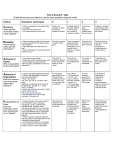
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
CSIRO PUBLISHING Wildlife Research, 2009, 36, 81–97 www.publish.csiro.au/journals/wr Use of artificial tree hollows by Australian birds and bats Ross L. Goldingay A,B and Jane R. Stevens A A School of Environmental Science and Management, Southern Cross University, PO Box 157, Lismore, NSW 2480, Australia. B Corresponding author. Email: [email protected] Abstract. Artificial tree hollows (nest or roost boxes) may be of considerable importance to the conservation and management of Australian hollow-using birds and microbats. This is suggested by recognition that the rate of collapse of hollow-bearing trees may exceed replacement in some landscapes. We review the published literature to synthesise current information on the use of artificial hollows by Australian birds and bats, and to provide guidance to future research and management. The use of artificial hollows has been documented in some detail for 15 native bird and eight microbat species. A range of hollow designs has been employed but there is a limited understanding of favoured designs. General designs (e.g. front-entry plywood boxes) have been used extensively by some species and should continue to be used until more effective designs are identified. Species tend to use artificial hollows that have entrance sizes just sufficient for their body size, and this should guide hollow design. Competitive interactions with a range of non-target species (native and non-native) may have a pronounced influence on artificial hollow use and must be considered in any management program involving artificial hollows. We highlight some design elements that may reduce interference by non-target species. Temperature inside artificial hollows may have a particular influence on their use by bats due to the role of microclimate in bat thermoregulation. Trials are needed to investigate this factor and to inform general approaches to positioning of artificial hollows. Several distinct management uses exist for artificial hollows, including assisting threatened species recovery, e.g. Kangaroo Island glossy black-cockatoo (Calyptorhynchus lathami halmaturinus) and orange-bellied parrot (Neophema chrysogaster). Artificial hollows offer an interim solution to hollow shortage but their full potential will only be realised when preferences for different designs are better understood. This will require a commitment to monitoring and should be conducted in an adaptive management context. Increased knowledge of the use of artificial hollows by Australian birds and bats should be of global relevance to the management of hollow-using species because it provides an independent test of ideas and strengthens any generalisations. Introduction Species of wildlife throughout the world use tree hollows for protection when inactive and as breeding sites (e.g. Saunders et al. 1982; Mackowski 1984; Newton 1994; Nowak 1999; Kunz and Lumsden 2003; Zinner et al. 2003). In Australia, as many as 300 native vertebrate species (birds, bats, arboreal marsupials, reptiles and frogs) use tree hollows (Gibbons and Lindenmayer 2002). This corresponds to a far greater proportion of wildlife species dependent on tree hollows for survival compared to that on other continents (see Saunders et al. 1982; Newton 1994; Gibbons and Lindenmayer 2002). This prevalence of hollow dependence among Australian wildlife should result in land managers and researchers leading the way internationally in understanding the hollow requirements of species and devising effective management programs for them. At present, this does not appear to be the case with widespread concern that the abundance of tree hollows has declined within many landscapes in Australia (Joseph et al. 1991; Bennett et al. 1994; Mawson and Long 1994; Lindenmayer et al. 1997; Pell and Tidemann 1997a; Gibbons et al. 2002; Heinsohn et al. 2003; Eyre 2005; Harper et al. 2005a; Courtney and Debus 2006). As a consequence, many hollow-dependent species are now listed CSIRO 2009 as threatened (Gibbons and Lindenmayer 2002). Management programs in response to this appear to be in their infancy. Artificial hollows (nest and roost boxes) can potentially simulate the tree hollow environment for many hollow-using species (Menkhorst 1984; Boyd and Stebbings 1989; Petty et al. 1994; Smith and Agnew 2002; Harley 2004; Beyer and Goldingay 2006; Goldingay et al. 2007) and this is well recognised in Europe and North America (Stebbings and Walsh 1991; Newton 1994). Indeed, understanding factors that may influence the use of artificial hollows is an active area of research in Europe and North America for birds (e.g. Pogue and Schnell 1994; Ardia et al. 2006) and bats (e.g. Kerth et al. 2001; Flaquer et al. 2006). Artificial hollows have long been recognised as an important research and management tool for Australian arboreal marsupials (Menkhorst 1984; Beyer and Goldingay 2006). In contrast, the use of artificial hollows by Australian birds is poorly documented in the published literature despite the deployment of artificial hollows as a management tool for over a decade (e.g. Olsen 1996; Mooney and Pedler 2005). There is increasing evidence that roost boxes will be used by Australian microbats (Golding 1979; Irvine and Bender 1995; Ward 2000; Smith and Agnew 2002) but the extent to which this can result in 10.1071/WR08064 1035-3712/09/020081 Wildlife Research valuable research and management applications is not well understood. This paper reviews published information on the use of artificial hollows by Australian birds and bats. The large number of hollow-using species in Australia means that any increase in knowledge and understanding can benefit the management of a significant component of biodiversity. Specifically we aim to: (i) synthesise available published information on the use of artificial hollows; (ii) examine different applications of artificial hollows; and (iii) identify gaps in our knowledge so as to stimulate future research. The present review complements one by Beyer and Goldingay (2006) concerning the use of artificial hollows by Australian arboreal marsupials. We make extensive reference to studies in the northern hemisphere because there is a substantial literature for that area that can greatly inform our understanding of the requirements and behaviour of Australian species. Furthermore, an increased understanding of Australian species can provide independent tests of various hypotheses about the use of artificial hollows, which will allow greater generalisation for any hollow-using birds and bats. Use of artificial hollows by Australian birds and microbats To review the literature on the use of artificial hollows by Australian birds and bats, we conducted searches of the Web of Science as well as the Australian journals Australian Journal of Zoology, Emu and Wildlife Research, using webpage search tools and keywords relating to artificial hollows. The journal Australian Mammalogy was searched manually. Theses were only included when commonly referred to by published studies. Some studies and most books on this topic have documented species observed using artificial hollows but have provided few details of such use. We restrict our attention to studies that describe more than one observation of nest and roost box use by birds or bats, and in which some details of the artificial hollows used were described. Where possible we have collated data on all species referred to in studies, regardless of whether one species or wildlife group was targeted. Only 27 studies have been published during the last 32 years on the use of nest boxes by birds, with 14 published during 1996–2000 (Fig. 1). This included 15 native and two introduced species. Nest box use has