p≤ 0.05 in ANOVA and Scheffe post hoc test. Error bars
indicate ± s.e.m.
- 73 -
Chapter 3
3.5 Discussion
The current in vitro study presents a detailed functional investigation of hippocampal
neurons lacking the four extracellular matrix molecules tenascin-C, tenascin-R,
neurocan and brevican. We show that mutant neurons exhibit severe deficits in PNNand synapse formation, paralleled with changes in their electrophysiological properties.
Moreover, we provide evidence for expression level changes of key synaptic proteins.
The combinatorial cultivation of both wild type and mutant neurons as well as
astrocytes revealed the contribution of both cell types to the matrix assembly.
Hippocampal neurons were able to produce tenascin-C, tenascin-R, brevican and
neurocan, as reported previously (Engel et al., 1996; Ferhat et al., 1996a; Ferhat et al.,
1996b; Fuss et al., 1993; Lander et al., 1998; Pyka et al., 2011b; Wintergerst et al.,
1993; Zhang et al., 1995), while astrocytes contributed to the release of brevican and
tenascin-C (see Fig.1) (Bartsch et al., 1992; Cahoy et al., 2008; Karus et al., 2011). The
distinct accumulation of brevican observed around one selected neurite (see Fig. 1)
nicely fits to the work from Seidenbecher and colleagues, who reported the localization
of Brevican at the axon hillock (Hedstrom et al., 2007; John et al., 2006).
Via whole-cell voltage-clamp recordings we observed a significantly reduced frequency
of mIPSCs and mEPSCs when neurons were deficient of the four matrix molecules (see
Fig. 2). Interestingly, the observed reduction was independent from the genotype of the
astrocytes, and could not be rescued by adding wild-type astrocytes (see Fig. 2).
Different changes in the synaptic connectivity in relation to matrix proteins have been
described previously (Bukalo et al., 2007, 2001; Evers et al., 2002; Irintchev et al., 2005;
Morellini et al., 2010). Thus, hyaluronan and tenascin-C are involved in LTP induction,
via acting on L-type voltage-dependent Ca2+channels (Evers et al., 2002;
Kochlamazashvili et al., 2010). Recently, it has been shown that AMPA receptors´
lateral mobility is enhanced after enzymatic removal of hyaluronan (Frischknecht et al.,
2009) and the ECM is considered as a diffusion barrier for transmitter molecules and
other substances acting within the synaptic cleft (Dityatev et al., 2010; Gundelfinger et
al., 2010). Therefore, ECM molecules seem to influence the synaptic machinery at
different points of action. Pyka and colleagues reported a decreased mEPSC amplitude
- 74 -
Chapter 3
and charge after ChondroitinaseABC treatment, while the mEPSC frequency was
unchanged, and mIPSCs were not affected at all (Pyka et al., 2011b). Surprisingly, both
parameters were shown to be unchanged in the mutant in vitro (see Fig. S1). However,
the mutation of the four matrix molecules points to a rather presynaptic effect,
underlining the fundamental differences of the two models (enzymatic treatment vs.
genetic matrix manipulation). Changes in the mPSC frequency can be explained by
changes in the number of functional release sites (Hsia et al., 1998). Further, the pool of
ready-to-release vesicles and the number of presynaptic inputs affects the frequency of
such events. Further, changes in the neuronal populations can not be excluded. To
obtain a first idea about the neuronal subpopulations, we performed western blot
analysis for the most characteristic population proteins GAD 65/67 and vGlut.
Surprisingly, we found a significantly increased GAD65/67 expression in mutant
neurons regardless of the astrocytes while the vGlut levels were unaltered (see Fig. 3).
Different studies describe a shift in the GABAergic population towards more GABAergic
neurons in the hippocampus of TNR -/- mice (Morellini et al., 2010), while in the cortex
of TNC-/- mice a lower density of parvalbumin-positive interneurons was observed
(Irintchev et al., 2005). However, the increased GAD65/67 expression seemed to have
no direct one-to-one functional outcome, as the mIPSC frequency was decreased and
the amplitude of mIPSCs was not altered compared to the wild-type. At this point, we
can not decide whether the matrix mutation led to an increase in the portion of GAD
expressing neurons or to a higher expression of GAD in a given number of cells.
Nevertheless, despite the increased GAD expression, the GABA release mechanism
seemed to be altered. The increased amount of GAD65/67 may have led to the
enhanced formation of silent, but not of active GABAergic synapses, explaining the
decreased mIPSC frequency in the mutant. Evidence for the involvement of the ECM in
activating silent synapses comes from previous experiments, and interestingly sugar
binding proteins, such as neuronal pentaxin (Cho et al., 2008) and integrin ECM
receptors (Milner and Campbell, 2002) seem to be highly involved in these processes.
Despite the observed changes in the frequency of mEPSCs, no alterations in the vGLUT
expression could be demonstrated (see Fig. 3). We could not determine, however,
- 75 -
Chapter 3
whether this was due to a reduced glutamatergic neuron population with concomitantly
higher vGLUT expression.
A further hint to the involvement of the ECM in synaptic plasticity and a mechanistic
explanation for the aforementioned changes in the frequency of postsynaptic miniature
currents was obtained by quantifying the expression of synaptic proteins in this assay.
Pyka and colleagues demonstrated that the enzymatic treatment with ChABC resulted
in a significantly increased expression of synaptic proteins in rat hippocampal cultures
(Pyka et al., 2011b). In line with that we found an enhanced expression of Bassoon and
PSD95 in matrix deficient neurons after 14 DIV (see Fig. 4). Further, these neurons
displayed an enhanced co-localization of both proteins, indicating the occurrence of a
structural synapse. Whether intact functionality of these synapses was given could not
be concluded, and a structural but not functional synaptic protein assembly would fit to
the observed decreased frequency. This increase in synaptic proteins formed after 14
DIV fits to the idea, that the matrix plays a repellent and restrictive role, which was
demonstrated in several elegant studies in the context of regeneration (Garcia-Alias
and Fawcett, 2011; Wang et al., 2011). Thus, we propose that the lack of matrix
molecules resulted in an altered synapse formation during early development.
Interestingly, the effect was pronounced when astrocytes were also deficient of the
four matrix molecules, underlining the glial contribution. To gain insight into the role of
the ECM in more mature networks, we further investigated the expression of synaptic
proteins after 21 DIV. The total expression of synaptic proteins increased from 14 to 21
DIV in each condition, indicating the proper development of the networks, similar to the
increased GAD and vGlut expression and the increases in frequency and amplitude
during that developmental switch from 14 to 21 DIV. Yet, the expression of synaptic
proteins was significantly decreased in cultures deficient of the four matrix molecules,
particular when both cell types lacked the four components compared to the wild type
condition. The ECM plays a multifarious role during development. Beside the restriction
and control of synapse formation (Pyka et al., 2011b) , CSPGs, especially in the flavor of
PNNs, tightly enwrapping neurons, are thought to maintain synapse integrity (for review
see Dansie and Ethell, 2011). Thus, the lack in matrix molecules stabilizing the
synapses built, may explain the observed changes. PNNs are an outstanding example
- 76 -
Chapter 3
for the involvement of the ECM in the synaptic machinery. PNNs are characteristic
accumulations of matrix molecules around a subpopulation of mature neurons (Celio et
al., 1998). In the indirect hippocampal co-cultures, initial PNN formation was observed
around 7 DIV, with increasing complexity emerging over time in vitro. During the last
year, evidence accumulated that aggrecan and Crtl1 are crucial for the initiation of PNN
formation (Giamanco et al., 2010; Kwok et al., 2010). Previous studies have
demonstrated that tenascins and the lecticans are expressed in PNNs (Carulli et al.,
2006; Deepa et al., 2006; Matsui et al., 1998). In line with this, an altered PNN formation
in vivo was previously described for the Tenascin-R mutant (Bruckner et al., 2000), and
the quadruple mouse showed altered WFA staining pattern in vivo (Rauch et al., 2005).
In line with that, we found a significantly reduced formation of PNN around neurons
from the quadruple knock-out mouse (see Fig. 5). This could not be rescued by adding
wild-type astrocytes. Interestingly, although the lack of astrocyte derived matrix
molecules did not lead to a reduced number of PNN wearing wild-type neurons, the
complexity of the PNNs was severely affected. Thus, it appears that astrocytes at least
in part contributed to the PNN formation. In contrast, neuronally delivered matrix
appeared to be sufficient to initiate the formation. The coincidence of the PNN
formation and the closure of the critical period have led to the idea, that the removal of
PNNs may restore the plastic capacity of the brain (Balmer et al., 2009; Gogolla et al.,
2009). Whether the diminished PNN formation and the altered ECM expression also led
to enhanced plasticity in our assay remains a question for further investigations. On a
behavioral level it was shown that the TNR mutant exhibits faster reversal learning,
improved working memory, and enhanced reactivity to novelty compared to wild-type
littermates (Morellini et al., 2010). The importance of PNNs in synapse formation and
stabilization was clearly demonstrated in the triple stainings, which revealed that
neurons, bearing rudimentary PNNs show significantly reduced synapse formation after
14 and 21 DIV (see Fig. 6).
The neuronal activity, the PNN expression and the synapse formation were altered in
our assay. Accordingly, the subsequent lack in synapse stabilization after 21 DIV is
accompanied by a decreased activity and both phenomena (reduced matrix expression
and reduced activity) may have contributed to the reduced expression of synaptic
- 77 -
Chapter 3
proteins in the matrix deficient cultures. Thus, we propose a dual role of the brain´s
ECM in our assay: While the early matrix is needed to confine synapse formation to a
homeostatic level, the matrix at later stages is needed to stabilize the synaptic contact.
Conclusion
Several recent studies using ChABC demonstrated the important role of the neuronal
ECM for plasticity in vivo and in vitro. However, these studies focus on the impact of the
growth inhibitory GAG chains in a regenerative context. Here, we used a mouse model,
which lacks four major components of the ECM and show that a lack of the core
proteins led to significant changes during neuronal development.
In summary, the quadruple mutant represents a new model system to study neuronal
development in general and synaptic plasticity in particular.
3.6 Acknowledgements
This work was supported by the Research Department of Neuroscience of the Ruhr
University Bochum (http://www.rd.ruhr-uni-bochum.de/neuro), the DFG GRK 396, the
research school of the Ruhr University Bochum (GSC98/1) and the priority programme
SSP 172 “Glia and Synapse” of the German research foundation (DFG).
- 78 -
Chapter 3
3.7 References
Balmer, T. S., V. M. Carels, et al. (2009). "Modulation of perineuronal nets and parvalbumin with
developmental song learning." J Neurosci 29(41): 12878-12885.
Bartsch, S., U. Bartsch, et al. (1992). "Expression of tenascin in the developing and adult
cerebellar cortex." J Neurosci 12(3): 736-749.
Beattie, E. C., D. Stellwagen, et al. (2002). "Control of synaptic strength by glial TNFalpha."
Science 295(5563): 2282-2285.
Boehler, M. D., B. C. Wheeler, et al. (2007). "Added astroglia promote greater synapse density
and higher activity in neuronal networks." Neuron Glia Biol 3: 127-140.
Brakebusch, C., C. I. Seidenbecher, et al. (2002). "Brevican-deficient mice display impaired
hippocampal CA1 long-term potentiation but show no obvious deficits in learning and
memory." Mol Cell Biol 22(21): 7417-7427.
Bruckner, G., J. Grosche, et al. (2000). "Postnatal development of perineuronal nets in wild-type
mice and in a mutant deficient in tenascin-R." J Comp Neurol 428(4): 616-629.
Bukalo, O., M. Schachner, et al. (2001). "Modification of extracellular matrix by enzymatic
removal of chondroitin sulfate and by lack of tenascin-R differentially affects several
forms of synaptic plasticity in the hippocampus." Neuroscience 104(2): 359-369.
Bukalo, O., M. Schachner, et al. (2007). "Hippocampal metaplasticity induced by deficiency in
the extracellular matrix glycoprotein tenascin-R." J Neurosci 27(22): 6019-6028.
Cahoy, J. D., B. Emery, et al. (2008). "A transcriptome database for astrocytes, neurons, and
oligodendrocytes: a new resource for understanding brain development and function." J
Neurosci 28(1): 264-278.
Carulli, D., T. Laabs, et al. (2005). "Chondroitin sulfate proteoglycans in neural development and
regeneration." Curr Opin Neurobiol 15(1): 116-120.
Carulli, D., T. Pizzorusso, et al. (2010). "Animals lacking link protein have attenuated
perineuronal nets and persistent plasticity." Brain 133(Pt 8): 2331-2347.
Carulli, D., K. E. Rhodes, et al. (2006). "Composition of perineuronal nets in the adult rat
cerebellum and the cellular origin of their components." J Comp Neurol 494(4): 559577.
- 79 -
Chapter 3
Celio, M. R., R. Spreafico, et al. (1998). "Perineuronal nets: past and present." Trends Neurosci
21(12): 510-515.
Cho, R. W., J. M. Park, et al. (2008). "mGluR1/5-dependent long-term depression requires the
regulated ectodomain cleavage of neuronal pentraxin NPR by TACE." Neuron 57(6):
858-871.
Christopherson, K. S., E. M. Ullian, et al. (2005). "Thrombospondins are astrocyte-secreted
proteins that promote CNS synaptogenesis." Cell 120(3): 421-433.
Cybulska-Klosowicz, A., R. Zakrzewska, et al. (2004). "Reduced plasticity of cortical whisker
representation in adult tenascin-C-deficient mice after vibrissectomy." Eur J Neurosci
20(6): 1538-1544.
Dansie, L. E. and I. M. Ethell (2011). "Casting a net on dendritic spines: The extracellular matrix
and its receptors." Dev Neurobiol 71(11): 956-981.
Deepa, S. S., D. Carulli, et al. (2006). "Composition of perineuronal net extracellular matrix in rat
brain: a different disaccharide composition for the net-associated proteoglycans." J Biol
Chem 281(26): 17789-17800.
Di Castro, M. A., J. Chuquet, et al. (2011). "Local Ca(2+) detection and modulation of synaptic
release by astrocytes." Nat Neurosci 14(10): 1276-1284.
Dityatev, A., G. Bruckner, et al. (2007). "Activity-dependent formation and functions of
chondroitin sulfate-rich extracellular matrix of perineuronal nets." Dev Neurobiol 67(5):
570-588.
Dityatev, A., C. I. Seidenbecher, et al. (2010). "Compartmentalization from the outside: the
extracellular matrix and functional microdomains in the brain." Trends Neurosci 33(11):
503-512.
Elmariah, S. B., E. G. Hughes, et al. (2005). "Neurotrophin signaling among neurons and glia
during formation of tripartite synapses." Neuron Glia Biol 1: 1-11.
Engel, M., P. Maurel, et al. (1996). "Chondroitin sulfate proteoglycans in the developing central
nervous system. I. cellular sites of synthesis of neurocan and phosphacan." J Comp
Neurol 366(1): 34-43.
Evers, M. R., B. Salmen, et al. (2002). "Impairment of L-type Ca2+ channel-dependent forms of
hippocampal synaptic plasticity in mice deficient in the extracellular matrix glycoprotein
tenascin-C." J Neurosci 22(16): 7177-7194.
Faissner, A. and J. Kruse (1990). "J1/tenascin is a repulsive substrate for central nervous
system neurons." Neuron 5(5): 627-637.
- 80 -
Chapter 3
Faissner, A., M. Pyka, et al. (2010). "Contributions of astrocytes to synapse formation and
maturation - Potential functions of the perisynaptic extracellular matrix." Brain Res Rev
63(1-2): 26-38.
Ferhat, L., N. Chevassus-Au-Louis, et al. (1996). "Seizures induce tenascin-C mRNA expression
in neurons." J Neurocytol 25(9): 535-546.
Ferhat, L., N. Chevassus au Louis, et al. (1996). "Transient increase of tenascin-C in immature
hippocampus: astroglial and neuronal expression." J Neurocytol 25(1): 53-66.
Frischknecht, R., M. Heine, et al. (2009). "Brain extracellular matrix affects AMPA receptor
lateral mobility and short-term synaptic plasticity." Nat Neurosci 12(7): 897-904.
Fuss, B., E. S. Wintergerst, et al. (1993). "Molecular characterization and in situ mRNA
localization of the neural recognition molecule J1-160/180: a modular structure similar
to tenascin." J Cell Biol 120(5): 1237-1249.
Galtrey, C. M., J. C. Kwok, et al. (2008). "Distribution and synthesis of extracellular matrix
proteoglycans, hyaluronan, link proteins and tenascin-R in the rat spinal cord." Eur J
Neurosci 27(6): 1373-1390.
Garcia-Alias, G. and J. W. Fawcett (2011). "Training and anti-CSPG combination therapy for
spinal cord injury." Exp Neurol.
Geissler, M. and A. Faissner (2011). "A new indirect co-culture set up of mouse hippocampal
neurons and cortical astrocytes on microelectrode arrays." J Neurosci Methods.
Giamanco, K. A., M. Morawski, et al. (2010). "Perineuronal net formation and structure in
aggrecan knockout mice." Neuroscience 170(4): 1314-1327.
Gogolla, N., P. Caroni, et al. (2009). "Perineuronal nets protect fear memories from erasure."
Science 325(5945): 1258-1261.
Gomes, F. C., T. C. Spohr, et al. (2001). "Cross-talk between neurons and glia: highlights on
soluble factors." Braz J Med Biol Res 34(5): 611-620.