been described for seven parrot species, three passerines, one nocturnal bird and two waterfowl species. The use of artificial hollows by microbats has fared much more poorly with just five studies covering eight species published in the last 32 years (Fig. 1). The most commonly encountered species in artificial hollows, in terms of the number of individuals or boxes occupied and the number of studies in which they featured, were the crimson rosella (Platycercus elegans), chestnut teal (Anas castanea), common myna (Acridotheres tristis), Gould’s wattled bat (Chalinolobus gouldi) and the large forest bat (Vespadelus darlingtoni) (Table 1). Welcome swallows (Hirundo neoxena) used at least 800 plastic drum nest boxes in Western Australia but no further details were provided (Norman and Riggert 1977). The chestnut teal (Anas castanea) accounted for 363 boxes occupied in Victoria (Norman and Riggert 1977). This and one other study (Briggs 1991) are the R. L. Goldingay and J. Stevens 16 Number of studies 82 14 Microbats 12 Birds-Management 10 Birds-Research 8 6 4 2 0 1976–80 1981–85 1986–90 1991–95 1996–00 2001–05 2006+ Year of publication Fig. 1. The number of studies on the use of artificial hollows by birds and microbats published in 5-year periods since 1976. only ones that have targeted waterfowl despite the use of tree hollows for nesting by 10 Australian species (Saunders et al. 1982; Marchant and Higgins 1990) and the common use of nest boxes to study waterfowl in the USA (see Eadie et al. 1998; Evans et al. 2002). Sample sizes in Australian studies exceeded 50 (either boxes occupied or individuals encountered) for just six species (Table 1), though some studies did not document the number of individuals. The endangered orange-bellied parrot (Neophema chrysogaster) and Kangaroo Island glossy blackcockatoo (Calyptorhynchus lathami halmaturinus) have used nest boxes repeatedly but the details of this are not well documented (Mooney and Pedler 2005; Orange-bellied Parrot Recovery Team 2006). Factors that affect the use of artificial hollows Several factors will influence the use of nest and roost boxes. Of particular importance are hollow design and placement, natural hollow availability, and competition with other species. Breeding patterns will produce seasonal patterns of use, and these may differ across taxa. Temperature is also a key factor and may produce a seasonal response in the timing of use. These factors have not been well studied and represent significant gaps in our understanding of artificial hollow use. Hollow design and placement The main design elements that may influence the use of artificial hollows are entrance size, hollow volume, hollow depth below entrance and wall thickness. A great amount of general literature is available recommending detailed box designs for birds and bats (McCulloch and Thomas 1986; Stebbings and Walsh 1991; Grant 1997; Franks and Franks 2003). However, many recommendations appear to have been developed in an ad hoc way. Understanding whether species show preferences for particular design elements and placement positions is central to all research and management applications of artificial hollows. Despite this only a few published studies have provided a choice of hollow types or design elements for birds and bats in Australia (see section Research applications). In the absence of preference studies (e.g. Lumsden 1989; Radunzel et al. 1997) it is likely that future studies will simply follow designs used elsewhere and be unable to state whether low – – XX X – X X X X – – 24 24 Volume (m3) Small (<0.005) Medium (0.005–0.03) Large (>0.03) Height above ground <2 m 2–3.9 m 4–6 m >6 m Aspect North South East West Sample sizes Individuals Boxes occupied 3, 4, 5, 8 – XX – Entrance location Slit below Front/top Spout References – X XX X X 24 Australian owlet-nightjar Entrance diameter <3 cm 3–4.9 cm 5–8 cm 8.1–12 >12 Body size – birds (cm), bats (g) Box variable 10 >2 >2 – – – – – – – – – – X – – – – – – – – 3, 4, 5, 8 21 21 X X – – – – X X – – XX – XX – – – XX – – 15 Norfolk Island boobook owl White-throated treecreeper 30 Grey shrike-thrush 5 – 3 X X – – X – X X – – X – X – – – – – X 24 Striated pardalote 2 4 4 – – – – – XX – – XX – – – XX – – XX – – – 12 Crimson rosella Eastern rosella – 8 X – X – X – X – – X – – X – – – X X – 30 – 6 X – – – – – X – – X – – X – – – – X – 28 Rainbow lorikeet 3, 4, 8, 12, 17 5, 14, 15 12, 13 >85 >95 X X – – X – XX X – XX XX – XX XX – – XX X – 35 Red-rumped parrot 8 – 4 – – X – – – – – – X – – X – – – – – – 27 Glossy black-cockatoo 11 – 41 – – – – – – – XX – – XX – XX – – – – – XX 48 Red-tailed black-cockatoo 9 5 5 – – – – – – – X – – XX – – XX – – – – XX 63 Galah 8, 18 – 2 X – X – – – X – – X – – X – – – – X – 36 Orange-bellied parrot 19 >30 28 – – – – – – – – – X – – X – – X – – – 21 Chestnut teal 1 >360 >360 – – – – XX XX – – – XX XX – X – – – – – XX 48 Turquoise parrot 6 2 2 – – – – X X – – – X – – – X – – X – – 20 Common starling Common myna – 81 XX XX XX – – – XX – – XX – – XX – – – XX X – 25 >250 30 X X – X – X X X X XX X XX X – XX – X – – 12 Gould’s wattled bat 12, 14, 12, 15, 3, 4, 7 15 18 – 12 – X – – – – X – – XX – – X – – – X – – 22 Gould’s long-eared bat 17, 20 50 24 X X X X – X X – X X – XX – – XX – – – – 9 Eastern false pipistrelle Lesser long-eared bat 56 – – – – – – – X X – X X – X – X – X – – 7 Large forest bat >100 10 – – – – – – X X X X X X X – X – – – – 7 >2 10 – – – – – – X X X X X X X – X – – – – 5 Southern forest bat 3, 4, 3, 4, 7, 16 7, 16 16 7, 16 – 3 – – – – – – X X – – X X X – X – – – – 21 Chocolate wattled bat 7 >1 – – – – – – – X – X X – X X – X – – – – 8 7 >5 – – – – – – – X – X X – X – – X – – – – 38 White-striped freetail bat Table 1. Characteristics of artificial hollows used by different species of bird and bat Frequency of use of variable categories: XX = frequently used (>20% of boxes used at a site or in a study); X = known to have been used; – has not been used or absent from studies. Not all variable categories were present where species used nest boxes. Sample sizes have been pooled across studies. Some studies may not give precise data so ‘>’ is used to indicate the true value exceeds that shown. Species body sizes, Simpson and Day (1993), Churchill (1998). References: 1, Norman and Riggert (1977); 2, Milledge (1978); 3, Golding (1979); 4, Calder et al. (1983); 5, Menkhorst (1984); 6, Quin and Baker-Gabb (1993); 7, Irvine and Bender (1995), R. Bender and R. Irvine (unpubl. data), R. Bender (unpubl. data); 8, Trainor (1995a, 1995b); 9, Emison (1996); 10, Olsen (1996); 11, Pedler (1996); 12, Pell and Tidemann (1997a, 1997b); 13, Krebs (1998, 1999), Krebs et al. (1999); 14, Gleeson (1999); 15, Homan (1999); 16, Ward (2000); 17, Smith and Agnew (2002); 18, Harper et al. (2005b); 19, Orange-bellied Parrot Recovery Team (2006); 20, Goldingay (2007) Use of artificial tree hollows by Australian birds and bats Wildlife Research 83 84 Wildlife Research usage was due to the design or local habitat factors. Studies reporting use of artificial hollows by bats are a good example of this. Golding (1979) used 22 sawn-timber boxes and 143 adapted log hollows. Both designs were of similar volume and had a 6.5 cm diameter circular front entrance. He reported use of 23 boxes by 260 bats of five species but did not report whether one design was favoured. Menkhorst (1984) reported no use by bats of 240 front-entry boxes with circular entrances of 5–15 cm diameter. Irvine and Bender (1995) recorded 34 bats in 5 of 10 boxes with a slit-entry at the base. Ward (2000) used a small box with a slit-entry under the lid and reported 73 captures of four species of bats. Smith and Agnew (2002) found that 17 of 48 wedge-shaped boxes with a basal slit-entry were used by bats at two sites. It is impossible to determine