Goslin, K. and G. Banker (1989). "Experimental observations on the development of polarity by
hippocampal neurons in culture." J Cell Biol 108(4): 1507-1516.
Guimaraes, A., S. Zaremba, et al. (1990). "Molecular and morphological changes in the cat
lateral geniculate nucleus and visual cortex induced by visual deprivation are revealed
by monoclonal antibodies Cat-304 and Cat-301." J Neurosci 10(9): 3014-3024.
Gundelfinger, E. D., R. Frischknecht, et al. (2010). "Converting juvenile into adult plasticity: a
role for the brain's extracellular matrix." Eur J Neurosci 31(12): 2156-2165.
- 81 -
Chapter 3
Haas, C. A., U. Rauch, et al. (1999). "Entorhinal cortex lesion in adult rats induces the expression
of the neuronal chondroitin sulfate proteoglycan neurocan in reactive astrocytes." J
Neurosci 19(22): 9953-9963.
Hartley, R. S., M. Margulis, et al. (1999). "Functional synapses are formed between human
NTera2 (NT2N, hNT) neurons grown on astrocytes." J Comp Neurol 407(1): 1-10.
Haydon, P. G. (2001). "GLIA: listening and talking to the synapse." Nat Rev Neurosci 2(3): 185193.
Hedstrom, K. L., X. Xu, et al. (2007). "Neurofascin assembles a specialized extracellular matrix
at the axon initial segment." J Cell Biol 178(5): 875-886.
Hsia, A. Y., R. C. Malenka, et al. (1998). "Development of excitatory circuitry in the
hippocampus." J Neurophysiol 79(4): 2013-2024.
Ippolito, D. M. and C. Eroglu (2010). "Quantifying synapses: an immunocytochemistry-based
assay to quantify synapse number." J Vis Exp(45).
Irintchev, A., A. Rollenhagen, et al. (2005). "Structural and functional aberrations in the cerebral
cortex of tenascin-C deficient mice." Cereb Cortex 15(7): 950-962.
John, N., H. Krugel, et al. (2006). "Brevican-containing perineuronal nets of extracellular matrix
in dissociated hippocampal primary cultures." Mol Cell Neurosci 31(4): 774-784.
Kaech, S. and G. Banker (2006). "Culturing hippocampal neurons." Nat Protoc 1(5): 2406-2415.
Karus, M., B. Denecke, et al. (2011). "The extracellular matrix molecule tenascin C modulates
expression levels and territories of key patterning genes during spinal cord astrocyte
specification." Development 138(24): 5321-5331.
Kochlamazashvili, G., C. Henneberger, et al. (2010). "The extracellular matrix molecule
hyaluronic acid regulates hippocampal synaptic plasticity by modulating postsynaptic
L-type Ca(2+) channels." Neuron 67(1): 116-128.
Kucukdereli, H., N. J. Allen, et al. (2011). "Control of excitatory CNS synaptogenesis by
astrocyte-secreted proteins Hevin and SPARC." Proc Natl Acad Sci U S A 108(32):
E440-449.
Kwok, J. C., D. Carulli, et al. (2010). "In vitro modeling of perineuronal nets: hyaluronan
synthase and link protein are necessary for their formation and integrity." J Neurochem
114(5): 1447-1459.
Kwok, J. C., G. Dick, et al. (2011). "Extracellular matrix and perineuronal nets in CNS repair."
Dev Neurobiol.
- 82 -
Chapter 3
Lafon-Cazal, M., O. Adjali, et al. (2003). "Proteomic analysis of astrocytic secretion in the
mouse. Comparison with the cerebrospinal fluid proteome." J Biol Chem 278(27):
24438-24448.
Lander, C., H. Zhang, et al. (1998). "Neurons produce a neuronal cell surface-associated
chondroitin sulfate proteoglycan." J Neurosci 18(1): 174-183.
Matsui, F., M. Nishizuka, et al. (1998). "Occurrence of a N-terminal proteolytic fragment of
neurocan, not a C-terminal half, in a perineuronal net in the adult rat cerebrum." Brain
Res 790(1-2): 45-51.
Mauch, D. H., K. Nagler, et al. (2001). "CNS synaptogenesis promoted by glia-derived
cholesterol." Science 294(5545): 1354-1357.
Michele, M. and A. Faissner (2009). "Tenascin-C stimulates contactin-dependent neurite
outgrowth via activation of phospholipase C." Mol Cell Neurosci 41(4): 397-408.
Milner, R. and I. L. Campbell (2002). "The integrin family of cell adhesion molecules has
multiple functions within the CNS." J Neurosci Res 69(3): 286-291.
Mjaatvedt, C. H., H. Yamamura, et al. (1998). "The Cspg2 gene, disrupted in the hdf mutant, is
required for right cardiac chamber and endocardial cushion formation." Dev Biol 202(1):
56-66.
Morellini, F., E. Sivukhina, et al. (2010). "Improved reversal learning and working memory and
enhanced reactivity to novelty in mice with enhanced GABAergic innervation in the
dentate gyrus." Cereb Cortex 20(11): 2712-2727.
Nagler, K., D. H. Mauch, et al. (2001). "Glia-derived signals induce synapse formation in
neurones of the rat central nervous system." J Physiol 533(Pt 3): 665-679.
Navarrete, M. and A. Araque (2011). "Basal synaptic transmission: astrocytes rule!" Cell 146(5):
675-677.
Nikonenko, A., S. Schmidt, et al. (2003). "Tenascin-R-deficient mice show structural alterations
of symmetric perisomatic synapses in the CA1 region of the hippocampus." J Comp
Neurol 456(4): 338-349.
Perea, G., M. Navarrete, et al. (2009). "Tripartite synapses: astrocytes process and control
synaptic information." Trends Neurosci 32(8): 421-431.
Pizzorusso, T., P. Medini, et al. (2002). "Reactivation of ocular dominance plasticity in the adult
visual cortex." Science 298(5596): 1248-1251.
Porto-Pazos, A. B., N. Veiguela, et al. (2011). "Artificial astrocytes improve neural network
performance." PLoS One 6(4): e19109.
- 83 -
Chapter 3
Pyka, M., C. Busse, et al. (2011). "Astrocytes are crucial for survival and maturation of
embryonic hippocampal neurons in a neuron-glia cell-insert coculture assay." Synapse
65(1): 41-53.
Pyka, M., C. Wetzel, et al. (2011). "Chondroitin sulfate proteoglycans regulate astrocytedependent synaptogenesis and modulate synaptic activity in primary embryonic
hippocampal neurons." Eur J Neurosci.
Rathjen, F. G., J. M. Wolff, et al. (1991). "Restrictin: a chick neural extracellular matrix protein
involved in cell attachment co-purifies with the cell recognition molecule F11."
Development 113(1): 151-164.
Rauch, U., X. H. Zhou, et al. (2005). "Extracellular matrix alterations in brains lacking four of its
components." Biochem Biophys Res Commun 328(2): 608-617.
Saghatelyan, A. K., A. Dityatev, et al. (2001). "Reduced perisomatic inhibition, increased
excitatory transmission, and impaired long-term potentiation in mice deficient for the
extracellular matrix glycoprotein tenascin-R." Mol Cell Neurosci 17(1): 226-240.
Saghatelyan, A. K., S. Gorissen, et al. (2000). "The extracellular matrix molecule tenascin-R and
its HNK-1 carbohydrate modulate perisomatic inhibition and long-term potentiation in
the CA1 region of the hippocampus." Eur J Neurosci 12(9): 3331-3342.
Saghatelyan, A. K., A. G. Nikonenko, et al. (2004). "Reduced GABAergic transmission and
number of hippocampal perisomatic inhibitory synapses in juvenile mice deficient in the
neural cell adhesion molecule L1." Mol Cell Neurosci 26(1): 191-203.
Seidenbecher, C. I., K. Richter, et al. (1995). "Brevican, a chondroitin sulfate proteoglycan of rat
brain, occurs as secreted and cell surface glycosylphosphatidylinositol-anchored
isoforms." J Biol Chem 270(45): 27206-27212.
Slezak, M. and F. W. Pfrieger (2003). "New roles for astrocytes: regulation of CNS
synaptogenesis." Trends Neurosci 26(10): 531-535.
Theodosis, D. T., D. A. Poulain, et al. (2008). "Activity-dependent structural and functional
plasticity of astrocyte-neuron interactions." Physiol Rev 88(3): 983-1008.
tom Dieck, S., L. Sanmarti-Vila, et al. (1998). "Bassoon, a novel zinc-finger CAG/glutaminerepeat protein selectively localized at the active zone of presynaptic nerve terminals." J
Cell Biol 142(2): 499-509.
van den Pol, A. N. and D. D. Spencer (2000). "Differential neurite growth on astrocyte
substrates: interspecies facilitation in green fluorescent protein-transfected rat and
human neurons." Neuroscience 95(2): 603-616.
- 84 -
Chapter 3
Wang, D., R. M. Ichiyama, et al. (2011). "Chondroitinase combined with rehabilitation promotes
recovery of forelimb function in rats with chronic spinal cord injury." J Neurosci 31(25):
9332-9344.
Watanabe, H., K. Kimata, et al. (1994). "Mouse cartilage matrix deficiency (cmd) caused by a 7
bp deletion in the aggrecan gene." Nat Genet 7(2): 154-157.
Wegner, F., W. Hartig, et al. (2003). "Diffuse perineuronal nets and modified pyramidal cells
immunoreactive for glutamate and the GABA(A) receptor alpha1 subunit form a unique
entity in rat cerebral cortex." Exp Neurol 184(2): 705-714.
Wintergerst, E. S., A. Faissner, et al. (1996). "The proteoglycan DSD-1-PG occurs in
perineuronal nets around parvalbumin-immunoreactive interneurons of the rat cerebral
cortex." Int J Dev Neurosci 14(3): 249-255.
Wintergerst, E. S., B. Fuss, et al. (1993). "Localization of janusin mRNA in the central nervous
system of the developing and adult mouse." Eur J Neurosci 5(4): 299-310.
Zhang, Y., P. N. Anderson, et al. (1995). "Tenascin-C expression by neurons and glial cells in the
rat spinal cord: changes during postnatal development and after dorsal root or sciatic
nerve injury." J Neurocytol 24(8): 585-601.
- 85 -
Chapter 4
Chapter 4
A new indirect co-culture set up of mouse hippocampal
neurons and astrocytes on microelectrode arrays
Maren Geissler1 and Andreas Faissner1
1
Department of Cell Morphology and Molecular Neurobiology, Ruhr-University, D-
44780 Bochum, Germany
4.1 Abstract
Microelectrode Arrays (MEAs) are widely used to investigate neuronal network activity
in vitro at multiple sites. While this system has been successfully used with primary
embryonic rat hippocampal or cortical neurons, its applicability for mouse hippocampal
neurons has so far not been reported in detail. As mouse genetics offer a large variety
of models, it is highly desirable to close this gap. For that purpose, we established and
characterized an indirect co-culture assay of mouse hippocampal neurons in the
presence of astrocytes on MEAs. Embryonic day 15.5 (E15.5) mouse hippocampal
neurons were cultivated on MEAs in completely defined medium. We show, that the coculture with postnatal primary mouse astrocytes allows the establishment and the
maintenance of neuronal networks under these conditions. We were able to cultivate
the neurons for at least 28 days in vitro (DIV) and observed the first neuronal network
activity around 7 DIV. Hippocampal neurons showed early bursting behavior and
synchronous activity that evolved further with increasing time in culture. The
application of bicuculline increased network activity, which revealed the presence of
active gabaergic interneurons. Taken together, this study provides a novel MEA-based
assay for investigating the activity in neuronal networks in an indirect neuron-astrocyte
co-culture setting, and leads to first insights into the physiological development of
mouse hippocampal neurons under these conditions in vitro.
- 86 -
Chapter 4
4.2 Introduction
During the last ten years, the understanding that glia cells are crucial for the
maturation, maintenance and function of neurons, especially at the synapse, became
more and more established (Bacci, Verderio et al. 1999; Boehler, Wheeler et al. 2007;
Faissner, Pyka et al. 2010). In particular astrocytes are accepted as important partners,
taking part in the formation as well as in the maturation of, and the signal trafficking at
the synapse (Ullian, Christopherson et al. 2004; Christopherson, Ullian et al. 2005;
Elmariah, Hughes et al. 2005; Pyka, Wetzel et al. 2011). Without the addition of
astrocyte-derived factors, retinal ganglion cells remain silent (Pfrieger and Barres
1997) and hippocampal networks are less active, compared to glia supported cultures
(Boehler, Wheeler et al. 2007). Primary hippocampal neurons, grown in defined
medium, die within few days without the addition of astrocytes (Pyka, Busse et al.
2011). Astrocytes are known to produce numerous neuron-survival promoting factors,
underlined by a recent study describing the astrocytic transcriptome (Cahoy, Emery et
al. 2008). An important factor involved in neuronal synapse formation and stabilization,
is the neuron- and glia released extracellular matrix (ECM) (Frischknecht, Heine et al.
2009; Pyka, Wetzel et al. 2011). In particular a specialized from of the matrix, the so
called perineuronal nets (PNNs), which accumulate around a subpopulation of mature
neurons and are crucial for the limitation of synaptic plasticity (Pizzorusso, Medini et al.
2002; Gogolla, Caroni et al. 2009). For review see (Kwok, Dick et al. 2011). However,
there is still a variety of open questions concerning the interplay of astrocyte-secreted
factors in the formation and function of developing neuronal networks.
In order to achieve serum-free conditions and pure neuronal cultures to investigate the
impact of soluble astrocyte-derived factors on neuronal maturation in vitro in a
controlled manner, we designed an indirect neuron-astrocyte co-culture set-up. This
approach has previously been used under common cell culture conditions (Pyka, Busse
et al. 2011; Pyka, Wetzel et al. 2011) and has now been adapted for the handling on
Microelectrode Arrays (MEAs).
MEAs are convenient tools to investigate the development of neuronal activity in vitro
(Potter and DeMarse 2001; Hales, Rolston et al. 2010). Neurons can be cultivated on
- 87 -
Chapter 4
top of the electrodes and the evolution of the neuronal activity over time can easily be
monitored and repeatedly be assessed without being invasive. After a while in culture,
neurons become quite active, despite the lack of external input (Habets, Van Dongen et
al. 1987). Thus, investigating the pattern of spontaneous activity in such cultures
provides a versatile tool for their characterization. Further insights into a developing
neuronal network can be obtained by addition of bicuculline, which blocks the GABAA
receptor (Barbin, Pollard et al. 1993; Arnold, Hofmann et al. 2005; Boehler, Wheeler et
al. 2007)
The majority of the studies describing the development of neurons on MEAs employed
rat cells (Chiappalone, Novellino et al. 2005; Eytan and Marom 2006; Wagenaar, Pine et
al. 2006; Mazzoni, Broccard et al. 2007; Cohen, Ivenshitz et al. 2008) and to our
knowledge, there is only one study focusing on mouse hippocampal cells (Valor,
Charlesworth et al. 2007). Other studies dealing with mouse cells use cortical cultures
(Sun, Kilb et al. 2010). In general, mouse cells are more susceptible and not as easy to
handle and cultivate compared to rat cells. However, the use of embryonic mouse
hippocampal cells would provide an important tool with the possibility to study a
diversity of knock-out strains. This advantage could open new insights into the
description of genes involved in the development and in the formation and maintenance
of neuronal network communication pathways. In this perspective, we have chosen the
hippocampus as an excellent model structure to investigate open issues concerning
memory consolidation and plasticity in the central nervous system in future studies.
With regard to the role of astrocytes, one may envisage the use of genetically modified
mice, carrying mutations in systems important for neuron-glia and ECM interaction. The
indirect co-culture assay would allow for the combination of wild-type and knock-out
cells within one assay.
In the present study, we have established a new model for cultivating primary
embryonic mouse hippocampal neurons on MEA substrates. Furthermore, we present
initial results characterizing the development of neuronal network activity in these
cultures over time
- 88 -
Chapter 4
4.3 Material and methods
4.3.1 Ethical standards
All experimental procedures were performed in accordance with the Society for
Neuroscience and the German guidelines and were approved by the animal care and
utilization committees at the Ruhr University Bochum.
4.3.2 Animal housing
NMRI mice were reared in standard housing conditions, temperature and humidity
controlled, with 12h/12h light/dark cycle and food and water ad libitum.
4.3.3 Cell culture
4.3.3.1 Astrocytes
Astrocyte preparation was carried out as described previously (McCarthy and de Vellis
1980; Kaech and Banker 2006) and the protocol was adapted to mouse cells, with
some modifications. Cortices were dissected from postnatal day (P) 1-3 NMRI mouse
pups and digested in 0.1% w/v papain (Worthington, Lakewood, NJ, USA) in MEM
(Invitrogen, San Diego, CA, USA) for 1h.
Afterwards the tissue was triturated,
centrifuged and the cell pellet was resuspended in astrocyte culture medium (DMEM
(Invitrogen) with 10% v/v horse serum (Biochrom, Berlin, Germany) and 1% v/v
penicillin/streptomycin (Gibco, Karlsruhe, Germany)). Cells were plated in T-75 flasks
(Nunc, Roskilde, Denmark) with a density of 3 mouse cortices per flask in 10 ml
astrocyte medium and incubated at 37°C with 6 % v/v CO2. Complete medium changes
were performed every third day. After a cultivation time of 10 days, culture flasks were
shaken at 250 rpm and 37°C over night on an orbital shaker (New Brunswick, Edison,
NJ, USA) followed by complete medium change and addition of 20 μM cytosine-1-ß-D
- 89 -
Chapter 4
arabinofuranosid (AraC, Sigma-Aldrich,), to get rid of the oligodendrocyte precursor
cells.