from these studies spanning a broad latitudinal range whether one design or design element (e.g. slit v. circular entry) might be favoured. Given the influence of temperature on roost selection by bats (see below), it is also likely that box temperature has played a role in these results. We have summarised information on the range in box design elements used by bird and bat species (Table 1), which provides a broad indication of preference in the absence of direct testing. The materials that boxes are made from have varied substantially among studies. Hollows have been made from wooden ammunition boxes and plastic drums for use by waterfowl (Norman and Riggert 1977), salvaged tree hollows placed in trees for turquoise parrots (Quin and Baker-Gabb 1993) or in trees and on powerpoles for red-tailed black-cockatoos (Calyptorhynchus banksii graptogyne) (Emison 1996; Fig. 2), polyvinyl chloride (PVC) pipe for use by glossy black-cockatoos on Kangaroo Island (Pedler 1996) (Fig. 3), and plywood boxes for eastern rosellas (Fig. 4) and crimson rosellas (Krebs 1998) and for bats (Fig. 5). In Britain, plastic drums have been used extensively by barn owls (Petty et al. 1994). Bat boxes have been made from marine ply (Smith and Agnew 2002) or from pine (Irvine and Bender 1995). No detailed studies have been conducted in Australia to examine the influence of roost box design on bat occupancy. A variety of bat box designs have been used overseas, and often in the context of providing alternative roost sites to bats displaced from roof spaces (Neilson and Fenton 1994). Brittingham and Williams (2000) tested a vertical and a horizontal box of the same dimensions and found a preference for the horizontal box. They also suggested that boxes needed to experience high internal temperatures. Flaquer et al. (2006) compared single- and doublecompartment bat boxes installed on houses, posts and trees. They found greater use of the double-compartment box, and boxes attached to houses and posts, and recorded the highest occupancy by bats of any studies using bat boxes. Their single-compartment box was similar to that used by Irvine and Bender (1995) but with a 1.5-cm rather than a 3-cm slit opening at the base. Entrance size is a key design element, possibly enabling boxes to be designed to target species and it is one element that is easily varied. Most species generally prefer a minimum entrance size to allow access (i.e. one close to body width) but avoid predation by larger species. Menkhorst (1984) found that crimson rosellas preferred one entrance size (8 cm) compared to three others. Several studies found that the Australian owlet-nightjar (Aegotheles cristatus) occupied a range of entrance diameters, R. L. Goldingay and J. Stevens (a) (b) Fig. 2. (a) An artificial hollow installed for red-tailed black-cockatoos in western Victoria. Metal flashing was attached near the base of the pole to exclude access by brushtail possums. (b) This artificial hollow consisted of a salvaged tree hollow attached to a powerpole at a height of approximately 12 m. Photos: R. Goldingay. though it was more frequently encountered in boxes with entrances of 5–8 cm diameter (Table 1) (Fig. 6). Entrance size may be important to exclude some predatory species. Krebs (1998) implicated pied currawongs (Strepera graculina) in high levels of predation on crimson rosella chicks that ceased when she used a 10 cm-long PVC tube to create a spout entrance. Most studies (Table 1) have found that bats generally use small (<3 cm) slit openings located at the bottom (Smith and Agnew 2002; Goldingay et al. 2007; R. Bender, unpubl. data) or top (Ward 2000) of boxes. Large entrances (suitable for larger mammals) are thought to deter bats from nest boxes (Menkhorst 1984), though Golding (1979) reported the use of 23 boxes with a 6.5 cm diameter circular opening by bats. Boxes with slit entrances >20 mm were found occupied by competing larger mammals such as the sugar glider (Petaurus breviceps), introduced black rat (Rattus rattus) and common ring-tailed possum (Pseudocheirus peregrinus) (R. Bender and R. Irvine, unpubl. data) or by feathertail gliders (Acrobates pygmaeus), sugar gliders and squirrel gliders (Petaurus norfolcensis) (Smith and Agnew 2002; Goldingay et al. 2007). Use of artificial tree hollows by Australian birds and bats (a) Wildlife Research 85 (b) Fig. 3. (a) A polyvinyl chloride artificial hollow installed for glossy black-cockatoos on Kangaroo Island. (b) The same design of artificial hollow (~90 cm tall, 30 cm diameter; entrance 16 20 cm) installed for red-tailed black-cockatoos in western Victoria on a powerpole. Photos: (a) E. Sobey; (b) R. Goldingay. Fig. 4. Nestling eastern rosellas in a plywood nest box. Photo: R. Goldingay. Volume and depth are thought to influence the suitability of artificial hollows to birds and bats (Trainor 1995a) but information on this is limited. These parameters of natural hollows influence selection of hollow sites (Saunders et al. 1982; Gibbons et al. 2002), and are known to affect the breeding success of birds (Newton 1994). Nest box depth may provide security from predators (Trainor 1995a). The insulating ability of nest boxes is determined by the type and thickness of materials used. Insulation is thought to affect nest box use (Calder et al. 1983; Menkhorst 1984), leading to natural hollows being favoured when available. Menkhorst (1984) postulated this to explain the relative absence of nest box use by the Australian owlet-nightjar and white-throated tree-creeper (Cormobates leucophaea) at one site where natural hollows were abundant compared to another with low abundance where boxes were used. Trainor (1995a) hypothesised that low insulating properties may affect the breeding success of birds. Common mynas showed no preference for insulating properties (25-mm rough-sawn pine or 12-mm plywood) in one study (Harper et al. 2005b). The insulating ability of nest boxes may be significant to the requirements of bats, which are thought to use hollows that facilitate thermoregulation (Gibbons and Lindenmayer 2002; Lourenço and Palmeirim 2004). R. Bender (unpubl. data) suggested no greater use of 10 roost boxes with a thick wall (45 mm) designed for winter roosting by bats compared to other boxes (19 mm), though the extent to which this changed the microclimate inside these boxes is unknown. Aspect is likely to influence temperature changes within nest and roost boxes (Stebbings and Walsh 1991; Brittingham and Williams 2000; Ardia et al. 2006). Its influence may differ between birds and bats due to the thermoregulatory requirements of bats (see below). No preference for nest box aspect has been detected in any Australian study (Table 1), though few species have been assessed and sample sizes have been small (Calder et al. 1983; Menkhorst 1984; Smith and Agnew 2002). Furthermore, the actual position of boxes on trees may not have provided an adequate test of aspect (Menkhorst 1984). Stebbings and Walsh 86 Wildlife Research R. L. Goldingay and J. Stevens (a) (b) Fig. 5. (a) A wedge-shaped artificial hollow for bats (26 20 18 cm; 1.5-cm slit-entry). (b) Gould’s wattled bats in a wedgeshaped bat box. Photos: (a) R. Goldingay; (b) M. Grimson. (a) (b) Fig. 6. (a) An artificial hollow designed for rosellas (45 20 15 cm; 6.5-cm entrance). No research has addressed the need for perches on these hollows. (b) An owlet-nightjar roosting in this artificial hollow. Photos: R. Goldingay. (1991) recommended choosing aspects for bat boxes that allow the sun to fall directly on the box for part of the day, as well as providing boxes with various aspects in case overheating should occur. Preference trials are needed to resolve this issue. Placement should also consider the surrounding habitat because some species may be influenced by habitat variables in their choice of suitable nest boxes (e.g. Willner et al. 1983; Pogue and Schnell 1994). Few studies have tested whether bird or bat species show a preference for the height at which boxes are placed in trees. Most studies placed boxes at heights of 2–6 m, presumably because artificial hollows placed at low heights will be quicker and safer to monitor. These relatively low heights may have limited the use of boxes but for the species detected there is little evidence as yet that greater heights will be preferred (Table 1). Stebbings and Walsh (1991) suggested that some bat species in Great Britain prefer boxes to be at least 5 m high. The height at which boxes are placed may influence rates of predation (Milledge 1978; Menkhorst 1984) but this remains to be tested. Lumsden et al. (2002a) found that a large proportion of tree roosts of lesser long-eared bats were <5 m high, whereas most roosts of Gould’s wattled bats were >5 m high. However, the latter species accounted for 91% of bats present in boxes placed 4–6 m high in Organ Pipes National Park (Irvine and Bender 1995; R. Bender, unpubl. data). Use of artificial tree hollows by Australian birds and bats The actual spacing of boxes may influence their frequency of use. Nest boxes located too close together may be subject to intraspecific competition and nesting failure of birds (see Krebs 1998). Among North American waterfowl, nest boxes placed at high density can lead to high levels of conspecific brood parasitism (multiple females depositing eggs into one clutch), which can have adverse population effects (Eadie et al. 1998). Studies of tree hollow use in white-tailed black-cockatoos (Calyptorhynchus latirostris) and Major Mitchell’s cockatoos (Cacatua leadbeateri) suggest that conspecific interactions lead to a wide spacing of nest trees (Saunders et al. 1982). Given the lack of proper testing of elements of hollow design and placement, there is a great need for research on this topic. The effective use of artificial hollows in research and management will depend on sound knowledge of hollow design and placement. Hollow availability The availability of natural hollows is commonly assumed to have a strong influence on the frequency of use of artificial hollows (e.g. Newton 1994). However, few studies in Australia have considered this issue in detail with many studies having been conducted where it was apparent that a dearth of natural hollows was precluding use of an area (e.g. Irvine and Bender 1995) or hindering reproduction (e.g. Orange-bellied Parrot Recovery Team 2006). Three studies in Victoria (Ambrose 1982; Calder et al. 1983; Menkhorst 1984) each installed boxes at several locations where natural hollow abundance was either low or high. In all studies boxes were used more frequently by Australian owlet-nightjars where natural hollows were scarce. Whitethroated tree-creepers and crimson rosellas showed higher use where natural hollows were scarce in two studies but no difference in one study (crimson rosella) or higher use where natural hollows were abundant (white-throated tree-creeper). Ambrose (1982) found box use by barn owls (Tyto alba) to be more frequent in forest with few hollows whereas boobook owls (Ninox novaeseelandiae) showed no difference between sites with few or many hollows. Menkhorst (1984) found that grey shrike-thrushes (Colluricincla harmonica) only used boxes where natural hollows were abundant. Few studies have been conducted of bats in this context. Golding (1979) found that 8% of boxes were used by the lesser long-eared bat and Gould’s wattled bat where hollows were abundant, compared to 3% where no hollows occurred and 43% where hollows were in low abundance. Ambrose (1982) reported a single bat detection in a box whereas Menkhorst (1984) reported none. Smith and Agnew (2002) found that Gould’s longeared bat used boxes frequently at two sites with no hollows and there was little or no use where natural hollows were either in low or high abundance. Goldingay et al. (2007) observed low use of bat boxes across five locations that varied in abundance of natural hollows including the two sites where Smith and Agnew (2002) reported frequent use. For both birds and bats, it appears that site factors (habitat, competitors) may have a strong influence on box use that may mask a species’ response to natural hollow abundance and that properly replicated studies are required to better understand whether natural hollow abundance influences use of artificial hollows. Due to the mobility of birds and bats it is Wildlife Research 87 likely that they can readily colonise areas devoid of natural hollows if nest and roost boxes are installed. Seasonal patterns related to breeding Many species of hollow-using birds only use hollows for breeding, so their breeding patterns will determine when nest boxes are used. This may require frequent checking of artificial hollows during the year to accurately document use as well as to determine the adequacy of the artificial hollows provided for breeding. The crimson rosella, white-throated tree-creeper (Calder et al. 1983; Menkhorst 1984), grey shrike-thrush (Menkhorst 1984), eastern rosella (Platycercus eximius) and red-rumped parrot (Psephotus haematonotus) (Trainor 1995a) used nest boxes in the spring and summer periods for breeding, although nest site preparation (hole chewing and nest building) by the crimson rosella (Calder et al. 1983; Pell and Tidemann 1997a; Krebs 1998) and white-throated tree-creeper (Calder et al. 1983) was observed during winter. In Victoria, crimson rosellas used artificial hollows until chicks were fledged in mid-January, a period of 56 days following egg laying (Golding 1979). Orangebellied parrots laid eggs between November and January, and had an incubation period of 24 days and a nestling period of 35 days (Orange-bellied Parrot Recovery Team 2006). Glossy blackcockatoos on Kangaroo Island laid eggs from late January to late July, and had an incubation period of 30 days and a nestling period of 90 days (Garnett et al. 1999). Red-tailed blackcockatoos laid eggs during September to December, and had an incubation period of 30 days and a nestling period of 87 days (Commonwealth of Australia 2006). Maned or wood ducks (Chenonetta jubata) bred in nest boxes near Canberra between July and December with a 34-day incubation period (Briggs 1991). The nocturnal Australian owlet-nightjar used nest boxes primarily in spring and summer for breeding, but as a diurnal roost site throughout the year (Calder et al. 1983; Menkhorst 1984; Trainor 1995a). The white-throated tree-creeper was found to use nest boxes for roosting all year except during summer (Ambrose 1982). The boobook owl and barn owl used nest boxes during winter and spring, and less frequently during summer (Ambrose 1982). The above data indicate that seasonal patterns of use can be highly variable among bird species and consequently the design of monitoring programs will need to accommodate this. Few data are available for bats but if artificial hollow use mirrors natural hollow use then the requirements of some species may differ during the breeding period from other times of the year, when females aggregate into maternity colonies when breeding (e.g. Churchill 1998). For example, females of the lesser longeared bat preferred enclosed hollows with fissure entrances in very large-girth trees during breeding periods compared to when not breeding (e.g. Lumsden et al. 2002a). Golding (1979) found that individual or small groups of Gould’s wattled bat used roost boxes throughout most of the year but female-dominated groups (up to 39 bats) occurred in spring and for the lesser long-eared bat (10 bats) in December in a few boxes. Smith and Agnew (2002) observed small numbers (four to eight) of