48 hours before neurons were prepared (see below), astrocytes were removed from the
flasks through enzymatic digestion (0.25% w/v trypsin in MEM (both Invitrogen)) and
cells were replated in cell culture inserts with 0.4 μm pore size (BD Biosciences San
Jose, CA, USA) at a density of 25000 cells per insert in astrocyte medium (see above).
For the control staining of astrocytes (see Fig. S1) 4-well plates (Nunc, Wiesbaden,
Germany) were coated with 10 µg/ ml Poly-D-Lysine (Sigma-Aldrich) and astrocytes
were replated with a density of 25000 cells. After three days the cell were grown until
confluence and used for immunocytochemistry (see 2.5.).
4.3.3.2 Hippocampal neurons
Hippocampal neurons were prepared as described previously (Goslin and Banker 1989;
Rigato, Garwood et al. 2002; Kaech and Banker 2006; Michele and Faissner 2009) with
minor modifications to adapt to mouse cells. Hippocampi were dissected from E15.5
embryos derived from timed pregnant NMRI mice. The tissue was enzymatically
digested with 0.25% w/v trypsin (Sigma-Aldrich) in MEM (Invitrogen) for 15 minutes at
37°C. After trituration and centrifugation of the cells, the pellet was resuspended in
hippocampus medium (MEM (Invitrogen) with 10 mM sodium pyruvate, 0.1% w/v
ovalbumin (both from Sigma-Aldrich) and 2% v/v B27 (Invitrogen), see table 1) and
seeded on the MEA (Multi Channel Systems, MCS GmbH Reutlingen, Germany).
For the control stainings (see Fig. S1 and Fig. 1), the hippocampal neurons where
plated on polyornithin-coated (15 µg/ml Sigma-Aldrich) glass cover slips (Thermo
Scientific, Pittsburgh, PA, USA) and grown for 7 or 14 days under the same conditions
as MEA cultures prior to immunocytochemical analysis.
- 90 -
Chapter 4
4.3.4 Microelectrode Array recordings
4.3.4.1 MEA insert retainer
To arrange the insert above the neurons we developed an insert retainer, which was
placed around the integrated culture dish of the MEA, fixing the astrocyte layer nearby
the neurons and permitting factor exchange through the medium.
4.3.4.2 MEA boxes
To prevent evaporation and contamination of the culture, but to ensure gas exchange,
we developed special boxes for MEA cultivation. We used household plastic boxes
(Emsa, Emsdetten, Germany) and cut a 5x5 cm-sized hole into the lid. This lid was then
closed by gluing a gas but not water permeable fluorinated ethylene–propylene
membrane (FEP Teflon®, ALA MEA Sheets from ALA Scientific Instruments Inc, MCS
GmBH) on it (Potter and DeMarse 2001). The plastic boxes were completely airtight
and the gas exchange occurred only via the membrane, thus preventing evaporation.
After finishing one experiment, the boxes and membranes were cleaned with 70% v/v
EtOH and reused.
4.3.4.3 MEA preparation
For MEA recordings we used 8x8 standard MEAs with 60 titanium nitride electrodes
with a 30 µm electrode diameter and 200 µm electrode spacing, from Multichannel
Systems GMBH.
MEA preparation was performed as described (Hales et al. 2010), with slight
modifications. The area of the electrode field was coated with 0.05% v/v
Polyethylenimine (PEI, Sigma-Aldrich) for 30 minutes at room temperature. In a second
step, MEAs were coated with a 10 µl drop of 10 µg/ml laminin (Sigma-Aldrich) in PBS at
37 °C and 6% v/v CO2 for 20 minutes. Just before seeding the neurons, the laminin drop
was aspirated and 30.000 cells were plated in a 30 µl drop of hippocampus medium,
directly over the electrode field. After an adherence time of 30 minutes, the MEA was
carefully flooded with 1 ml pre-warmed hippocampus medium. Just before the
- 91 -
Chapter 4
astrocyte insert was positioned above the neurons, the medium was replaced by
hippocampus medium to avoid serum contamination. Afterwards, the astrocyte cell
culture insert was arranged in the insert retainer on top of the MEA. MEAs were placed
into the MEA boxes and incubated at 37 °C and 6% CO2. Every third day, 50% of the
medium was changed.
Control MEAs without astrocytes were handled in the same way but without astrocytes
in the insert.
Cleaning of MEAs after use was performed with incubation of 1% w/v Tergazym™
(Alconox, White Plains, NY, USA) solution over night, followed by mechanical cell debris
removal with a Cell Scraper (Sarstedt, Nümbrecht, Germany). For sterilization, MEAs
were baked at 121 °C for 15 minutes.
4.3.4.4 Electrophysiology
During recording the insert retainer was replaced by a closed lid, to prevent
contamination and to decrease loss of CO2 and shifts in pH. Spontaneous network
activity of hippocampal neurons, grown on MEAs, was recorded using set up and
software from MCS. MEAs were placed into a preamplifier (MEA1060BC, MCS) with
heating plate, which was adjusted to 35°C. After a recovery time of 20 minutes, the
spontaneous activity was recorded for 10 minutes with a sampling rate of 20 kHz. The
software MCRack (Version 3.9.0, MCS) was used to collect and analyze the data. A high
pass filter with a cut-off frequency of 200 Hz eliminated the field potentials. A
software-integrated spike detector was used to isolate the spontaneous events that
exceeded a threshold of five times of the standard deviation of the background noise.
All MEA recordings were carried out in culture medium without perfusion.
After the recordings MEAs were placed back into the incubator in co-culture with
astrocytes.
Measurements were performed at 7, 10, 12, 14, 17, 21, and 24 DIV.
Bursts were detected using the Max Interval algorithm integrated in MCRack (Legendy
and Salcman 1985). Bursts were defined using the following parameters: maximal
interval to start burst: 10 ms, maximal interval to end burst: 100 ms, maximal interval
- 92 -
Chapter 4
between bursts: 210 ms, minimal duration of a burst: 50 ms, minimal number of spikes
in bursts was 5.
4.3.4.5 Bicuculline treatment
Bicuculline treated cells were cultivated as describes above (2.4.3.) and experiments
were done after 12 DIV. MEAs were removed from the incubator and the spontaneous
activity was recorded for 5 minutes. Afterwards, 25 µM bicuculline ((-)-Bicucullinemethiodid from Sigma-Aldrich, dissolved in water) was added into the medium and
MEAs were incubated for 15 minutes, followed by another recording period of 5
minutes. The burst- and spike- parameters were the same as for the non-treated
culture (see 2.4.4.).
4.3.5 Immuncytochemistry
Three MEAs were additionally immunostained after 14 DIV. Furthermore, we have
carried out control experiments and immunostained for astrocytes and neurons in order
to check the purity of cell monolayers and for cell death (see Fig. S1). The culture
medium was carefully aspirated and neurons were fixed with 500 µl 4% w/v
paraformaldehyde (PFA, Sigma-Aldrich), for 10 minutes at room temperature.
Afterwards, the cells were washed once with PBS and two times with PBST (PBS with
0.1 % v⁄ v Triton X -100 (Sigma-Aldrich)). The primary antibodies used were the
following: neurofilament (NF200, polyclonal, rabbit, Sigma-Aldrich 1 : 300), Glial
fibrillary acidic protein (GFAP, rabbit, polyclonal and mouse IgG, both from SigmaAldrich 1 : 300), βIII-tubulin (mouse IgG, Sigma-Aldrich 1 : 200) and CD68 ( rabbit,
polyclonal, Santa Cruz Biotechnology, Heidelberg, Germany, 1:10). For the detection of
PNNs we used Wisteria floribunda agglutinin (WFA, Sigma-Aldrich; 1 : 100).
Primary antibodies and WFA were diluted in blocking buffer PBST and incubated for 30
minutes. Thereafter, cells were washed twice with PBS/A (PBS with 0.1 % w/v BSA,
Sigma-Aldrich) and afterwards secondary antibodies (subclass-specific CY3-coupled
anti rabbit antibody (1 : 500), CY2-coupled anti mouse and streptavidin coupled with
- 93 -
Chapter 4
CY2 (1 : 250)) (all secondary antibodies were derived from Dianova, Hamburg,
Germany)) with bisbenzimide (Hoechst 33528, Sigma-Aldrich, 1 : 100 000) were added
for 30 minutes in PBS/A. After two washing steps with PBS and one final step with
Milli-Q water, the cells were covered in Immumount (Thermo Scientific) with a 12 mm
glas-coverslip (Thermo Scientific).
4.3.6 Microscopy
The fixed and stained neurons were examined using an Axioplan 2 microscope,
equipped with UV-epifluorescence (Zeiss, Göttingen, Germany). Images were
photographed with a digital camera (AxioCam MRm, Zeiss) and documented using the
Axiovision 4.5 software (Zeiss). The phase-contrast images of living neurons on the
MEA were taken using the inverted microscope Axiovert 200M (Zeiss) and the
Axiovision software 4.8.1 (Zeiss).
4.3.7 Plating efficiency and cell death
To compare the efficiency of plating and the occurring cell death in cultures grown with
and without astrocytes, images of the cultures were made after 2 hours in vitro (2 HIV),
after 24 HIV; after 3, 4 and 7 DIV. Cells which attached to the MEA, appeared bright in
the phase contrast. The outgrowth of the first processes could be observed after 24
HIV,
were
detected
by
eye
and
counted
(http://rsbweb.nih.gov/ij/plugins/cell-counter.html)
with
the
for
Cell
counter
Image
Plug-in
J
(http://rsbweb.nih.gov/ij/). As a further indicator for occurring cell death we used 1
µg/ml propidiumiodid (PI) (Sigma-Aldrich). Therefore, 1 µg PI was added to the neurons
on the MEA and incubated for 30 minutes at 37 °C and 6% CO2. Afterwards, the cells
were fixed with 4% PFA for 10 minutes and washed twice with PBT1, before
bisbenzimide (Hoechst 33528, Sigma-Aldrich, 1 : 100 000 in PBS/A) was added for 30
minutes. Afterwards, cells were washed two times with PBS. Cells were covered with a
12 mm glass cover slip in Immumount and were investigated under the microscope.
Dying cells incorporated PI and appeared red.
- 94 -
Chapter 4
4.3.8 Statistics
The data is given in mean ± standard error of the mean. The mean values of two
different cultures were compared using Student´s t-test. We performed ANOVA and
Scheffé post hoc test for multiple comparisons. The significance level was set to≤ p
0.05.
4.4 Results
4.4.1 The indirect neuron astrocyte co-culture set up
Astrocytes and astrocyte-derived molecules are known to be crucial for synapse
formation, maturation, maintenance and function (Pfrieger and Barres 1997;
Christopherson, Ullian et al. 2005; Boehler, Wheeler et al. 2007; Faissner, Pyka et al.
2010; Pyka, Busse et al. 2011). To support neuronal survival, but to avoid direct
membrane-mediated astrocyte-neuron interactions we used an indirect neuronastrocyte co-culture (Pyka, Busse et al. 2011),
Table 1: Composition of the defined medium
where neurons can benefit from soluble, astrocyte-derived factors. In order to provide
neurons with beneficial astrocyte-secreted factors, and to allow the interaction of both
cell types without direct contact, we have chosen cell culture inserts with a pore size of
0.4 µm for the cultivation of astrocytes, separate from the neurons. For common cell
- 95 -
Chapter 4
culture purposes, the use of 24 well plates is sufficient to arrange the insert, but for
fixing the inserts on top of the MEA, we had to develop an insert retainer (Fig. 1 A). For
this purpose, we constructed a plastic retainer (Fig. 1, A, b), which was placed directly
beneath the integrated culture dish at the MEA and thus fixed the membranous insert
with astrocytes approximately 500 µm above the neurons. Both cell types shared the
same defined medium (see table 1) but had no direct, membrane-mediated contact.
This construction was placed into a household plastic box (Fig. 1A, d) with a permeable
fluorinated ethylene–propylene membrane (Fig. 1, A, e), to allow gas exchange within
the humidified incubator while preventing evaporation. Figure 1 gives a complete
schematic overview of the indirect MEA cell culture set up.
Figure 1: Indirect neuron astrocyte co-culture set-up for the use of MEAs
A) Schematic drawing of the indirect neuron astrocyte co-culture set up. Cultivation of neurons (white
dots) on the MEA (a) was restricted to the coated electrode field (small black bars). A custom made
insert retainer (b) was placed around the MEA ring fixing the cell culture insert (c) with feeding
astrocytes (blue stars) about 500 µm above the neurons. MEAs were placed in airtight plastic boxes (d)
with a fluorinated ethylene–propylene (FEP Teflon®) membrane (e) on top of the box, to allow gas
exchange and to simultaneously prevent evaporation of the medium. B) Neurons adhered nearby the
electrodes, built complex networks and survived up to 4 weeks in completely defined medium. The
images show neurons after 10 DIV. A higher magnification of B) is given in C). The scale bars indicate 200
µm (B) and 30 µm (C).
- 96 -
Chapter 4
In this construction neurons survived quite well. After 10 DIV, neurons (Fig. 1B, higher
magnification in C) developed numerous neurites and the complexity and density of the
network increased with time in culture. The purity of both, the neuron and the astrocyte
cultures was warranted and confirmed by immune fluorescent staining (Fig. S1).
Figure SI: Immuncytochemical staining of neurons and astrocytes
A)-F) Immuncytochemical staining of astrocytes. Astrocytes were stained with antibodies against GFAP
(A, C, E), Neurofilament 200 (A, B), βIII tubulin (C, D) and CD68 (E, F) as well as Hoechst for the detection
of cell nuclei. No contamination with other cell types than astrocytes could be detected. G)-J) Neurons
were stained with antibodies against GFAP (G, H), Neurofilament 200 (G, H) and CD68 (I, J) after 14 DIV
in culture. Cell nuclei were visualized with Hoechst. No non-neuronal cell type could be detected.
Scale Bar is 200 µm.
- 97 -
Chapter 4
The comparison of neurons grown in the indirect co-culture (Fig. 2A and B) with the
same culture grown without astrocytes (Fig. 2C and D) revealed, that pure neuronal
cultures in defined medium displayed significantly (ANOVA p = 0.034, 4 DIV: p = 0.004;
7 DIV: p = 0.003) higher rates of cell death (Fig. 2, F). Both cultures showed a high initial
loss of cells in the first 24 hours after plating (with astrocytes: 78% ± 4% plating
efficiency, without astrocytes: 67% ± 4%, p ≥ 0.05).
Figure 2: Plating efficiency and neuronal death with and without astrocytes
Comparison of the plating efficiency in neuronal cultures grown in the indirect co-culture with and
without astrocytes. Exemplary images of neurons grown in the indrect co-culture with astrocytes after
4 (A) and 7 DIV (B) and images of neurons grown without added astroglia after 4 (C) and 7 (D) DIV are
shown. Cell death and disintegrating neurites in the cultures grown without astrocytes become visible
(arrowheads in C and D). Scale Bar is 200 µm and 30µm in the higher magnifications. The quantification
of the plating efficiency is given in E). Error bars represent ± SEM, n= 3. * indicate≤ p0.05.F)
-G)
Propidiumiodid (PI) and Hoechst staining of neurons grown with (F) and without astrocytes (G, H) after 7
DIV. The incorporation of PI identified compromised neurons. Increased rates of cell death could be
demonstrated. Arrowheads show examples for cells grown on the MEA which took up the PI through the
permeable membrane. Scale bar is 30 µm.
In neuronal cultures grown in the presence of astrocytes the cell loss was stable for
the next week (3 DIV: 78% ± 4%, 4 DIV: 75 % ± 6%, 7 DIV 75 % ± 6% plating efficiency)
and did not changed significantly over time (ANOVA p = ≥ 0.05), whereas the amount of
- 98 -
Chapter 4
dying cells increased significantly (ANOVA p = 0.001) with time in vitro in cultures
without added astroglial cells compared to the initial plating density (3 DIV: 62% ± 7% p
= 0.001, 4 DIV: 48 % ± 6% p = 0.001, 7 DIV 46 % ± 9% p = 0.001plating efficiency).
Beside the detection of dying cells, degenerating neurites could be observed (higher
magnification in Fig. 2 C and D).
Figure 3: Immuncytochemical staining of hippocampal MEA cultures
A) Neuronal cultures were immunostained against Neurofilament 200 (red), WFA (green) and Hoechst
(blue) after 14 DIV. Phase contrast images were added to visualize cell bodies and neurites. Higher
magnification of the insert in A) is given in B). C) Perineuronal nets (PNNs) accumulated around a
subpopulation of neurons and were detected with Wisteria floribunda agglutinin (WFA, green), in
combination of Hoechst (blue). A phase contrast picture was added. A higher magnification of the insert
in C) is given in D). The scale bar is 100 µm (A, C) and 30 µm (B, D).
The incorporation of propidiumiodid trough permeable membranes in cells grown
without astrocytes for 7 DIV further corroborates this observation (Fig. 2, E). After 10 or
12 DIV most of the pure neuronal cultures where completely dead and showed
fragmented neurites.
- 99 -
Chapter 4
To
further
characterize
the
culture,
we
analyzed
the
neurons
using
immunocytochemistry (Fig. 3). After 14 DIV the neurofilament positive neurites crossed
the electrodes (Fig. 3, A, higher magnification in B).