Gould’s long-eared bat in several months but one aggregation of 16 females with young in October. R. Bender (unpubl. data) observed a summer peak in breeding by Gould’s wattled bats in boxes. The frequency with which patterns of use mimic those within natural hollows by 88 Wildlife Research catering for non-breeding females as well as breeding aggregations should indicate the effectiveness of artificial hollows. Studies that involve a choice of box size and location are needed to better understand the potentially varied seasonal requirements of bats. Temperature The influence of temperature on hollow use will vary depending on whether a species requires hollows throughout the year (bats, hollow-roosting birds) or just during the breeding season (hollow-breeding birds). Temperature within hollows may be critical to the daily thermoregulation of individuals or in promoting faster rates of growth by developing young (e.g. Dawson et al. 2005). The temperature inside artificial hollows can be an important factor in their use if hollows either exceed or do not reach a preferred thermal zone. There may be a seasonal element to this due to the poor insulative properties of artificial hollows (Calder et al. 1983; Kerth et al. 2001), so they may be ignored in winter when temperatures are low or ignored in summer when temperatures are high (R. Goldingay, unpubl. obs). This seasonal pattern is likely to vary latitudinally with climate and may lead to variation in the preferred orientation of entrances to nests (e.g. Burton 2007) or the preferred position of an artificial hollow (full sun v. shade). Furthermore, elevational gradients in temperature may also have an influence. Therefore, where we refer to studies from different geographic locations below, we acknowledge that some findings may be location specific. In Australia, the influence of the thermal environment in artificial hollows has not been well studied. Norman and Riggert (1977) found that temperatures inside black plasticdrum nest boxes in southern Western Australia were an average of 12C hotter than ambient. Temperatures in the boxes commonly exceeded 35C in October but were not assessed later in the year when they might have been much hotter. Menkhorst (1984) suggested that reduced survival of white-throated tree-creeper hatchlings in his study may have resulted from overheating within nest boxes. Calder et al. (1983) have shown that aspect, wall thickness, entrance position and box depth can all have profound influences on temperatures within artificial hollows. Overly high summer temperatures may not be such a problem for hollow-breeding birds that breed in spring, but the young of some species (e.g. crimson rosella, white-throated tree-creeper) may not fledge until January–February (Calder et al. 1983). Some bird species may choose warm hollows in spring due to the positive influence that temperature can have on incubation and nestling development (e.g. Ardia et al. 2006). Consistent with this was the observation that Bechstein’s bats (Myotis bechsteinii) in Germany preferred sun-exposed boxes during lactation whereas shaded boxes were preferred pre-lactation (Kerth et al. 2001). Temperature is a critical issue for microbats in roost selection (Boyles 2007) and therefore roost box selection. Many microbats are quite small in size (<20 g) (Churchill 1998) so energy conservation through thermoregulation will be critical to their survival and the need for passive rewarming from daily torpor may influence roost selection (e.g. Turbill et al. 2003a, 2003b; Ruczynski 2006; Turbill 2006). This indicates that thermal environments within roost boxes should have a strong R. L. Goldingay and J. Stevens influence on their occupation and frequency of use (Stebbings and Walsh 1991). Indeed, this has been observed in microbats in Europe and North America. Lourenço and Palmeirim (2004) demonstrated that soprano pipistrelles (Pipistrellus pygmaeus) in Portugal were tolerant of high temperatures (~40C) and preferred black roost boxes over white or grey boxes. These boxes attained greater diurnal temperatures and had temperature profiles more similar to the roost cavities where many bats had been roosting. Brittingham and Williams (2000) found that aspect had a pronounced influence on temperatures within bat boxes in Pennsylvania and that bats used boxes that experienced long periods (7 h) of direct sun but not those with shorter periods (5 h) of sun. In Germany, bats used sun-exposed bat boxes more frequently than those that were shaded (Kerth et al. 2001). In contrast, Neilson and Fenton (1994) recorded little use of four types of bat box in New York State but noted that the mean temperature in boxes was less than that in occupied roof cavities. Thus, placement of boxes where suitable thermal ranges are not experienced may result in avoidance by bats. No studies in Australia have placed bat boxes with the explicit purpose of creating hot microclimates inside. Menkhorst (1984) placed boxes with a north or south entrance orientation but detected no bats. Irvine and Bender (1995) placed bat boxes across a range of aspects but most were predominantly in shaded positions that they suggested may have contributed to a low frequency of use by bats. Subsequent monitoring (R. Bender, unpubl. data) has shown that the greatest use by Gould’s wattled bat, the most frequent occupant, occurred in summer, which is consistent with the hypothesis that the boxes do not have optimal thermal environments. Smith and Agnew (2002) placed boxes equally across the range of cardinal compass points but it is unlikely that this had a consistent influence on direct sun due to the cover of tree canopies at these sites (R. Goldingay, pers. obs). Goldingay et al. (2007) placed bat boxes so that they would not receive direct sun in the hottest months of the year because the feathertail glider was the focus of their study. Lumsden et al. (2002a) found that the highest proportion of roosts of the lesser long-eared bat (tree hollows and some artificial structures) and Gould’s wattled bat (all were tree hollows) had a north-north-east aspect. A north aspect is likely to have a higher temperature profile than other aspects but the selected aspects were not compared to those of available sites. However, exfoliating bark of trees is expected to have a random or even distribution and yet Turbill et al. (2003a) found that the roosts under bark of the lesser longeared bat were significantly concentrated on the north-west aspect of trees. Future studies that target bats must position roost boxes with consideration for the thermal requirements of bats. Studies should investigate the influence of box design on temperature profiles. Studies are also needed to investigate how the thermal requirements of birds during nesting may influence hollow choice. Furthermore, studies are needed to determine how latitude and altitude may interact with temperature, and lead to varied responses to artificial hollow placement. Competitive interactions Some hollow-using species may directly or indirectly exclude others, which may reduce the effectiveness of artificial hollows Use of artificial tree hollows by Australian birds and bats for some target species. Competition may arise from native or non-native species. The impacts of displacement by non-native species is relatively well documented. The common myna (Pell and Tidemann 1997a; Homan 1999; Harper et al. 2005b), common starling (Sturnus vulgaris) (Ambrose 1982; Trainor 1995a; Pell and Tidemann 1997a, 1997b) and introduced honeybee (Ambrose 1982; Trainor 1995b; Emison 1996; Pell and Tidemann 1997a; Homan 1999; Harper et al. 2005b) are the species of most concern, though the black rat could also be a problem at some locations. These species have the potential to