4.4.2 Spontaneous activity
To characterize the development of the neurons, within the astrocyte co-culture, we
recorded the spontaneously occurring activity and monitored how neurons behave
within the first 24 days in vitro without external stimulation.
First activity, in terms of sparse spikes recorded on single electrodes, could be detected
in a portion of MEAs after around 4 or 5 DIV (data not shown), but the majority of
cultures became active around 7 DIV. Thus, we started the recordings and analyses at 7
DIV. First, we estimated the number of electrodes detecting spikes on each MEA. In Fig.
4A the mean percent of activated electrodes over time is shown. It appears that
neurons in the indirect co-culture became active very early, and from 7 days on spikes
could be recorded on most of the electrodes (83% ± 20% electrodes detected spikes) in
the majority of the cultures. The percentage of activated electrodes increased further
with time in culture (10 DIV: 88% ± 19 % of electrodes were recorded) and was
saturated at later time points (19 DIV: 98% ± 2%), with only minor variation between
different MEAs. These differences were not considered significant (ANOVA p ≥ 0.05)
Furthermore, the degree and type of neuronal activity detected at these electrodes also
changed significantly (ANOVA p = 0.03) over time. For monitoring and further
characterizing the physiological properties of the neuronal network in the indirect coculture, we decided to focus on the five electrodes with the highest detected spike rate
for further analyses.
- 100 -
Chapter 4
Figure 4: Development of electrical activity of mouse hippocampal neurons cultivated on MEAs
using the indirect co-culture system. A) Neurons were cultured for 24 DIV. After 7 DIV the cultures
(n=9) became quite active and increased the activity further within the following days, since more
electrodes detected spikes in there vicinity. B) The number of recorded spikes detected within ten
minutes recording time increased until the third week in culture. Furthermore, the burst rate (C) and the
percent of spikes occurring within bursts (D) increased with time in culture, peaked, and decreased after
21 DIV. * indicates p values≤ 0.05, the error bars represent ± SEM. N=9 E) displays a screenshot from
the MCRack display showing the recorded activity of one representative MEA within 10 minutes after 10
DIV. Each window represents the spike train recorded at the respective electrode. The scale bar indicates
5 minutes. In F), a higher magnification of the screenshot from electrode 55 (E55) is given. The scale bar
indicates 60 seconds. G) A recording from the same MEA as shown in E), after 21 DIV is displayed. The
scale bar is 5 minutes, and the red box indicates the higher magnification of E55, which is shown in H).
The appearance of rhythmic bursts after 21 DIV (G, H) becomes visible, whereas they were still missing
after 10 DIV (E, F).
- 101 -
Chapter 4
After 7 DIV a mean of 3648 ± 581 spikes was recorded per electrode. This value
increased further with time in culture and reached a peak (post hoc test p≥ 0.0 5) after
17 DIV, with 7946 ± 606 spikes per electrode. After 19 DIV there was a slight decline
(post hoc test p≥ 0.05) with a higher variability in the mean activity per electrode,
leading to a mean detection of 6036 ± 1550 spikes/electrode in 10 minutes. After 21
DIV, again a maximum of 8419 ± 1390 spikes was recorded (post hoc test
≥p
0.05),
decreasing to 4189 ± 775 spikes after 24 DIV (post hoc test p ≥ 0.05) (Fig. 4, B).
We observed that the detected spike number increased over time until a peak and
decreased after 24 DIV, but this allowed no conclusions about the network´s intrinsic
organization and spike patterns. Comparing the activity pattern in young cultures after
10 DIV (Fig. 4, E, higher magnification in F) with more mature cultures after 21 DIV (Fig.
4, G, higher magnification in H) revealed a clear change of spike patterning with time in
culture. Networks after 10 DIV showed intense and unorganized spiking behavior
without interruptions (Fig. 4, E, F) whereas MEAs after 21 DIV (Fig. 4, G, H) exhibited
periods of fast spiking with silent periods in between. These bursts could be detected
on almost all electrodes.
Furthermore, an enhanced synchrony after three weeks in culture became detectable.
All electrodes displayed comparable activity patterns (Fig. 4, G).
Using the burst analysis tool in MCRack (see 2.4.4.) we detected and counted the
occurring bursts. Quantification of the number of detected bursts (Fig. 4, C) revealed a
significantly (ANOVA p = 0.001) increasing frequency of bursts with time in culture.
After 7 DIV already 6.9 ± 1.2 bursts per minute were detected. This rate increased
further to 27.9 ± 7.3 bursts per minute after 21 DIV (p = 0.003 compared to 7DIV). After
19 and 24 DIV a small decline was apparent.
To analyze these phases of enhanced spiking frequency in more detail, the amount of
spikes occurring within bursts (% of spikes in bursts Fig. 4, D) was calculated and
significant differences were found (ANOVA p = 0.001). After 7 DIV 31.8 ± 5.4% of all
detected spikes occurred within bursts. This value increased gradually with time in
culture (10 DIV: 48.8 ± 7.6% (p≥ 0.05) ; 12 DIV: 61.8 ± 11.3% (p ≥ 0.05); 14 DIV: 67.9 ±
7.3% (p ≥ 0.05); 17 DIV: 73.8 ± 7.9% (p = 0.047) of spikes in bursts) and after 21 DIV a
significant maximum 79.3 ± 2.7% of spikes (p = 0.047) were located within bursts and
- 102 -
Chapter 4
did not occur as single events. After 24 DIV, this value dropped again to 63.2 ± 9.5% of
spikes detected within bursts (p ≥ 0.05).
4.4.3 Bursting behavior
To further characterize the bursts over time, we took a closer look at the burst
parameters. Within bursts, spikes occurred in a dramatically increased frequency
compared to the average spike frequency. This became clear when comparing the spike
frequency in general (Fig. 5, A) with the spike frequency detected within a burst (Fig. 5,
B).
Figure 5: Burst analysis of cells in MEA in indirect co-culture
A) Progression of the spike frequency [Hz] over time in culture and the frequency of spikes within bursts
(B). C) Mean duration of the recorded bursts and interburst intervals (D). In E) spike trains detected at the
electrode 55 at 10 and 21 DIV are shown. It can clearly be recognized that the rhythmic activity is more
stringently patterned after 21 DIV as compared to 10 DIV. Each of the green bars represents one spike.
Large grey bar indicates time intervals of 500 ms and in addition, the scale bar is 500ms. N=9, *≤= p
0.05
- 103 -
Chapter 4
At 7 DIV spikes were detected at low frequencies with 6 ± 0.9 Hz. That value increased
to 13.9 ± 2 Hz after 21 DIV (ANOVA p≥ 0.05). Compared to that, the frequency the
neurons exhibited within bursts was dramatically increased over time (ANOVA p =
0.002) : after 7 DIV the spike frequency within bursts amounted to 35.7 ± 3.5 Hz and
reached 65 ± 5 Hz at 21 DIV. Thus, spike frequency within bursts was a
developmentally dependent feature and increased with time. In parallel, we could
observe a slight decrease in the duration of bursts (ANOVA≥ p0.05;
Fig. 5, C). The
burst duration was longer at early time points (393 ± 39 ms) and decreased with time in
culture, especially during early maturation steps (10 DIV: 349 ± 34 ms, 12 DIV: 260 ±
34 ms). After 21 DIV we observed again the tendency to more prolonged bursts 383 ±
60 ms.
In parallel we could show that the time between bursts (inter burst interval, IBI)
decreased strongly (ANOVA p≥ 0.05) with time in culture ( Fig. 5, D). After 7 DIV the
mean IBI was 25.6 ± 8.5 seconds. After 10 DIV we found an intermediate state, where
bursts were detected with an IBI of 10.3 ± 4.5 seconds, decreasing further to 2.6 ± 0.6
seconds after 21 DIV (Fig. 5, D).
In Fig. 5 E and F spike trains, which show the typical frequencies and patterns of bursts
at 10 DIV and 21 DIV, are given. The differences in the organization of spikes become
apparent. After 21 DIV, bursts with clear silent periods between bursts were present,
whereas the spike pattern after 10 DIV was less organized.
4.4.4 Bicuculline treatment
To obtain first insights about the composition of the network concerning inhibitory and
excitatory neurons we treated some of the growing cultures with the GABAA receptor
blocker bicuculline.
The application of 25 µM bicuculline led to a significant increase (p = 0.038) of the
mean spike frequency (Fig. 6, A) after 15 minutes of incubation (before: 17.2 ± 0.9 Hz;
after bicuculline treatment: 27.4 ± 4.2 Hz) due to the blockage of GABAA receptors.
Further, we found a change in the distribution of spikes, towards a higher percentage of
spikes within bursts after the treatment with bicuculline (Fig. 6, B). Before the
application of the GABAA receptor blocker, 66.3 ± 9.8 % of spikes appeared within
- 104 -
Chapter 4
bursts. This value increased significantly (p = 0.036) to 91.2 ± 3.1 % of spikes within
bursts. Concerning the characteristics of the occurring bursts we could detect that
more spikes are located within a burst (Fig. 6, C), in line with the higher spike frequency.
The mean spikes in bursts were 47.5 ± 11.4 before the chemical stimulation and
increased significantly (p = 0.001) to 250 ± 54.6 mean spikes in a burst.
Figure 6: Bicuculline treatment
Comparison of the spike frequency (A), the percent of spikes within bursts (B), the mean spike number in
a burst (C) and the mean spike duration (D) before and after the treatment with 25 µM bicuculline is
shown. Bicuculline application led to a significant increase in the four investigated parameters. The
exemplary long term displays before (E) abd after (F) bicuculline addition indicate further changes in the
overall activity pattern. Scale Bar is 5 minutes.
Examples for the burst detection in the same recording are shown in G and H. The changed bursting
behavior becomes visible. Vertical green bars indicate a single spike. Horizontal green lines indicate a
burst detected by MC Rack. Scale Bar is 2 seconds.
- 105 -
Chapter 4
Additionally, we could demonstrate a significantly (p = 0.0013) prolonged duration of
the bursts after the application of bicuculline (Fig. 6 D; before: 400.1 ± 46.0 ms; after
bicuculline: 1580.5 ± 173.1 ms). The exemplary long term display of the recordings
from all electrodes of the MEA 18984 within the five minutes before (Fig. 6, E) and after
(Fig. 6, F) the exposure to bicuculline indicates, that there was also an overall change of
the occurring activity. The recorded activity was more repetitive, more stringently
organized and comparable for most of the electrodes.
The prolonged duration of the burst, the higher percentage of spikes within bursts and
the increased amount auf spikes in a burst could clearly be recognized in the burst
diagrams (before: Fig. 6 G; after: Fig. 6 H).
4.5 Discussion
The analysis of electrical activity using microelectrode arrays (MEAs) represents a
patent tool for the investigation of neuronal networks in vitro. So far, the use of MEAs
has been focused on rat cell cultures or cortical mouse cultures, due to their relative
robustness and compliance with standard cell culture procedures. The availability of a
growing number of genetic mutants in mice, however, opens the perspective to study
the roles of given genes in neuronal networks, using this technology. Therefore, it
seemed highly desirable to adopt the MEA technology to primary mouse embryonic
hippocampal neurons.
In the present study, we successfully developed a new cell culture paradigm that
allows for the investigation of mouse hippocampal neurons in indirect co-culture with
astrocytes, under serum-free conditions on MEAs.
The emergence of neuronal activity, especially during development, is tightly coupled
to the neurons´(Faissner, Heck et al. 2006) interaction with astrocytes and astrocyte
derived factors (Cahoy et al., 2008; Christopherson et al., 2005; Elmariah et al., 2005;
Pfrieger and Barres, 1997; Pyka et al., 2011a). Without the addition of astrocytes, pure
neuronal cultures in defined medium die within a few days as shown by the
quantification of cell death in the assay (Fig. 2 and (Pyka et al., 2011a)). Despite the
accumulating information about the astrocyte transcriptome (Cahoy et al., 2008) and
- 106 -
Chapter 4
the factors important for neuronal survival and activity (Pfrieger and Barres, 1997)
there is still a gap to close concerning the understanding of the neuron-glia interplay.
Our novel approach renders it possible to investigate neuron-glia interaction on the
physiological and network-wide level, thus permitting for the use of defined knock-out
strains of glia-related genes. The use of the indirect co-culture assay allows for the
separation of both cell types and can be utilized for the detailed investigation of the
soluble components released by astrocytes. The membrane contact mediated
interactions of both cell types can thereby be distinguished in this assay from paracrine
effects.
The expression of matrix, as revealed by staining of PNNs, further provides a possibility
to investigate matrix dependent activity changes on the MEAs.
To evaluate the experimental set-up and to investigate neuronal development, we
characterized the culture system and monitored the neuronal network development
over the first three weeks in culture by repeatedly recording spontaneous activity. This
intense spontaneous activity, which is also found in vivo, appears to be important for
network formation and functions both for the development and plasticity of neuronal
circuits (Bacci et al., 1999b; Ben-Ari, 2002; Blankenship and Feller, 2010; Garaschuk et
al., 1998; Goodman and Shatz, 1993; Lisman, 1997). When the spontaneous activity is
blocked or altered, the network shows an abnormal behavior (Cohen et al., 2008;
Mazzoni et al., 2007). Thus, spontaneous activity can be utilized as an important
hallmark to characterize the maturation of neuronal populations.
The hippocampal neurons in co-culture with astrocytes developed quite rapidly and
became active around 7 DIV, which was followed by a dramatic increase and
developmental change of activity over time. In the light of earlier reports, it was
expected, that first network activity occurred around 3 to 7 DIV as in rat hippocampal
cultures (Cohen et al., 2008). In mouse cortical cultures the activity increases
significantly between 6 and 15 DIV (Sun et al., 2010). A characteristic change of activity
patterns over time in culture was observed in agreement with other studies (Cohen et
al., 2008; Kamioka et al., 1996; Sun et al., 2010). In vitro, the synapse number of
hippocampal neurons strongly increases until the third week and tends to decrease
thereafter (Boyer et al., 1998; Li and Sheng, 2003; Papa et al., 1995; Pyka et al., 2011b).
- 107 -
Chapter 4
This is consistent with our observation, that a peak of spike- and burst frequency was
reached around 17 DIV, whereas it tended to decrease again afterwards (Fig. 3 and 4).
Phases of augmented activity and intense spontaneous activity in terms of bursts are
hallmarks of neuronal development in vivo and in vitro (Kamioka et al., 1996; Lisman,
1997) and rhythmic discharge patterns were also described by others (Arnold et al.,
2005; Bacci et al., 1999b; Murphy et al., 1992). Neurons growing in the indirect neuronastrocyte co-culture assay started bursting around 7 DIV and this bursting activity
dramatically increased with time in culture. Interestingly, a study performed with rat
cortical cultures reported bursts starting at 14 DIV, with a maximum of bursts/minute
at 21 DIV (Chiappalone et al., 2005). Interestingly, also this study describes a drop of
activity after this peak time, as observed in our culture system.
The activity pattern and especially the bursting behavior of a neuronal culture can be
changed dramatically via the application of the GABAA receptor blocker bicuculline
(Arnold et al., 2005; Barbin et al., 1993). The application of the antagonist may also
induce long lasting changes (Arnold et al., 2005), which were not subject of the present
experiments. Nevertheless, we were able to demonstrate, that neurons, grown in the
co-culture with primary astrocytes form proper networks which include GABAergic
neurons. Indeed, the treatment with bicuculline induced a significant increase in the
percentage of spikes within a burst, most probably due to the GABAA receptor
blockade. This is consistent with data derived from rat cells grown on MEAs (Arnold et
al., 2005; Hofmann and Bading, 2006) and was also reported for the hippocampus in
vivo (Buzsaki et al., 1987).
In summary, mouse hippocampal neurons cultivated in the indirect co-culture
configuration behaved comparably to other culture systems described in the literature.
The new approach, however, confers clear advantages over other approaches, namely:
i) avoiding the addition of serum; ii) opening the access to genetic mouse mutants´
neuronal hippocampal networks in vitro; iii) giving the opportunity to work with a
separated astroglial population of feeding cells.
We expect that this novel and versatile procedure will open an avenue to the
investigation of functional aspects of distinct genes in neuronal networks.
- 108 -
Chapter 4
4.6 Acknowledgements
This work was supported by the Research Department of Neuroscience of the RuhrUniversity Bochum (http://www.rd.ruhr-uni-bochum.de/neuro), the DFG GRK 736, the
research school of the Ruhr-University Bochum (GSC98/1) and the special research
program “Synapse and Glia” of the German research foundation (DFG, SPP1172).
Thanks for the support go further to Multi Channel Systems GMBH Reutlingen
especially to Dr. Frank Hofmann for critical reading of the manuscript. Further, we
thank Dipl. Biol. Michael Karus for proof reading of the manuscript and cand. B.Sc. Alina
Blusch for her help during the revision process.
- 109 -
Chapter 4
4.7 References
Arnold, F. J., F. Hofmann, et al. (2005). "Microelectrode array recordings of cultured
hippocampal networks reveal a simple model for transcription and protein synthesisdependent plasticity." J Physiol 564(Pt 1): 3-19.
Bacci, A., C. Verderio, et al. (1999). "The role of glial cells in synaptic function." Philos Trans R
Soc Lond B Biol Sci 354(1381): 403-409.
Barbin, G., H. Pollard, et al. (1993). "Involvement of GABAA receptors in the outgrowth of
cultured hippocampal neurons." Neuroscience Letters 152(1-2): 150-154.
Boehler, M. D., B. C. Wheeler, et al. (2007). "Added astroglia promote greater synapse density
and higher activity in neuronal networks." Neuron Glia Biol 3: 127-140.
Cahoy, J. D., B. Emery, et al. (2008). "A transcriptome database for astrocytes, neurons, and
oligodendrocytes: a new resource for understanding brain development and function." J
Neurosci 28(1): 264-278.
Chiappalone, M., A. Novellino, et al. (2005). "Burst detection algorithms for the analysis of
spatio-temporal patterns in cortical networks of neurons." Neurocomputing 65: 653662.
Christopherson, K. S., E. M. Ullian, et al. (2005). "Thrombospondins are astrocyte-secreted
proteins that promote CNS synaptogenesis." Cell 120(3): 421-433.
Cohen, E., M. Ivenshitz, et al. (2008). "Determinants of spontaneous activity in networks of
cultured hippocampus." Brain Res 1235: 21-30.
Elmariah, S. B., E. G. Hughes, et al. (2005). "Neurotrophin signaling among neurons and glia
during formation of tripartite synapses." Neuron Glia Biol 1: 1-11.
Eytan, D. and S. Marom (2006). "Dynamics and effective topology underlying synchronization in
networks of cortical neurons." J Neurosci 26(33): 8465-8476.
Faissner, A., N. Heck, et al. (2006). "DSD-1-Proteoglycan/Phosphacan and receptor protein
tyrosine phosphatase-beta isoforms during development and regeneration of neural
tissues." Adv Exp Med Biol 557: 25-53.
Faissner, A., M. Pyka, et al. (2010). "Contributions of astrocytes to synapse formation and
maturation - Potential functions of the perisynaptic extracellular matrix." Brain Res Rev
63(1-2): 26-38.
Frischknecht, R., M. Heine, et al. (2009). "Brain extracellular matrix affects AMPA receptor
lateral mobility and short-term synaptic plasticity." Nat Neurosci 12(7): 897-904.
- 110 -
Chapter 4
Gogolla, N., P. Caroni, et al. (2009). "Perineuronal nets protect fear memories from erasure."
Science 325(5945): 1258-1261.
Goslin, K. and G. Banker (1989). "Experimental observations on the development of polarity by
hippocampal neurons in culture." J Cell Biol 108(4): 1507-1516.
Habets, A. M., A. M. Van Dongen, et al. (1987). "Spontaneous neuronal firing patterns in fetal rat
cortical networks during development in vitro: a quantitative analysis." Exp Brain Res
69(1): 43-52.
Hales, C. M., J. D. Rolston, et al. (2010). "How to culture, record and stimulate neuronal
networks on micro-electrode arrays (MEAs)." J Vis Exp(39).
Kaech, S. and G. Banker (2006). "Culturing hippocampal neurons." Nat Protoc 1(5): 2406-2415.
Kwok, J. C., G. Dick, et al. (2011). "Extracellular matrix and perineuronal nets in CNS repair."
Dev Neurobiol.
Legendy, C. R. and M. Salcman (1985). "Bursts and recurrences of bursts in the spike trains of
spontaneously active striate cortex neurons." J Neurophysiol 53(4): 926-939.
Mazzoni, A., F. D. Broccard, et al. (2007). "On the dynamics of the spontaneous activity in
neuronal networks." PLoS One 2(5): e439.
McCarthy, K. D. and J. de Vellis (1980). "Preparation of separate astroglial and oligodendroglial
cell cultures from rat cerebral tissue." J Cell Biol 85(3): 890-902.
Michele, M. and A. Faissner (2009). "Tenascin-C stimulates contactin-dependent neurite
outgrowth via activation of phospholipase C." Mol Cell Neurosci 41(4): 397-408.
Pfrieger, F. W. and B. A. Barres (1997). "Synaptic efficacy enhanced by glial cells in vitro."
Science 277(5332): 1684-1687.
Pizzorusso, T., P. Medini, et al. (2002). "Reactivation of ocular dominance plasticity in the adult
visual cortex." Science 298(5596): 1248-1251.
Potter, S. M. and T. B. DeMarse (2001). "A new approach to neural cell culture for long-term
studies." J Neurosci Methods 110(1-2): 17-24.
Pyka, M., C. Busse, et al. (2011). "Astrocytes are crucial for survival and maturation of
embryonic hippocampal neurons in a neuron-glia cell-insert coculture assay." Synapse
65(1): 41-53.
Pyka, M., C. Wetzel, et al. (2011). "Chondroitin sulfate proteoglycans regulate astrocytedependent synaptogenesis and modulate synaptic activity in primary embryonic
hippocampal neurons." Eur J Neurosci.
Rigato, F., J. Garwood, et al. (2002). "Tenascin-C promotes neurite outgrowth of embryonic
hippocampal neurons through the alternatively spliced fibronectin type III BD domains
- 111 -
Chapter 4
via activation of the cell adhesion molecule F3/contactin." J Neurosci 22(15): 65966609.
Sun, J. J., W. Kilb, et al. (2010). "Self-organization of repetitive spike patterns in developing
neuronal networks in vitro." Eur J Neurosci 32(8): 1289-1299.
Ullian, E. M., K. S. Christopherson, et al. (2004). "Role for glia in synaptogenesis." Glia 47(3):
209-216.
Valor, L. M., P. Charlesworth, et al. (2007). "Network activity-independent coordinated gene
expression program for synapse assembly." Proc Natl Acad Sci U S A 104(11): 46584663.
Wagenaar, D. A., J. Pine, et al. (2006). "An extremely rich repertoire of bursting patterns during
the development of cortical cultures." BMC Neurosci 7: 11.
- 112 -
Chapter 5
Comprehensive Discussion and Outlook
Chapter 5
5.1 Comprehensive Discussion and 0utlook
The present study comprises the two following parts:
In chapter 3 a manuscript including the majority of the data obtained during the PhD
project is included. This manuscript is entitled “Primary hippocampal neurons, which
lack four crucial extracellular matrix molecules, display abnormalities of synaptic
structure and function and severe deficits in perineuronal net formation” (Geissler et al.,
manuscript in preparation). In this study, we used neurons and astrocytes derived from
a quadruple knock-out mouse, which is genetically deficient for the four ECM
components tenascin-C, tenascin-R, neurocan, and brevican. We performed a
combinatorial cell-culture approach, using the respective wild-type cells, and combined
immunocytochemical, protein-biochemical, and electrophysiological patch-clamp
analyses. We found out that neurons derived from the mutant displayed an altered
expression of synaptic key proteins, changes in the expression of the gammaaminobutyric acid (GABA) synthesizing enzyme Glutamate-Decarboxylase (GAD65/67),
severe deficits in the formation of perineuronal nets (PNNs), and alterations in the
frequency of postsynaptic miniature currents. Further, we reported that the reduction in
PNN complexity led to a decreased synapse formation on the respective neurons.
With chapter 4 a recently published paper entitled “A new indirect co-culture set up of
mouse hippocampal neurons and astrocytes on microelectrode arrays” (Geissler and
Faissner 2011) is included into the present thesis. The published data were obtained
during the establishment of the multi electrode array (MEA) set-up in our lab. It
describes the adaptation of the indirect neuron-astrocyte co-culture set-up to the MEA.
Prior to that, the cultivation of neurons on MEAs was mainly restricted to the more
robust rat cells and to mouse cells plated in high density with directly added glia cells
or in serum-containing medium, to promote the neuronal survival. In the present study,
we transferred the indirect neuron-astrocyte co-culture set-up in defined medium to
- 113 -
Chapter 5
Comprehensive Discussion and Outlook
the MEA and performed a detailed investigation of the spontaneous activity that
developed in the cultures over three weeks. This was completed by recordings
following antagonization of GABA with bicuculline and by immunocytochemical
approaches. Together, this novel set-up opens new possibilities to investigate neuronal
development on a network-wide level.
Comparable to the in vivo development, hippocampal neurons extend neurites, from
growth cones, polarize, undergo extensive synaptogenesis and become active in vitro
(Basarsky, Parpura et al. 1994; Verderio, Coco et al. 1999; Kaech and Banker 2006).
Therefore, the use of cell cultures of primary hippocampal neurons is an elegant tool to
study a diversity of these aspects of neuronal development in vitro (Kaech and Banker
2006; Pyka, Busse et al. 2011; Pyka, Wetzel et al. 2011; Jones, Cook et al. 2012). The
indirect co-culture of neurons with a supporting monolayer of astrocytes allows for the
cultivation of these cells in defined media for at least three weeks (Kaech and Banker
2006), while the substitution of astrocytes by fibroblasts or glial cell-lines does not
lead to the same survival-promoting effects (Pyka, Busse et al. 2011). Accordingly,
neurons grown in defined medium without supporting astrocytes die within a few days
(Pyka, Busse et al. 2011).
Beyond that, the support of astrocyte-released factors has been shown to be
indispensable for proper synapse formation and synaptic activity of neuronal networks
in vitro (Christopherson, Ullian et al. 2005; Boehler, Wheeler et al. 2007; Pyka, Busse et
al. 2011; Jones, Cook et al. 2012). Some putative astrocyte-released synaptogenic
molecules and their receptors have been unraveled so far. Along these lines,
cholesterol, apoliprotein E and thrombospondin, binding to the calcium receptor
alpha2delta-1, have been shown to exert synaptogenic properties in vitro (Mauch,
Nagler et al. 2001; Nagler, Mauch et al. 2001; Christopherson, Ullian et al. 2005;
Eroglu, Allen et al. 2009). However, the addition of selected factors in vitro did not lead
to complete functional synapse formation in the absence of astrocytes (Christopherson,
Ullian et al. 2005). Therefore, it is assumed that further glia-derived molecules are
involved in these processes.
- 114 -
Chapter 5
Comprehensive Discussion and Outlook
There accumulated evidence that extracellular matrix (ECM) molecules play a
particular role in synapse formation and synaptic activity (Faissner, Pyka et al. 2010;
Dityatev and Rusakov 2011). Studies focusing on the role of the ECM during synapse
formation and synaptic transmission during maturation were performed recently in our
lab (Pyka, Wetzel et al. 2011). Via the application of ChondroitinaseABC (ChABC), an
enzyme degrading the covalently attached glycosaminoglycan (GAG) chains of
chondroitinsulfate proteoglycans (CSPGs), the regulatory impact of the ECM on the
formation and function of synapses in the system was investigated in detail (Pyka,
Wetzel et al. 2011). Thus, Pyka and colleagues found a significantly increased
formation of synaptic puncta, paralleled with a decrease in the amplitude and charge of
miniature postsynaptic excitatory currents (mEPSCs) in primary rat hippocampal cells.
However, there are still innumerable questions concerning the interplay of neurons,
astrocytes, and the ECM, especially with regard to the emerging picture of the
tetrapartite synapse (Dityatev and Rusakov 2011).
The use of mouse cells in the described assay allows now for the utilization of
genetically modified ECM. This may help to answer open questions concerning the
detailed role of the ECM during synapse development. The present study delivered first
important steps in this direction.
The first manuscript focuses on the detailed characterization of ECM deficient primary
hippocampal neurons in a combinatorial approach with primary astrocytes. Because of
the investigation of a diversity of aspects over a long time in vitro, this work nicely
contributed to the existing knowledge about the role of the ECM in proper synapse
formation and synaptic transmission in vitro.
Via in vitro immunocytochemical detection of the synaptic key proteins bassoon and
PSD95 in neurons lacking the four ECM molecules tenascin-C, tenascin-R, neurocan
and brevican, we wanted to investigate the crucial role of the ECM in synapse
formation. Structural synapses started to form around 7 days in vitro (DIV) and
increased over time in culture. In mutant neurons we found an increased expression of
synaptic key proteins after 14 DIV followed a significantly decreased synaptic puncta
expression in more mature ECM deficient cultures after 21 DIV compared to the wild- 115 -
Chapter 5
Comprehensive Discussion and Outlook
type. In contrast to the enzymatic ECM disruption, which previously revealed an
increased synapse formation after 13 as well as after 20 DIV (Pyka, Wetzel et al. 2011),
we have found a dual role of the ECM that became apparent after genetic ECM
manipulation. The extracellular matrix (ECM) of the brain was already known to have a
strong regulatory impact on the formation, maintenance, and function of neuronal
circuits during development (for review see (Dityatev, Schachner et al. 2010). While the
“early” ECM constrains and regulates the synapse formation in vitro, the “more mature”
ECM is supposed to maintain and stabilize synaptic connections (Dityatev, Schachner et
al. 2010; Pyka, Wetzel et al. 2011). Along these lines, the enzymatic digestion of the
characteristic negatively charged GAG chains confirmed the functional importance for
maintaining synaptic connections and restricting plasticity in the adult brain. Thus, the
digestion of ECM components led to juvenile levels of plasticity in the adult (Pizzorusso,
Medini et al. 2002; Gogolla, Caroni et al. 2009). The two distinct effects observed in our
assay, are thought to root in different molecular mechanisms, which were not subject
of the present investigations. Nevertheless, different aspects are discussed in the
literature. One possible explanation is the alteration in the molecular configuration of
the ECM over time. Thus, the composition of the ECM is known to change dramatically
during development and the distinct expression of ECM molecules depends highly on
the spatial and temporal requirements (Jones and Jones 2000; Wilson and Snow 2000;
Galtrey, Kwok et al. 2008). For example, tenascin-C is highly expressed in the
developing brain but the expression is restricted in the adult brain (Ferhat, Chevassus
au Louis et al. 1996) while a CNS lesion can lead to a dramatic up-regulation of
tenascin-C in the adult (Deller, Haas et al. 1997). In contrast, brevican was assumed to
be part of the matured ECM (Yamaguchi 1996) and the same was described for
tenascin-R, a prominent component of PNNs (Carulli, Rhodes et al. 2006).
Nevertheless, other reports described neurocan and brevican to be expressed also in
the developing brain (Pyka, Wetzel et al. 2011). However, the distinct developmental
regulation of the ECM molecules is crucial for proper neuronal development (Bandtlow
and Zimmermann 2000; Wilson and Snow 2000; Kwok, Warren et al. 2012). The precise
timing of the ECM expression and the changes in the ECM composition will result in
different cellular interactions with the ECM and different receptor binding, thereby
- 116 -
Chapter 5
Comprehensive Discussion and Outlook
explaining the dual role of the ECM. One can assume that the expression of ECM
molecules changes over time in the present assay. As a proof of principle, we found all
of the mentioned four ECM molecules to be expressed in the used assay and in the
immature hippocampus at E15.5 in vivo. Nevertheless, we recommend for future
experiments, to investigate the detailed developmental expression of the four crucial
ECM molecules at different time points in vivo and in vitro. Although this was already
described for other models (Pyka, Wetzel et al. 2011) it will give new hints concerning
the impact of the ECM in the given assay. Further, one should check the expression of
additional important ECM molecules, such as versican, aggrecan, phosphacan, and
RPTP beta/zeta. Another reason for the described dual role of the ECM during
development may lie in the changing receptor repertoire expressed by the neurons over
time. In this direction, the investigation of the ECM receptor expression patterns in the
cultures is recommended for the future. Changes in the expression of ECM receptors,
such as integrins were described in the past (Lathia, Patton et al. 2007), but studies
concerning the ECM receptor expression in vitro are sparse. However, the importance of
ECM receptors in shaping synaptic structure and function was recently demonstrated
by the observed close interaction of β3 integrin and AMPA receptors (Pozo, Cingolani et
al. 2012). In general, it can be assumed that the neuron-ECM interaction is tightly
adjusted by both partners, if not by three including the glial part (Araque, Parpura et al.
1999), and that neuronal activity is crucially involved in these mechanisms.
Therefore, the immunocytochemical investigations concerning the matrix expression
and the formation of synapses in vitro were paralleled by whole-cell voltage-clamp
recordings (performed in cooperation with Dr. rer. nat. Ainhara Aguado in the lab of
Prof. Dr. Dr. Dr. Hanns Hatt). As described and discussed in chapter 3, the collaborators
were able to detect significant alterations in synaptic transmission when neurons were
derived from the mutant tissue. Thus, the matrix deficiency led to a significant
reduction in the frequency of miniature postsynaptic currents (mPSPs). In line with the
immunocytochemcial data, the phenotype was particularly attenuated when neurons
were derived from the mutant, almost independent of the astrocytic phenotype.
Interestingly, this decrease was found in inhibitory as well as in excitatory miniature
postsynaptic currents. However, the amplitude of both postsynaptic potentials
- 117 -
Chapter 5
Comprehensive Discussion and Outlook
measured remained unchanged. Nevertheless, this physiological phenotype could not
be directly explained by the expression of the investigated synaptic key proteins, as we
assumed. On the contrary, we have found an increased synapse formation after 14 DIV
paralleled with a decreased frequency of mPSCs and a decreased synapse formation
combined with a decrease in the mPSC frequency after 21 DIV. However, as discussed
in chapter 3, at this point, we can not decide whether the immunocytochemically
detectable synapses were also physiologically active. The formation of silent synapses
in the absence of the four crucial matrix molecules could provide an explanation for the
reduced frequency of mPSCs. The speculative idea of the un-silencing of AMPA
responsiveness-lacking synapses via astrocyte secreted ECM molecules has also been
proposed before (Christopherson, Ullian et al. 2005).
Beyond that, the impact of the ECM on synaptic transmission and synaptic plasticity
was already shown in a diversity of knock-out mice lacking distinct ECM components.