deter native species from using nest boxes and reduce the number of available hollows (Trainor 1995b; Pell and Tidemann 1997b; Smith and Agnew 2002). The common myna was found to outcompete several species of native parrot and the common starling can cause nest failure of native species (Ambrose 1982; Trainor 1995a; Pell and Tidemann 1997b; Gleeson 1999; Orange-bellied Parrot Recovery Team 2006). Harper et al. (2005b) found that mynas occupied 45 of 120 nest boxes in Melbourne over a 7-month period, and rebuilt their nests following removal. Although competition for nest sites with the starling has led to few declines among cavity-nesting birds in the USA (Koenig 2003), the longer-term effects of starlings and mynas on Australian birds are unknown. The invasion of both artificial and natural hollows by feral honeybees can be a significant management issue (e.g. Trainor 1995b; Wood and Wallis 1998). The recovery plans of the Kangaroo Island glossy black-cockatoo and orange-bellied parrot have identified the need for regular maintenance to deter and remove feral honeybees from artificial and natural hollows (Mooney and Pedler 2005; Orange-bellied Parrot Recovery Team 2006). Feral honeybee invasion has been reported in many studies of nest boxes (e.g. Wood and Wallis 1998; Harper et al. 2005b; Goldingay et al. 2007) and needs to be documented in detail so its incidence and treatment can be better understood. For example, the use of carpet (Franks and Franks 2003) and insecticide strips (Irvine and Bender 1995; Soderquist et al. 1996) inside nest boxes have been trialled to prevent honeybee infestations but few details have been reported. Nest box design may mitigate the impact of pest species and reduce the need for costly maintenance. Lumsden (1989) reported that starlings did not use nest boxes painted white inside. In a preliminary study by Homan (2000), the use of a baffle installed on the front of nest boxes successfully excluded the common myna without excluding native species. A slit entrance for bat boxes has been suggested to exclude competition and predation by introduced birds (Smith and Agnew 2002). Small nest box volume may reduce hive building by honeybees (Goldingay et al. 2007). The influence of pest species is relevant to the success and economic cost of artificial hollows in management, and further investigation using an experimental approach is needed. Some native hollow-using species will displace others. Boxes installed in Australia for use by bats may be used by arboreal mammals (Smith and Agnew 2002; Goldingay et al. 2007). Irvine and Bender (1995) reported that 2 of 10 bat boxes had nests of sugar gliders but over time all of these boxes had sugar gliders present either often or occasionally (R. Bender and R. Irvine, unpubl. data). Small entrance size will not overcome this problem because feathertail gliders will use entrances of 1.5 cm and their nests can completely fill a box (Fig. 7a). Goldingay et al. (2007) Wildlife Research 89 reported that 6 of 12 bat boxes at locations in south-east Queensland had feathertail gliders or their nests and only four boxes were used by bats. This may require a reliance on openbottom roost boxes for bats that preclude the construction of a leaf nest inside (Fig. 7b, c). Arboreal marsupials are likely to compete with birds as well but the extent of this is not well documented. Menkhorst (1984) provided circumstantial evidence that two bobucks (Trichosurus caninus) discouraged the use of many nest boxes by other mammals and birds. Common brushtail possums (Trichosurus vulpecula) will kill nestlings present in artificial hollows. On Kangaroo Island, this has required placing metal guards around trees containing hollows used by glossy black-cockatoos to prevent possums gaining access (Garnett et al. 1999). Sugar gliders compete with orange-bellied parrots for nest sites and may even kill incubating birds (Orange-bellied Parrot Recovery Team 2006). This requires specific research to identify methods to reduce this impact. For example, squirrel gliders and sugar gliders may favour rear-entry nest boxes (the entrance occurs on the back of the box; see Beyer and Goldingay 2006) and be less likely to use other front-entry box types (with the entrance on the front or side of the box) when a choice is provided (Goldingay et al. 2007; R. Goldingay, unpubl. obs). Loeb and Hooper (1997) found that the provision of nest boxes reduced the use of natural cavities required by endangered red-cockaded woodpeckers (Picoides borealis) by competing cavity users. Crimson rosellas on Norfolk Island competed for nest boxes with endangered Norfolk Island boobook owls (Olsen 1996) and for natural hollows with endangered Norfolk Island green parrots (Cyanoramphus novaezelandiae cookii) (Hill 2002). Galahs (Cacatua roseicapilla) have been implicated in the loss of eggs and little corellas (Cacatua pastinator) implicated in the loss of nestlings of the glossy blackcockatoo (Garnett et al. 1999). Deployment of decoy nest boxes may help to alleviate competition and provide easier access to these species for control. Emison (1996) recorded yellow-tailed black-cockatoos (Calyptorhynchus funereus), long-billed corellas (Cacatua tenuirostris), maned ducks and owls using artificial hollows erected for red-tailed black-cockatoos. The expansion in the geographic ranges of common hollow-nesting parrots may put competitive pressure on other hollow-using native species. Clearly, competition from non-target species can have a substantial influence on the effectiveness of any artificial hollow program. However, although the problem is well acknowledged by many authors it remains poorly documented and this will hamper attempts to reduce its impact. Therefore, research must be conducted to understand the magnitude of the problem and how it might be managed. Artificial hollow applications Artificial hollows have been used in a variety of ways for research and management. Beyer and Goldingay (2006) recognised for arboreal marsupials that nest boxes had three research applications (detection of species, ecological studies, investigation of nest-box design and placement) and three management applications (threatened species recovery, species introductions, strategic placement). These also apply to birds and 90 Wildlife Research R. L. Goldingay and J. Stevens (a) (b) (c) Fig. 7. (a) A feathertail glider nest completely filled this wedge-shaped bat box. (b) An open-bottom design bat box (42 17 15 cm; front panel 32 cm). (c) Gould’s long-eared bats in an open-bottom box. Photos: (a, b) R. Goldingay; (c) J. Lindsay. bats. In addition, we also recognise ‘species establishment through habitat enhancement’ and ‘hollow-bearing tree offset’. These research and management applications are discussed in detail below. Research applications Species detection Nest boxes have been used as a survey tool to determine the distribution and abundance of cryptic arboreal mammal species (Beyer and Goldingay 2006). Using artificial hollows in this way may not be of great importance for birds and bats, which may be detected more readily using conventional survey techniques, particularly sonar detection for bats (but see Flaquer et al. 2007). Homan (1999) installed 12 nest boxes to survey for small parrots but eight were quickly occupied by common starlings and common mynas. However, the value of nest and roost boxes is that they can be left in place for long periods and may provide insights not readily gained from periodic survey using other techniques. In Great Britain, the placement of large numbers of bat boxes has produced significant range extensions for some species (Stebbings and Use of artificial tree hollows by Australian birds and bats Walsh 1991). Artificial hollows could be deployed to identify species that