Accordingly, the tenascin-C deficient mice exhibit changes in the local field potentials in
freely moving mice (Gurevicius, Kuang et al. 2009) and reduced LTP in the CA1 region
of the hippocampus (Evers, Salmen et al. 2002). In line with that, a reduction in LTP
was also confirmed in the tenascin-R mutant (Bukalo, Schachner et al. 2001;
Saghatelyan, Dityatev et al. 2001) and mild deficits in the neurocan (Zhou, Brakebusch
et al. 2001) and brevican knock-out mice (Brakebusch, Seidenbecher et al. 2002) were
reported.
The observed modifications in synaptic transmission in the matrix deficient neurons
may also root in shifts of the neuronal subpopulations. To unravel such changes we
have made first attempts using protein biochemical approaches, so far without
conclusive results. In further studies we will try to estimate the detailed ratio of the
inhibitory and excitatory neuronal subpopulations in our model. In this direction, we
have already performed initial investigations in cooperation with Prof. Dr. Georg Juckel
and PD. Dr. Rainer Wolf at the Ruhr-University Bochum. Within this cooperation we
want to perform high-performance liquid chromatography (HPLC) experiments to
determine the exact amounts of gamma-aminobutyric acid (GABA) and glutamate in
lysates from wild-type and mutant neuronal cultures. This technique has been
established for serotonin and dopamine in the aforementioned lab (Winter, Reutiman et
- 118 -
Chapter 5
Comprehensive Discussion and Outlook
al. 2008) and is now available for GABA and glutamate. These measurements are
assumed to reveal details about shifts in the inhibitory and excitatory neuronal
subpopulations in vitro. In addition, we will do further attempt with immuncytochemical
stainings to unravel the reasons for the increased GAD 65/67 expression, observed in
protein lysates from mutant cultures. We want to figure out if the overall GAD65/67
expression is increased, if more GABAergic neurons can be found, or if less GABAergic
neurons express relatively more GAD 65/67. In addition, immuncytochemical detection
of vGLUT will reveal if the unaffected relative vGLUT expression is confirmed on a
cellular level or if there are less glutamatergic neurons, expressing higher levels of
vGLUT. Thus, determining the exact population-related changes in the mutant cultures
will be a key aspect of further investigations. In addition, we will focus on in vivo
experiments. For instance, we will compare the in vivo expression pattern of the
subpopulations at different developmental stages in both genotypes. In these lines, we
have made first immunocytochemical stainings against vGLUT, GABA, GAD 65/67,
Calbindin, and Parvalbumin in vivo. These preliminary experiments will be continued in
the future and are thought to lead to a comprehensive study of the in vivo situation in
the quadruple knock-out mouse. Changes in the neuronal density as well as in the
inhibitory neuronal population were previously described in the tenascin-C deficient
mice (Irintchev, Rollenhagen et al. 2005). Thus, the cortex of the mutant mice exhibited
abnormal high neuronal density with a low density of parvalbumin-positive neurons and
an overall and reduced ratio of inhibitory to excitatory neurons (Irintchev, Rollenhagen
et al. 2005). Thus, the intended in vivo analyses will probably contribute to the existing
knowledge about the CNS development.
The electrophysiological in vitro results are also scheduled to become completed by in
and ex vivo recordings. For that purpose we have already initiated two cooperations.
Together with the lab of Prof. Dr. Denise Manahan-Vaughan at the Ruhr-University
Bochum we will perform stimulations of the perforant pathway and recordings in the
dentate gyrus of the hippocampus in both genotypes. Via recordings of excitatory fieldpotentials (fEPSP) we want to detect changes in the mechanisms of synaptic plasticity
in these mice. With regard to the aforementioned changes in synaptic plasticity in
several ECM mutants (Bukalo, Schachner et al. 2001; Evers, Salmen et al. 2002), the
- 119 -
Chapter 5
Comprehensive Discussion and Outlook
investigations in mice lacking four crucial parts of the ECM will hopefully shed new
light on the ECM-mediated mechanism in synaptic plasticity.
Along these lines, we have already started with experiments focusing on the largescale
neuronal
network
of
the
ECM
deficient
mice
via
high-density
electroencephalogram (EEG) recordings together with the lab of Prof. MD. Jozsef
Zoltan Kiss and PhD Charles Quairiaux form the University of Geneva (Megevand,
Quairiaux et al. 2008; Quairiaux, Sizonenko et al. 2010; Quairiaux, Megevand et al.
2011). These collaborations are expected to complete the in vitro data on a systembiological level and will give further hints concerning the phenotype of the quadruple
ECM mutant. Complementary to these in vivo investigations in progress, we have
already completed the in vivo analysis of PNNs in the hippocampus as well as in the
cortex of mutant mice. Via immunohistochemical visualization of the PNNs with
Wisteria Floribunda agglutinin (WFA) we found a severe reduction of the PNN density
and complexity in the hippocampus as well as in the cortex of mutant mice. We
performed a developmental analysis of the PNN expression patterns at day postnatal
(P) 0, P5, P10, P15, P20, P25, and P35 in both CNS regions. The diminished PNN
formation found at all developmental stages was pronounced in the CA3 region, which
displayed an intense WFA staining under control conditions. This characteristic region
of intensive WFA staining has also been described before (Bruckner, Grosche et al.
2003). These data are not included into the present work and will lead to a separate
manuscript focusing on the in vivo consequences of the ECM deficiency.
Nevertheless, the observed reductions in PNN wearing neurons combined with the
reduced frequency in inhibitory mPSCs found in vitro provide a first hint into this
direction. PNNs are described to be expressed mainly on inhibitory GABAergic neurons
(Dityatev, Bruckner et al. 2007; Balmer, Carels et al. 2009) and they occupy a pivotal
role within the neuron-ECM relationship. Tightly enwrapping the neuronal cell body and
proximal dendrites they stabilize the mature synapse (for review see (Faissner, Pyka et
al. 2010). Despite the intense research focusing on PNNs, the knowledge about their
developmental importance is still fragmentary. The observed deficits in the formation
and maintenance of PNNs in mutant neurons in vitro provide a new tool for studies in
this direction and this will probably shed new light on the role of PNNs during neuronal
- 120 -
Chapter 5
Comprehensive Discussion and Outlook
network-formation. Studies in the aggrecan knock-out mice (Giamanco, Morawski et al.
2010) and in the Crtl-1 mutant (Carulli, Pizzorusso et al. 2010), lacking the Cartilage
link protein 1, revealed a lot of details about the initiation and formation of PNNs, such
as the sequential recruitment of distinct PNN components. In contrast, we aimed to
obtain details about the consequences of a matrix deficiency during neuronal
development and synapse formation. We observed that the ECM deficiency led to very
restricted and narrow PNN formation in vitro and as a functional correlate of this
phenotype we observed that neurons with only rudimentary PNNs express significantly
less synaptic proteins on their surface. The reduced synapse formation on these
neurons can have a diversity of reasons, which should be subject of further
investigations. However, one can discuss a few possible mechanisms with respect to
the function of PNNs. Thus, PNNs are known to be important in buffering local ion
concentrations, accumulating growth factors, thereby supporting highly active neurons
(Hartig, Derouiche et al. 1999; Dityatev, Bruckner et al. 2007). Along these lines, the
diminished complexity of the PNNs in the mutant could have led to changes in neuronal
functionality, resulting in a decreased synapse formation. On the other hand, the
deficits in proper synapse formation could have led to the observed deficits in PNN
formation. This would nicely fit to the described electrophysiological changes in the
whole network on randomly selected neuron. There exist a couple of publications,
showing that the PNN formation depends highly on neuronal activity (Lander, Kind et al.
1997; Dityatev, Bruckner et al. 2007). Thus, blocking neuronal activity has been shown
to lead to diminished PNN formation (Dityatev, Bruckner et al. 2007). Recent data
revealed that semaphorin 3A (sema 3A), an ECM molecule crucially involved in axon
guidance, localizes within PNNs (De Wit, De Winter et al. 2005) and unpublished data/
personal correspondence with Prof. Dr. James Fawcett from the University of
Cambridge). Interestingly, sema 3A was also shown to be involved in the synaptic
function of hippocampal neurons (Bouzioukh, Daoudal et al. 2006). These experiments
showed a decreased synaptic activity and decreased expression of synaptic key
proteins after exogenous application (Bouzioukh, Daoudal et al. 2006). If the sema 3A
expression is altered in the mutant neurons is subject of current investigation. However,
first immunocytochemical stainings revealed at least the expression of sema 3A in our
- 121 -
Chapter 5
Comprehensive Discussion and Outlook
assay and we will especially focus on investigations of the reduced synapse formation
on the rudimentary PNN wearing neurons in the future.
In sum the data concerning the matrix-deficiency, during synapse formation and
maintenance in vitro nicely revealed that the absence of tenascin-C, tenascin-R,
neurocan and brevican led to distinct phenotypes in young and more mature cultures. A
rather new and interesting concept of the present study is that the neuronal
compartment in the defined assay was capable of highly contributing to the matrix
production, while the addition of wild-type astrocytes could not rescue the observed
phenotypes. These observations point to a fundamental participation of neurons in the
ECM production under defined conditions in vitro. To corroborate these observations, it
would be reasonable to perform real time PCR experiments or in situ hybridization
experiments, to manifest the neuronal ECM expression on mRNA levels. With regard to
the synapse formation under matrix deficient conditions, it would be further advisable
to investigate the synapse formation at the earliest steps around 5/7 days in vitro (DIV).
For future experiments, we thought about performing the whole cell voltage-clamp
recordings, especially on the PNN wearing neurons instead of randomly picked neurons.
This might provide further hints with regard to the physiological changes on the PNN
deficient neurons in the mutant cultures. Further, it is also reasonable to combine the
investigations concerning the PNN deficiency with the MEA set-up. While the patch
clamp experiments performed in chapter 3 delivered data on a single cell level, the
implementation of MEA recordings will allow for the quantifications of a given culture
on a network-wide level. A further advantage of the MEA assay is that the cultures can
be subject of investigation over several weeks, instead of measuring two different
cultures at two different time points (14 and 21 DIV). First steps in this direction were
already made by the visualization of the PNNs in the cultures grown on MEAs. In
addition, first cultures of neurons from the quadruple knock-out were grown and
investigated on the MEA and this will be followed up intensively in the future.
The first experiments performed on the MEA and published in 2011 (Geissler and
Faissner 2011) dealt with the establishment of the indirect co-culture set-up for mouse
derived cells and monitored the spontaneous activity over an extended time in vitro. In
addition, we performed initial stimulation experiments via the addition of bicuculline,
- 122 -
Chapter 5
Comprehensive Discussion and Outlook
blocking GABAA receptors. In this direction, further emphasis will be given to the
investigation of the bicuculline induced activation in the mutant derived cultures.
Additionally, one can think about electrical stimulation, which is also possible with the
MEA set-up (Jun, Hynd et al. 2007; Jun, Smith et al. 2010). We assume that these
investigations will give further hints concerning a possible imbalance in neuronal
inhibition and excitation in the mutant. Regarding long-term considerations, we plan to
establish ex vivo recordings with hippocampal slices on the MEA in our lab. With these
experiments, the use of LTP- or paired-pulse protocols becomes possible (Steidl, Neveu
et al. 2006) and should reveal further details about the role of the crucial matrix
molecules in synaptic plasticity.
Overall, the performed work nicely contributed to the existing knowledge about the
impact of the extracellular matrix in synapse formation, maturation, and synaptic
plasticity and opened possibilities for further investigations on a network-wide level.
Together with future experiments, focusing on the downstream mechanisms and
combined with the use of ECM mutants in the MEA approach, these studies should
reveal the underlying processes of the ECM-synapse relationship. The use of genetic
manipulation of main PNN components, as in the investigated quadruple knock-out
mouse, complementary to the enzymatic modification, will shed new light on the ECM
function during neuronal development.
- 123 -
Chapter 5
Comprehensive Discussion and Outlook
5.2 References
Araque, A., V. Parpura, et al. (1999). "Tripartite synapses: glia, the unacknowledged partner."
Trends Neurosci 22(5): 208-215.
Balmer, T. S., V. M. Carels, et al. (2009). "Modulation of perineuronal nets and parvalbumin with
developmental song learning." J Neurosci 29(41): 12878-12885.
Bandtlow, C. E. and D. R. Zimmermann (2000). "Proteoglycans in the developing brain: new
conceptual insights for old proteins." Physiol Rev 80(4): 1267-1290.
Basarsky, T. A., V. Parpura, et al. (1994). "Hippocampal synaptogenesis in cell culture:
developmental time course of synapse formation, calcium influx, and synaptic protein
distribution." J Neurosci 14(11 Pt 1): 6402-6411.
Boehler, M. D., B. C. Wheeler, et al. (2007). "Added astroglia promote greater synapse density
and higher activity in neuronal networks." Neuron Glia Biol 3: 127-140.
Bouzioukh, F., G. Daoudal, et al. (2006). "Semaphorin3A regulates synaptic function of
differentiated hippocampal neurons." Eur J Neurosci 23(9): 2247-2254.
Brakebusch, C., C. I. Seidenbecher, et al. (2002). "Brevican-deficient mice display impaired
hippocampal CA1 long-term potentiation but show no obvious deficits in learning and
memory." Mol Cell Biol 22(21): 7417-7427.
Bruckner, G., J. Grosche, et al. (2003). "Region and lamina-specific distribution of extracellular
matrix proteoglycans, hyaluronan and tenascin-R in the mouse hippocampal formation."
J Chem Neuroanat 26(1): 37-50.
Bukalo, O., M. Schachner, et al. (2001). "Modification of extracellular matrix by enzymatic
removal of chondroitin sulfate and by lack of tenascin-R differentially affects several
forms of synaptic plasticity in the hippocampus." Neuroscience 104(2): 359-369.
Carulli, D., T. Pizzorusso, et al. (2010). "Animals lacking link protein have attenuated
perineuronal nets and persistent plasticity." Brain 133(Pt 8): 2331-2347.
Carulli, D., K. E. Rhodes, et al. (2006). "Composition of perineuronal nets in the adult rat
cerebellum and the cellular origin of their components." J Comp Neurol 494(4): 559577.
Christopherson, K. S., E. M. Ullian, et al. (2005). "Thrombospondins are astrocyte-secreted
proteins that promote CNS synaptogenesis." Cell 120(3): 421-433.
- 124 -
Chapter 5
Comprehensive Discussion and Outlook
De Wit, J., F. De Winter, et al. (2005). "Semaphorin 3A displays a punctate distribution on the
surface of neuronal cells and interacts with proteoglycans in the extracellular matrix."
Mol Cell Neurosci 29(1): 40-55.
Deller, T., C. A. Haas, et al. (1997). "Up-regulation of astrocyte-derived tenascin-C correlates
with neurite outgrowth in the rat dentate gyrus after unilateral entorhinal cortex lesion."
Neuroscience 81(3): 829-846.
Dityatev, A., G. Bruckner, et al. (2007). "Activity-dependent formation and functions of
chondroitin sulfate-rich extracellular matrix of perineuronal nets." Dev Neurobiol 67(5):
570-588.
Dityatev, A. and D. A. Rusakov (2011). "Molecular signals of plasticity at the tetrapartite
synapse." Curr Opin Neurobiol 21(2): 353-359.
Dityatev, A., M. Schachner, et al. (2010). "The dual role of the extracellular matrix in synaptic
plasticity and homeostasis." Nat Rev Neurosci 11(11): 735-746.
Eroglu, C., N. J. Allen, et al. (2009). "Gabapentin receptor alpha2delta-1 is a neuronal
thrombospondin receptor responsible for excitatory CNS synaptogenesis." Cell 139(2):
380-392.
Evers, M. R., B. Salmen, et al. (2002). "Impairment of L-type Ca2+ channel-dependent forms of
hippocampal synaptic plasticity in mice deficient in the extracellular matrix glycoprotein
tenascin-C." J Neurosci 22(16): 7177-7194.
Faissner, A., M. Pyka, et al. (2010). "Contributions of astrocytes to synapse formation and
maturation - Potential functions of the perisynaptic extracellular matrix." Brain Res Rev
63(1-2): 26-38.
Ferhat, L., N. Chevassus au Louis, et al. (1996). "Transient increase of tenascin-C in immature
hippocampus: astroglial and neuronal expression." J Neurocytol 25(1): 53-66.
Galtrey, C. M., J. C. Kwok, et al. (2008). "Distribution and synthesis of extracellular matrix
proteoglycans, hyaluronan, link proteins and tenascin-R in the rat spinal cord." Eur J
Neurosci 27(6): 1373-1390.
Geissler, M. and A. Faissner (2011). "A new indirect co-culture set up of mouse hippocampal
neurons and cortical astrocytes on microelectrode arrays." J Neurosci Methods.
Giamanco, K. A., M. Morawski, et al. (2010). "Perineuronal net formation and structure in
aggrecan knockout mice." Neuroscience 170(4): 1314-1327.
Gogolla, N., P. Caroni, et al. (2009). "Perineuronal nets protect fear memories from erasure."
Science 325(5945): 1258-1261.
- 125 -
Chapter 5
Comprehensive Discussion and Outlook
Gurevicius, K., F. Kuang, et al. (2009). "Genetic ablation of tenascin-C expression leads to
abnormal hippocampal CA1 structure and electrical activity in vivo." Hippocampus
19(12): 1232-1246.
Hartig, W., A. Derouiche, et al. (1999). "Cortical neurons immunoreactive for the potassium
channel Kv3.1b subunit are predominantly surrounded by perineuronal nets presumed
as a buffering system for cations." Brain Res 842(1): 15-29.
Irintchev, A., A. Rollenhagen, et al. (2005). "Structural and functional aberrations in the cerebral
cortex of tenascin-C deficient mice." Cereb Cortex 15(7): 950-962.