might respond to hollow provision, as a precursor to a management application. Ecological research Nest boxes have been used extensively to investigate the breeding biology and life histories of hollow-using birds in North America and Europe (Koenig et al. 1992; Evans et al. 2002). In Australia, only a small number of published studies have used artificial hollows to investigate the ecology of birds and none have investigated the ecology of bats. Norman (1982) used nest boxes to study egg laying and incubation in the chestnut teal. Briggs (1991) used nest boxes to investigate intraspecific nest parasitism in maned ducks. Pell and Tidemann (1997a) used nest boxes to study factors that affect the breeding success of the crimson rosella, and eastern rosella when in competition with introduced hollow-using birds. Pell and Tidemann (1997b) investigated the ecology of the common myna using nest boxes. Nest boxes have allowed detailed investigation of breeding biology, patterns of food allocation in broods, and nestling growth and survival in the crimson rosella (Krebs 1998, 1999, 2001; Krebs et al. 1999, 2002; Krebs and McGrath 2000). Nest box use by the glossy black-cockatoo has allowed aspects of its breeding ecology to be described (Garnett et al. 1999). Baltz and Clark (1999) used nest boxes to investigate the influence of conspecifics on nest choice in budgerigars (Melopsittacus undulates) in captivity. The above studies highlight the value of nest boxes as a research tool, in providing access to animals that may not be available any other way. Artificial hollow preferences The design of artificial hollows can have a pronounced influence on their frequency of use (see above). Despite this, few preference studies have been conducted in Australia. Norman and Riggert (1977) examined the use by waterfowl of eight nest box types that differed in construction material and dimensions. This revealed greater use of a thick wooden box (ammunition box) but few of some types were installed and it appears that a direct choice of different box types was not provided. Menkhorst (1984) provides the first example of a properly replicated choice experiment in which combinations of four entrance sizes, three height placements and two aspect types were used. He cautioned about the comparison of aspect because boxes were placed on the east or west side of a tree and likely to have received approximately equal exposure to the sun. He identified a preference by crimson rosellas for entrance size but sample sizes for owlet-nightjars, white-throated treecreepers and grey shrike-thrushes were too low to demonstrate any choice. No bats were detected in this study. R. Bender and R. Irvine (unpubl. data) explored the relationship between entrance size and bat body size with a small number of roost boxes. They suggested that Gould’s wattled bat (10 g) preferred a slit entrance size >15 mm, whereas smaller species, the large forest bat (7 g) and the southern forest bat (Vespadelus regulus) (5 g), preferred slit entrance sizes <12 mm. An entrance size <8 mm was unused by bats. This suggests that competitive interactions may occur Wildlife Research 91 among bats for roost sites and boxes with different entrance sizes may be required to avoid dominance by one bat species. It is clear that many trials of different bat box designs are needed in Australia. Furthermore, different bat species may prefer different designs (other than just entrance size) so different bat box designs should be field tested at multiple locations. Lindenmayer et al. (2003) placed four box types (small with small entrances (5 cm) or large with large entrances (10 cm), installed high or low) on 24 plots across two locations to examine box preferences by Leadbeater’s possum (Gymnobelideus leadbeateri). The owlet-nightjar and crimson rosella were recorded on only one occasion over a 3-year period. No other birds or bats were recorded. In a study designed to target Leadbeater’s possum, Harley (2004) installed 150 nest boxes of various volumes, wall thicknesses and with 47–65 mm diameter entrances but did not offer a direct choice of box type. Over a 5-year period he observed a small amount of use by owlet-nightjars, white-throated tree-creepers and one rosella but no use by any bats (D. Harley, pers. comm.). This may have been due to 76% of boxes having possums or their nest present within. Goldingay et al. (2007) placed four box types on 45 plots across five locations to examine box preferences by feathertail gliders. They observed bats using only a wedge-shaped bat box with a slit-entry at the bottom. Avoidance of another slit-entry box may have been due to a lack of mesh inside the box to which bats could attach. Two rear-entry box designs were probably unsuitable for bats. Many further studies that offer a choice of designs or design elements are needed to advance our understanding of artificial hollow preferences. Management applications Threatened species recovery Artificial hollows have been used in recovery programs for several threatened bird species, but the deployment of roost boxes for threatened bats has only just begun. Nest boxes have been used in areas where natural hollows have been depleted. Information on the use of nest boxes by threatened Australian parrots is largely only documented in the recovery plans for these species and in a cursory way. This needs to be properly reported in the peerreviewed literature for the usefulness of different artificial hollows and management programs to be evaluated, particularly so that this information can inform other projects. Publication of research findings is a fundamental element of conservation biology (Calver and King 1999). Nest boxes have been in use for decades to supplement nesting habitat of the critically endangered orange-bellied parrot, and both wild and released captive-bred birds used nest boxes (Orange-bellied Parrot Recovery Team 2006). The use of nest boxes has become fundamental to assessing population recovery by allowing access to individuals for banding and for assessing reproductive success (Orange-bellied Parrot Recovery Team 2006). Artificial hollows have become important in the recovery of the endangered Kangaroo Island glossy black-cockatoo. Over 80 artificial hollows made from PVC plumbing pipe (Fig. 3) have been used to supplement natural nesting sites (Pedler 1996; Mooney and Pedler 2005). Over several years, 25–35% of successful breeding occurred in nest boxes, with breeding 92 Wildlife Research success similar (and in some years greater) to that in natural hollows (Mooney and Pedler 2005). This program could inform the use of artificial hollows in other recovery programs. Artificial hollows consisting of hollows cut from fallen trees have been used to supplement the breeding habitat of the southeastern red-tailed black-cockatoo. A program was trialled for this subspecies and included erecting some nest boxes on 12 m-high power poles (Emison 1996). Nest boxes were quickly occupied, and over a 2-year period 30% of all artificial hollows were used (Emison 1996). Approximately 60 artificial hollows, comprising both PVC plumbing pipe and natural hollows cut from fallen trees (Figs 2, 3) have been installed on power poles in the last 5 years but only salvaged natural hollows (at least seven) were used (R. Hill, pers. comm.). The recovery plan for this species has recognised that although the availability of natural nest hollows is not currently limiting the population, dead nest trees are collapsing at a rate of 4–7% per year, and this is likely to be a serious threat in years to come (Commonwealth of Australia 2007). The recovery plan recommends for the situation to be monitored and the artificial hollow program expanded only if significant nest tree losses are observed (Commonwealth