Jones, E. V., D. Cook, et al. (2012). "A neuron-astrocyte co-culture system to investigate
astrocyte-secreted factors in mouse neuronal development." Methods Mol Biol 814:
341-352.
Jones, F. S. and P. L. Jones (2000). "The tenascin family of ECM glycoproteins: structure,
function, and regulation during embryonic development and tissue remodeling." Dev
Dyn 218(2): 235-259.
Jun, S. B., M. R. Hynd, et al. (2007). "Electrical stimulation-induced cell clustering in cultured
neural networks." Med Biol Eng Comput 45(11): 1015-1021.
Jun, S. B., K. L. Smith, et al. (2010). "Optical monitoring of neural networks evoked by focal
electrical stimulation on microelectrode arrays using FM dyes." Med Biol Eng Comput
48(9): 933-940.
Kaech, S. and G. Banker (2006). "Culturing hippocampal neurons." Nat Protoc 1(5): 2406-2415.
Kwok, J. C., P. Warren, et al. (2012). "Chondroitin sulfate: A key molecule in the brain matrix."
Int J Biochem Cell Biol.
Lander, C., P. Kind, et al. (1997). "A family of activity-dependent neuronal cell-surface
chondroitin sulfate proteoglycans in cat visual cortex." J Neurosci 17(6): 1928-1939.
Lathia, J. D., B. Patton, et al. (2007). "Patterns of laminins and integrins in the embryonic
ventricular zone of the CNS." J Comp Neurol 505(6): 630-643.
Mauch, D. H., K. Nagler, et al. (2001). "CNS synaptogenesis promoted by glia-derived
cholesterol." Science 294(5545): 1354-1357.
Megevand, P., C. Quairiaux, et al. (2008). "A mouse model for studying large-scale neuronal
networks using EEG mapping techniques." Neuroimage 42(2): 591-602.
Nagler, K., D. H. Mauch, et al. (2001). "Glia-derived signals induce synapse formation in
neurones of the rat central nervous system." J Physiol 533(Pt 3): 665-679.
Pizzorusso, T., P. Medini, et al. (2002). "Reactivation of ocular dominance plasticity in the adult
visual cortex." Science 298(5596): 1248-1251.
- 126 -
Chapter 5
Comprehensive Discussion and Outlook
Pozo, K., L. A. Cingolani, et al. (2012). "beta3 integrin interacts directly with GluA2 AMPA
receptor subunit and regulates AMPA receptor expression in hippocampal neurons."
Proc Natl Acad Sci U S A.
Pyka, M., C. Busse, et al. (2011). "Astrocytes are crucial for survival and maturation of
embryonic hippocampal neurons in a neuron-glia cell-insert coculture assay." Synapse
65(1): 41-53.
Pyka, M., C. Wetzel, et al. (2011). "Chondroitin sulfate proteoglycans regulate astrocytedependent synaptogenesis and modulate synaptic activity in primary embryonic
hippocampal neurons." Eur J Neurosci 33(12): 2187-2202.
Quairiaux, C., P. Megevand, et al. (2011). "Functional development of large-scale sensorimotor
cortical networks in the brain." J Neurosci 31(26): 9574-9584.
Quairiaux, C., S. V. Sizonenko, et al. (2010). "Functional deficit and recovery of developing
sensorimotor networks following neonatal hypoxic-ischemic injury in the rat." Cereb
Cortex 20(9): 2080-2091.
Saghatelyan, A. K., A. Dityatev, et al. (2001). "Reduced perisomatic inhibition, increased
excitatory transmission, and impaired long-term potentiation in mice deficient for the
extracellular matrix glycoprotein tenascin-R." Mol Cell Neurosci 17(1): 226-240.
Steidl, E. M., E. Neveu, et al. (2006). "The adult rat hippocampal slice revisited with multielectrode arrays." Brain Res 1096(1): 70-84.
Verderio, C., S. Coco, et al. (1999). "Synaptogenesis in hippocampal cultures." Cell Mol Life Sci
55(11): 1448-1462.
Wilson, M. T. and D. M. Snow (2000). "Chondroitin sulfate proteoglycan expression pattern in
hippocampal development: potential regulation of axon tract formation." J Comp Neurol
424(3): 532-546.
Winter, C., T. J. Reutiman, et al. (2008). "Dopamine and serotonin levels following prenatal viral
infection in mouse--implications for psychiatric disorders such as schizophrenia and
autism." Eur Neuropsychopharmacol 18(10): 712-716.
Yamaguchi, Y. (1996). "Brevican: a major proteoglycan in adult brain." Perspect Dev Neurobiol
3(4): 307-317.
Zhou, X. H., C. Brakebusch, et al. (2001). "Neurocan is dispensable for brain development." Mol
Cell Biol 21(17): 5970-5978.
- 127 -
Chapter 5
Summary
5.2 Summary
The research of the last decade provided evidence for the fundamental role the
extracellular matrix (ECM) plays during neuronal development. In particular, the
involvement of the ECM in synapse formation and maintenance as well as in synaptic
plasticity has been reported. Beyond that, perineuronal nets (PNNs), which assemble a
defined accumulation of ECM molecules around subpopulations of neurons, were
reported to be crucial for maintaining synaptic connections. However, the knowledge
about the precise function of the ECM molecules of PNNs in synapse formation and
synaptic transmission remained elusive so far.
To gain insight the role of ECM molecules in synapse formation and synaptic activity we
used an indirect neuron-astrocyte in vitro assay. Therein, primary mouse hippocampal
neurons were co-cultivated with primary cortical mouse astrocytes. The cells had no
direct, membrane mediated contact, but an indirect cell-cell communication was
enabled through sharing the same medium. Therefore, neurons could benefit from
astrocyte-released soluble factors. This allowed for the long-term cultivation of
neurons for up to three weeks and for the detailed investigation of a pure neuronal
culture with separated supporting astrocytes. For studying the consequences of an
ECM deficiency we have utilized a quadruple transgenic mouse model which lacks the
four crucial ECM components tenascin-C, tenascin-R, neurocan, and brevican. Neurons
and astrocytes were derived from the mutant and were used in a combinatorial
approach together with wild-type cells. Thus, four possible combinations were
investigated: both cells from wild-type (Awt/wt|Nwt/wt), astrocytes from the wild-type and
neurons from the mutant (Awt/wt|Nko/ko), the other way around (Ako/ko|Nwt/wt) and both
cells derived from the mutant (Ako/ko|Nko/ko). Using that approach we were able to
separate the neuronal form the astrocytic contributions and to investigate the neuronal
development in detail. First, we have made attempts to confirm the expression of the
aforementioned ECM molecules in the given assay via immunocytochemical stainings.
Thereby we noticed that tenascin-C, tenascin-R, neurocan, and brevican were
expressed in distinct expression patterns using the Awt/wt|Nwt/wt assay. The
combinatorial use of wild-type and mutant cells provided evidence that neurons
- 128 -
Chapter 5
Summary
themselves highly contributed to the ECM assembly under these defined conditions.
After confirming the complete absence of the four ECM molecules in the Ako/ko|Nko/ko
combination we investigated the physiological consequences of the ECM deficiency. Via
whole-cell voltage-clamp recordings in cooperation with the lab of Prof. Dr. Dr. Dr.
Hanns Hatt the primary neurons were characterized electrophysiologically (recordings
were done by Dr. Ainhara Aguado). By quantifying the miniature inhibitory and
excitatory postsynaptic potentials (mIPSCs/mEPSCs) we have found a significant
reduction in the frequency of these spontaneous events after 14 and 21 days in vitro
(DIV) in mutant-derived neurons. Thus, the ECM deficiency during neuronal
development and synapse formation was shown to result in severe changes in the
spontaneous transmitter release frequency. This effect was almost independent from
the genotype of the astroglia added. The amplitude as well as the kinetics remained
unchanged in the investigated combinations. To unravel the reasons for the observed
alterations,
we
investigated
the
expression
of
synaptic
key
proteins.
Via
immunocytochemical detection of PSD-95 and Bassoon we could show, that the
formation of synapses revealed additional modifications in the mutant neurons.
Therefore, we have found an overall trend to an increased synaptic puncta expression
in the mutant-cultures after 14 DIV. This became especially obvious in culture where
both cells were derived from the mutant. After 21 DIV a reverse picture appeared:
despite the developmental increase in the absolute expression of Bassoon and PSD95,
the relative expression of PSD 95 and Bassoon was significantly reduced in the
Ako/ko|Nko/ko combination compared to the Awt/wt|Nwt/wt cells. As mirrored in these
results, the ECM seems to play a dual role during synapse formation in vitro. While the
early ECM can be speculated to foster the formation of synapses, the more mature
ECM may play a crucial role in the maintenance of synaptic contacts. This was nicely
shown by the removal of the ECM molecules in the described assay. With proteinbiochemical approaches we have made first attempts to characterize the inhibitory and
excitatory subpopulations in the cultures. The data revealed a significantly increased
expression of the GABA synthesizing enzymes GAD 65 and 67, while the vesicular
glutamate transporter vGLUT displayed unchanged expressions levels in the matrixdeficient network. In how far these alterations are overlapping with changes in the
- 129 -
Chapter 5
Summary
neuronal subpopulations will be subject of further investigations. However, we could
show that PNNs are severely disrupted in the mutant derived cultures. This could not
be rescued by the addition of wild-type astrocytes.
These data for the first time show that the simultaneously genetic manipulation of four
ECM molecules leads to changes in the formation of PNNs paralleled with physiological
changes and altered synapse formation. These data concerning the impact of the ECM
molecules on synapse formation and synaptic activity were combined to a manuscript
which is presented in chapter 3.
The second part of the present work, included with chapter 4, presents a
methodological paper, recently published in the Journal of Neuroscience Methods. This
publication deals with the transfer of the indirect neuron-astrocyte co-culture assay to
a multi electrode array (MEA) set up. This allows the physiological investigation of the
primary hippocampal mouse cultures on a network-wide level under defined conditions.
In addition, we have undertaken steps in the characterization of the hippocampal
network. The cells were cultivated for up to four weeks and we performed a detailed
developmental investigation of the spontaneous activity. We were able to confirm the
proper development of the cells in the novel MEA assay, via analyzing a variety of
parameters, such as the spike-frequency and the burst characteristics. Additionally, we
investigated the neuronal activity during the antagonisation of the GABAA receptors
with bicuculline.
In summary, the present work highly contributed to the existing knowledge about the
ECM during synapse formation and synaptic transmission in vitro and opened new ways
for analyzing primary hippocampal mouse neurons and astrocytes under defined
conditions on a network-wide level.
- 130 -
Chapter 5
Zusammenfassung
5.3 Zusammenfassung
Die Forschung der vergangen Jahre hat viel zu dem Verständnis der fundamentalen
Bedeutung der extrazellulären Matrix (EZM) während der neuronalen Entwicklung
beigetragen. Insbesondere konnte gezeigt werden, dass die EZM bei der Bildung und
Plastizität von Synapsen eine wesentliche Rolle spielt. Darüber hinaus deuten einige
Arbeiten darauf hin, dass perineuronale Netze (PNNs), welche eine Akkumulation
definierter EZM Moleküle um Subpopulationen von Neuronen bilden, wichtig für die
Stabilisierung von Synapsen sind. Dennoch ist die detaillierte Funktion der EZM
Moleküle in den PNNs bis heute lückenhaft.
Um die Funktion der EZM Moleküle im Rahmen der Synapsen Bildung und der
synaptischen Aktivität zu analysieren, wurde in der vorliegenden Arbeit eine indirekte
Co-Kultur von Neuronen und Astrozyten verwendet. In diesem Ansatz wurden primäre
hippokampale Neurone und primäre kortikale Neurone aus der Maus verwendet. Dabei
hatten die Zellen keinen direkten Kontakt und Zell-Zell Kommunikation fand über das
gemeinsame Zellkulturmedium statt, so dass die Neuronen von löslichen astrozytären
Faktoren profitieren konnten. Das erlaubte die Langzeit-Kultivierung und die detaillierte
Untersuchung der primären Neurone über bis zu drei Wochen unter definierten
Bedingungen und getrennt von Gliazellen.
Um die Konsequenzen einer EZM Defizienz zu untersuchen, haben wir in den
Experimenten ein transgenes Mausmodell verwendet. Dieser quadruple knock-out
Maus fehlen die wichtigen EZM Moleküle Tenascin-C; Tenascin-R, Neurocan und
Brevican. Neurone und Astrozyten wurden aus Gewebe dieses transgenen Modells
gewonnen und mit Zellen aus dem Wildtyp kombiniert. So wurden vier verschiedene
Bedingungen untersucht: Beide Zelltypen aus dem Wildtyp (Awt/wt|Nwt/wt), Astrozyten
aus dem Wildtyp und Neurone aus der Mutante (Awt/wt|Nko/ko), umgekehrt (Ako/ko|Nwt/wt)
und beide Zellen aus der Mutante gewonnen (Ako/ko|Nko/ko). Dieser Ansatz machte es
möglich, die neuronalen und die astrozytären Einflüsse getrennt und die neuronale
Entwicklung im Detail zu untersuchen.
Zunächst konnten wir mittels immunzytochemischer Experimente zeigen, dass die
genannten EZM Bestandteile Tenascin-C; Tenascin-R, Neurocan und Brevican in
- 131 -
Chapter 5
Zusammenfassung
definierten Expressionsmustern in dem vorliegenden Kultursystem exprimiert wurden,
während in der Ako/ko|Nko/ko Kombination keine immunpositiven Zellen nachgewiesen
werden konnten. Durch die kombinatorische Kultivierung von Neuronen und Astrozyten
beider Genotypen konnten wir zeigen, dass der Aufbau der EZM in vitro unter definierten
Bedingungen maßgeblich von den Neuronen beeinflusst wurde.
Mittels voltage-clamp Messungen in Kooperation mit dem Labor von Prof. Dr. Dr. Dr.
Hanns Hatt wurden die Neurone elektrophysiologisch charakterisiert (die Messungen
wurden von Dr. Ainhara Aguado durchgeführt). Durch die Quantifizierung der
inhibitorischen
und
exzitatorischen
postsynaptischen
Miniatur
Potenziale
(mIPSC/mEPSC) konnte eine signifikant reduzierte Frequenz dieser spontanen
Potenziale nach 14 und 21 Tagen in vitro (DIV/ days in vitro) gezeigt werden. Somit
resultiert die EZM Defizienz während der neuronalen Entwicklung und Synapsen
Bildung in deutliche Veränderungen der Häufigkeit spontaner NeurotransmitterFreisetzung. Dieser Effekt war größtenteils unabhängig vom Genotyp der cokultivierten Astrozyten. Die Amplitude sowie die Kinetiken der Kanäle blieben in den
untersuchten Kombinationen unverändert.
Um die Hintergründe dieser physiologischen Veränderungen zu untersuchen, haben wir
die
Expression
von
wichtigen
synaptsichen
Proteinen
untersucht.
Mittels
immunzytochemischer Detektion von PSD-95 und Bassoon konnten wir weitere
Veränderungen in den Neuronen der Mutante zeigen. Wir konnten einen Trend zur stark
erhöhten Expression synaptischer Punkte nach 14 DIV nachweisen. Das war besonders
deutlich in Kulturen, in denen beide Zelltypen aus der knock-out Maus stammten. Nach
21 DIV zeigte sich ein gegenläufiger Effekt: Trotz der entwicklungsbiologisch erhöhten
absoluten Expression von PSD-95 und Bassoon zu diesem Zeitpunkt konnten wir eine
signifikant verringerte relative Expression der synaptischen Proteine in den EZMdefizienten Kulturen zeigen. In diesen Ergebnissen spiegelt sich die duale Rolle der EZM
während der Synapsen Entwicklung in vitro wieder. So wird vermutet, dass die frühe
EZM die Synapsen Bildung fördert, während die reife EZM eher eine wichtige Rolle in
der Stabilisierung von sypatischen Kontakten spielt. Das konnte anhand der Defizite in
der EZM Zusammensetzung in der vorliegenden Arbeit gezeigt werden.
- 132 -
Chapter 5
Zusammenfassung
Mittels proteinbiochemischer Methoden haben wir erste Ansätze etwickelt um die
neuronale Zusammensetzung der Kultur in Bezug auf exzitatorische und inhibitorische
Subpopulationen zu untersuchen. Diese Daten zeigten eine signifikant erhöhte
Expression der GABA-synthetisierenden Enzyme GAD 65 und GAD 67, während die
Expression des vesikulären Gluatamat Transporters vGLUT sich unverändert in den
Kulturen der quadruple knock-out Zellen zeigte. In wieweit diese Veränderungen mit
möglichen Verschiebungen in den neuronalen Subpopulationen übereinstimmen, soll
Gegenstand zukünftiger Untersuchungen sein.
Wir konnten zeigen, dass die PNNs in den EZM-defizienten Kulturen deutliche reduziert
waren. Dieser neuronale Effekt konnte nicht durch die Co-Kultivierung mit Astrozyten
aus dem Wildtyp verhindert werden. Weiterhin konnten wir zeigen, dass die Expression
der untersuchten synaptischen Proteine in den Neuronen mit lediglich rudimentär
ausgebildeten PNNs signifikant reduziert war. Das unterstützt die Annahme, dass die
PNNs eine wesentliche Rolle bei der Bildung und Stabilisierung von Synapsen spielen.
Diese Daten zum Einfluss der EZM Moleküle auf die Synapsen Bildung und die
synaptische Aktivität wurden zu einem Manuskript zusammengefasst und sind in
Kapitel 3 eingebunden.
Der zweite Teil der vorliegenden Arbeit wird in Kapitel 4 präsentiert und beschreibt eine
kürzlich im Journal of Neuroscience Methods publizierte methodische Studie. Diese
Publikation beschreibt die Vewendung der indirekten Neuron-Astrozyten Co-Kultur in
Verbindung mit einem Multi Elektroden Array (MEA). Die Anwendung dieses Ansatzes
macht die physiologische Untersuchung der primären hippokampalen Mausneuronen
auf einem Netzwerk weiten Level unter definierten Bedingungen möglich.
In der vorliegenden Arbeit wurden die Zellen für bis zu vier Wochen kultiviert und eine
detailierte Untersuchung der Entwicklung der Spontanaktivität in der Kultur
vorgenommen. Wir konnten die korrekte Entwicklung der hippokampalen Neurone
durch die Analyse verschiedener Parameter wie der Frequenz von Aktionspotenzialen
und
der
Burst-Charakteristika
bestätigen.
Weiterhin
untersuchten
wir
die
Veränderungen der neuronale Aktivität nach Verwendung des GABAA-RezeptorAntagonisten Bicuculline.
- 133 -
Chapter 5
Zusammenfassung
Zusammenfassend hat die vorliegende Arbeit zu dem bestehenden Wissen über die
Rolle
der
EZM während
der
Synapsen Bildung und
in der
synaptischen
Signalübertragung beigetragen und eröffnet neue Wege zur Analyse von Netzwerken
primärer hippokampalen Mausneuronen unter definierten Bedingungen in indirekter CoKultur mit Astrozyten.
- 134 -
Chapter 5
List of abbreviations
5.4 List of Abbreviations
Abbreviations
µm
µM
A
ACM
AKAP
Awt/wt|Nko/ko
Awt/wt|Nwt/wt
Ako/ko|Nko/ko
Ako/ko|Nwt/wt
AMPA
AMPAR
ANOVA
APOER2
AraC
BDNF
BIC
Bral2
Ca2+
CaCl2
CAM
ChABC
CNR
CNS
CO2
CRP
Crtl1
CS
CSPG
DABI
DIV
DMEM
DNQX
DS
DSPG
E
ECM
EDTA
micrometer
micromolar
genotype of astrocytes
astrocyte conditioned medium
A-kinase anchoring protein
Astrocytes derived from wild-type; neurons from the knockout
Astrocytes and neurons derived from wild-type
Astrocytes and neurons derived from knock-out
Astrocytes derived from knock-out; neurons der. from wildtype
3-hydroxyl-5-methyl-4-isoxazole-propionate
AMAP receptor
analysis of variance
apolipoprotein E receptor type 2
cytosine-1-ß-D arabinofuranosid
derived neurotrophic factor
bicuculline
brain-specific link protein 2
calcium
calcium chloride
cell adhesion molecule
Chrondroitinase ABC
cadherin-related neuronal receptor
Central nervous system
carbon dioxide
complement regulatory protein
cartilage link protein 1
Chondroitinsulfate
Chrondroitin sulfate proteoglycans
disabled 1
days in vitro
dulbecco´s modified eagle medium
6,7-dinitroquinoxaline-2,3-dione
Dermatansulfate
Dermatan sulfate proteoglycans
embryonal day
extracellular matrix
ethylenediaminetetraacetic acid
- 135 -
Chapter 5
EGF
EGTA
FEP
FGF
Fig.
FNIII
GABA
GAD
GAG
GFAP
GKAP
GRIPs
h
HA
HEPES
HIV
HNK-1
HRP
HRP
HS
HSPG
Hz
Ig
KCl
kDA
ko
KS
KSPG
LTD
LTP
LVDCC
MAGUK
MCS
MEA
MEM
mEPSC
MgCl2
mIPSC
ml
mOsm
mPSC
ms
List of abbreviations
epidermal growth factor
ethylene glycol tetraacetic acid
fluorinated ethylene–propylene
fibroblast growth factor
figure
fibronectin type III
gamma-aminobutyric acid
glutamate decarboxylase
glycosaminoglycan
Glial fibrillary acidic protein
guanylate kinase-associated protein
glutamate receptor interacting proteins
hour
Hyaluronan
4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid
hours in vitro
human natural killer-1 carbohydrate
horseradish peroxidase
horseradish peroxidase
Heparansulfate
Heparan sulfate proteoglycans
hertz
Immunoglobin
potassium chloride
kilo dalton
knock out
Keratansulfate
Keratan sulfate proteoglycans
long term depression
long term potentiation
L-type voltage-dependent Ca2+ channels
membrane-associated guanylate kinase
Multi Channel Systems GmbH
multi electrode array
Minimum Essential Media
miniature excitatory postsynaptic current
Magnesium chloride
miniature inhibitory postsynaptic current
milliliter
milliosmole
miniature postsynaptic current
millisecond
- 136 -
Chapter 5
N
narp
NF
NG2
nm
NMDA
NMDAR
NMJ
NT
OPCs
P
PBS
PFA
PI
PNN
PSD
PSD95
PTV
PTX
PVDF
s
SDS
SEM
Sema3A
SFK
STV
SV
SVZ
SynCAM
trk
TSP
TTX
UV
v/v
vGLUT
VLDLR
VZ
w/v
WFA
wnt
wt
List of abbreviations
genotype of neurons
neural activity related protein
Neurofilament
neuron glia antigen 2
nanometer
N-Methyl-D-aspartate
NMDA receptor
neuromuscular junction
neurotrophin
oligodendrocyte precursor cells
postnatal day
phosphate buffered saline
paraformaldehyde
propidiumiodid
perineuronal net
postsynaptic density
postsynaptic density protein of 95 kDA
piccolo transport vesicles
Picrotoxin
polyvinylidene fluoride
second
sodium dodecyl sulfate
standard error of the mean
semaphorin 3 A
Src family tyrosine kinases
synaptic vesicle protein transport vesicles
synaptic vesicle
subventricular zone
synaptic cell adhesion molecule
tropomyosin receptor kinase
thrombospondin
Tetrodotoxin
ultraviolett
volume per volume
vesicular glutamate transporter
very-low-density lipoprotein receptor
ventricular zone
weight per volume
Wisteria floribunda agglutinin
wingless gene
wild type
- 137 -
Chapter 6
Erklärung
Chapter 6
Appendix
6.1 Erklärung
Hiermit erkläre ich, dass ich die Arbeit selbstständig verfasst und bei keiner anderen
Fakultät eingereicht und dass ich keine anderen als die angegebenen Hilfsmittel
verwendet habe. Es handelt sich bei der heute von mir eingereichten Dissertation
um sechs in Wort und Bild völlig übereinstimmende Exemplare.
Weiterhin erkläre ich, dass digitale Abbildungen nur die originalen Daten enthalten
und in keinem Fall inhaltsverändernde Bildbearbeitung vorgenommen wurde.
Bochum, den 01.02.2012
____________________
Maren Geißler
- 138 -
Chapter 6
Curriculum Vitae
6.2 Curriculum Vitae
Name:
Maren Geißler
Address:
Graf-Engelbert-Strasse 16
40489 Düsseldorf
Date of birth:
November, 16th, 1981
Place of birth:
Essen
Academic career:
Since 10/2007
Research fellow at the Ruhr-University
Bochum at the Department of Cell
Morphology
and
Molecular
Neurobiology
10/2007-02/2012
PhD student at the International
Graduate School for Bioscience at the
Ruhr-University
Bochum
at
the
Department of Cell Morphology and
Molecular Neurobiology under the
supervision of Andreas Faissner.
Title: Extracellular matrix molecules of
perineuronal nets – Studies on
structure and function in synapse
formation and synaptic activity
10/2007-02/2012
Member of the research school of the
Ruhr-University Bochum (GSC98/1)
- 139 -
Chapter 6
Curriculum Vitae
10/2007-10/2010
Member of the DFG GRK 736,
Entwicklung und Plastizität des
Nervensystems:
Molekulare, synaptische und zelluläre
Mechanismen
04/2007-09/2007
PhD student at the institute of
anatomy/ Universitäts Klinikum Essen
within
the
DGF
GRK
1431
“Transcription, chromatin structure and
DNA repair in development and
differentiation”
Title:
“Transcriptional control of apoptotic
processes in the developing retina”
04/2006-04/2007
Research fellow and diploma student
the Ruhr-University Bochum at the
Department of Theoretical Biology/ Lab
for neuroinformatics/neural plasticity
lab under the supervision of PD. Dr.
Huber Dinse
Title:
Mononukleäre Zellen aus humanem
Nabelschnurblut
lindern
die
Auswirkungen
eines
perinatalen
hypoxisch-ischämischen Insults bei
Ratten.
Eine
elektrophysiologische
Untersuchung
10/2001-04/2007
Studies in Biology
University Bochum
- 140 -
at
the
Ruhr-
Chapter 6
Publications and benchmark of contribution
6.3 Publications and benchmark of contribution
1)
Maren Geissler, Ainhara Aguado, Uwe Rauch, Christian Wetzel, Hans Hatt,
Andreas Faissner
“Primary hippocampal neurons, lacking four ECM molecules, show severe
deficits in PNN formation and abnormalities in synaptic structure and function”
Manuscript in preparation
Planning the experimental set-up: 60 %
Performing experiments: 80 %
Writing the manuscript: 80 %
2)
M. Geissler and A. Faissner
“A new indirect co-culture set up of mouse hippocampal neurons and astrocytes
on microelectrode arrays”
Journal of neuroscience methods 2011 (accepted)
Planning the experimental set-up: 90 %
Performing experiments: 100 %
Writing the manuscript: 90 %
3)
M. Geissler, H. R. Dinse, S. Neuhoff, K. Kreikemeier and C. Meier (2011). "Human
umbilical cord blood cells restore brain damage induced changes in rat
somatosensory cortex." PLoS One 6(6): e20194., 2011
Planning the experimental set-up: 50 %
Performing experiments: 90 %
Writing the manuscript: 33 %
- 141 -
Chapter 6
4)
Publications and benchmark of contribution
M. Pyka, C. Wetzel, A. Aguado, M. Geissler, H. Hatt and A. Faissner
Chondroitin sulfate proteoglycans regulate astrocyte-dependent synaptogenesis
and modulate synaptic activity in primary embryonic hippocampal neurons, Eur J
Neurosci. 2011
Planning the experimental set-up: 5 %
Performing experiments: 5 %
Writing the manuscript: 5 %
5)
A. Faissner, M. Pyka, M. Geissler, T. Sobik, R. Frischknecht, E. D. Gundelfinger
and C. Seidenbecher
Contributions of astrocytes to synapse formation and maturation - Potential
functions of the perisynaptic extracellular matrix. Brain Res Rev 63, 26-38, 2010
Planning the experimental set-up: 0 %
Performing experiments: 0 %
Writing the manuscript: 10%
- 142 -
Chapter 6
Publications and benchmark of contribution
6.4 Conference participations and poster abstracts
1)
M.Geissler, K.Kreikemeier, S.Neuhoff, C.Meier, H.Dinse (2007)
7th meeting of the German Neuroscience Society (Göttingen, March 2007)
Stem cell induced cortical plasticity reduces brain damage after perinatal
asphyxia in rats
2)
M. Geissler, C. Meier , K.Kreikemeier, S. Neuhoff, H.R. Dinse (2007)
Society for Neuroscience (SfN) 37th Annual Meeting (San Diego, November
2007)
Human umbilical cord blood cells restore cortical maps and cortical excitability
after hypoxic ischemia in rats
3)
M.Geissler and A. Faissner (2008)
2nd GRK Meeting (Bochum 2008)
Contribution of extracellular matrix (ECM) molecules to synaptogenesis and
synaptic plasticity: studies in the quadruple knock out mice
4)
M.Geissler and A. Faissner (2009)
8th meeting of the German Neuroscience Society (Göttingen, March 2009)
Contribution of ECM molecules to synaptogenesis and synaptic plasticity: studies
in the quadruple knock out mice
5)
M.Geissler and A. Faissner (2009)
2nd Section day research school Bochum 2009:
Contribution of the extracellular matrix (ECM) and their perineuronal nets (PNNs)
to synaptogenesis and synaptic plasticity: studies in the quadruple knock out mice
6)
M. Geissler and A. Faissner (2010)
7th Forum of European neuroscience (FENS) (Amsterdam July 2010)
Contribution of the extracellular matrix (ECM) and their perineuronal nets (PNNs)
to synaptogenesis and synaptic plasticity: studies in the quadruple knock out mice
- 143 -
Chapter 6
Danksagung
6.5 Danksagung
Größter Dank gilt meinem Doktorvater Prof. Dr. Andreas Faissner, der mir ermöglicht
hat, meine Promotion am Lehrstuhl für Zellmorphologie und molekulare Neurobiologie
durchzuführen. Ich danke Ihm für das entgegengebrachte Vertrauen in meine Arbeit, für
die fortwährende Unterstützung, die stetige Motivation und die Anerkennung meiner
Leistungen.
Weiterhin danke ich Prof. Dr. Dr. Dr. Hanns Hatt für die Übernahme des
Zweitgutachtens, sowie für die erfolgreiche Kooperation zwischen den zwei Lehrstühlen
über die gesamte Promotionszeit hinweg.
Wesentlich zum Erfolg meiner Doktorarbeit sowie zu meiner wissenschaftlichen
Weiterentwicklung und zur Erweiterung meines neurobiologischen Horizontes, hat
meine Ausbildung innerhalb des DFG Graduiertenkollegs 736 unter der Leitung von
Prof. Dr. Petra Wahle beigetragen. Bei Ihr, dem GRK Kollegium sowie bei den GRK
Mitgliedern möchte ich mich ganz herzlich bedanken.
Den GRK Doktoranden Janine Neumann, Bettina Bertram, Kathrin Engelhart, Ben
Novak und Arne Buschler danke ich für diese großartige Zeit!
Innerhalb des Lehrstuhls gibt es viele Menschen, denen ich für Ihre Begleitung durch
diese nicht immer leichte Zeit danken möchte. Das sind ganz besonders Martin Pyka,
Anke Mommsen und Simon van Leeuwen, ohne die diese Zeit ein ganzes Stück
trostloser gewesen wäre. Aber auch Michael Karus, Eva Hennen, Jacky Reinhardt,
Thomas Sobik, Tim Czopka, Melanie Michele, Stefanie Hahn, Sören Moritz, Alexander
von Holst, Bettina Göldner, Sabine Kindermann, Mario Völzkow, Stefan Wiese und allen
anderen lieben Kollegen und Ehemaligen gilt mein Dank für Ihre Unterstützung und die
hilfreichen Tipps. Den von mir betreuten Studenten Gianna Springer, Christine
Gottschling, Ioanna Ioannidou
und Alina Blusch danke ich für die problemlose
Zusammenarbeit, Ihr Vertrauen und die tolle Arbeit! Alle vier haben wesentlich zum
Bestehen der „Synapsen“ beigetragen.
- 144 -
Chapter 6
Danksagung
Viele erfolgreiche Kooperationen haben meine Arbeit zu dem gemacht was sie
geworden ist. Daher gilt mein Dank Prof. Dr. Georg Juckel, PD Dr. Rainer Wolf, Jennifer
Plümper und Dr. Marie Pierre Manitz. Weiterhin danke ich Prof. Dr. Denise ManahanVaughan, Arne Buschler und Stefan Jansen. An dieser Stelle bedanke ich mich auch bei
PD. Dr. Christian Wetzel und Dr. rer. nat. Ainhara Aguado für diese gehaltvolle,
produktive und reibungslose Kooperation. Für die Unterstützung bei der erfolgreichen
Etablierung des MEA Set UPs danke ich der Firma Multi Channels Systems,
insbesondere Frank Hofmann und Mirco Schanz.
Zwei besondere Kommilitoninnen haben mich vom Anfang des Studiums bis zum Ende
der Promotion an der Uni sowie im Privaten begleitet und werden es hoffentlich auch
darüber hinaus noch tun. Meinen herzlichen Dank an Ina Wilms und Janina Kreuz!
An dieser Stelle möchte ich mich auch bei meinen liebsten Freunden bedanken, dass
sie mich durch die Höhen und Tiefen während der Jahre der Promotion begleitet haben,
immer für mich da waren und für den nötigen Ausgleich gesorgt haben. Dank gilt hier
besonders Carolin Butterfield, Angelika Görlich, Jessica Wagener und Melanie Leib!
Auf familiärer Ebene danke ich meiner Mutter, Klaus und Oma Hetti für Ihre tatkräftige
finanzielle aber vor allem mentale Unterstützung während meines gesamten Studiums.
Vor allem aber danke ich Ihnen für das große Interesse an meiner Arbeit und Ihren
Glauben an mich! Ohne Euch wäre ich nicht so stark geworden, das Alles zu packen!
Danke dafür und, dass Ihr so stolz auf mich seid! Dafür danke ich auch meinem Vater,
Sigi, Opa und Oma Toni! Danke, dass Ihr all die Jahre so hinter mir gestanden habt. Es
tut gut eine starke Familie im Rücken zu haben!
Dank gilt auch meiner „neuen Familie“: Danke an Opa, Werner und Nina!
Am meisten abbekommen hat jedoch der Mensch, der tagtäglich meine Launen,
Ängste, Sorgen und Zweifel angenommen hat! Tim, ich danke Dir dafür, sowie für Dein
fortwährendes Aufbauen, Aufmuntern, Unterstützen, für Deine Zuversicht, Deinen
Glauben an mich und für Deine Liebe!
- 145 -