of Australia 2006). Artificial hollows made from fallen branches were installed to supplement the breeding habitat of the turquoise parrot (Neophema pulchella) in Chiltern Park, Victoria (Quin and Baker-Gabb 1993). Two were used for nesting. Artificial hollows made from salvaged tree hollows and erected in known breeding areas have been used in breeding by Norfolk Island green parrots (Hill 2002). Nest boxes were installed in two studies in Tasmania to target the endangered forty-spotted pardalote (Pardalotus quadragintus) but were only successful in enabling the striated pardalote (Pardalotus striatus) to breed (Milledge 1978; Woinarski and Bulman 1985). Nest boxes are currently being trialled as part of the recovery plan for this species (Threatened Species Section 2006). Roost boxes could play an important role in the recovery of threatened bats but existing studies are inadequate to guide this application. The recovery plan for the Christmas Island pipistrelle (Pipistrellus murrayi) proposes to install roost boxes to supplement the natural roosts of this species, which may include beneath tree bark, under dead palm fronds, beneath tree canopies, and in tree hollows (Schulz and Lumsden 2004). The eastern false pipistrelle (Falsistrellus tasmaniensis), a vulnerable species in New South Wales, has been recorded using nest boxes in Victoria (Golding 1979; Ward 2000), suggesting there is potential to use roost boxes in the recovery of this species. Existing studies demonstrate the usefulness of artificial hollows to threatened species recovery. However, there is a need for further research to improve and expand current use. This application has potential for threatened bats but will depend on a dramatic increase in our understanding of preferred roost box designs. Species introductions We recognise this application as distinct from threatened species recovery because it may not always involve such species and the approach may be quite different. It involves installing R. L. Goldingay and J. Stevens artificial hollows at a location where a species is to be introduced. The deployment of artificial hollows to introduce a species at a site has been documented for just one species. Several nest boxes were installed on Norfolk Island in 1987 for the reintroduction of the endangered Norfolk Island boobook owl (Ninox novaeseelandiae undulata) (Olsen 1996). Male birds from the closest extant subspecies, the New Zealand morepork (Ninox novaeseelandiae novaeseelandiae), were translocated to the island. The nest boxes enabled successful breeding to occur. Species establishment through habitat enhancement Because birds and bats are highly mobile it is quite possible to attract and establish species at a location by the installation of artificial hollows. This is as a form of habitat enhancement where the decline or disappearance of hollow-using species is recognised and artificial hollows have been installed to prevent the permanent loss of biodiversity. Currently, the only successful example of this management application is that by Irvine and Bender (1995) for bats. Bat boxes were installed in Organ Pipes National Park in Victoria in 1992 to facilitate the establishment of bat populations within regenerating forest in the park where habitat restoration had commenced in 1972 (Irvine and Bender 1995). Bat boxes were not occupied for 30 months, which is in stark contrast to within 3 months of installation documented by Smith and Agnew (2002) in boxes in Queensland. The number of bats in Organ Pipes National Park increased from 15 per check in 1994–95 to >100 per check in 2004–05 from 34 boxes (R. Bender, unpubl. data). Gould’s wattled bat comprised 91% of records but a small number of large forest bats also consistently used the boxes. Boyd and Stebbings (1989) reported a doubling over a 10-year period in a population of brown long-eared bats (Plecotus auritus) supported by roost boxes in managed forest in Great Britain. These observations suggest that the local bat population in Organ Pipes National Park increased in size over time so the initial delay may have been due in part to a small local population. Harper et al. (2005b) installed nest boxes as habitat enhancement (~3 per ha) for vertebrate fauna across remnant vegetation within the urban and suburban landscape of Melbourne. Although nest box design favoured large arboreal mammals (large entrance and volume), a small number were used by rainbow lorikeets (six), an eastern rosella (one) and a galah (one). The provision of artificial hollows may also allow hollowusing species lost from urban areas to recolonise. Other studies that install artificial hollows as habitat enhancement have been planned and will target a range of birds and bats (B. Law and R. Kavanagh, pers. comm.; R. Goldingay, unpubl. data). This management application has considerable merit but is currently hampered by the lack of understanding of the factors that influence the use of artificial hollows (see above). Many trials are needed to determine which hollow designs should be installed, the number of each design, the most effective placement, and whether particular species have benefited from such habitat enhancement. Evidence that this application has been successful would be the on-going use and breeding in artificial hollows (Petty et al. 1994), rather than only sporadic use by species. Such trials should be Use of artificial tree hollows by Australian birds and bats conducted in an adaptive management framework so that changes can be made as information accumulates on factors that improve effectiveness. Monitoring will be fundamental to such studies and needs to be continued for 2–5 years to provide the best insights. Hollow-bearing tree offset Another management application that has been used by land managers (e.g. local government, road agencies, power companies) is to install artificial hollows to compensate for hollow-bearing trees lost during authorised clearing (Fig. 8). This might occur either adjacent to the development site or possibly away from the site if a landscape approach is taken to managing hollow-using species. We stress that the potential value of artificial hollows should not be used to justify the removal of hollow-bearing trees. The merit of this application is that birds and bats can be highly mobile and providing some replacement hollows may assist some species as an interim solution while other longer-term measures are devised. If such a management application is used there needs to be monitoring over a 2–5-year period to document the outcome. Currently, information is lacking to demonstrate the value of such a use of artificial hollows to hollow-using species so there is an obvious need for research on this application. Strategic placement This management application emphasises the specific locations where artificial hollows are placed. This might be a specific location in a landscape such as within wildlife corridors or it might be a location where a specific objective is to be achieved such as to attract particular kinds of hollow-using species (Beyer and Goldingay 2006). Although it will be concerned with establishing species in an area it primarily differs from ‘species establishment’ because fewer hollows will be used, their installation will be highly targeted and a more specific objective will be stated. In Europe there is increasing recognition of the value of nest boxes to attract hole-nesting birds as a means of controlling insect pests in forests and orchards (Mols and Visser 2002). Smith and Fig. 8. Bat boxes installed on powerpoles through a powerline easement in Brisbane. Some poles also have parrot boxes attached. These boxes were installed after expansion of an existing powerline easement. Photo: R. McLean. Wildlife Research 93 Agnew (2002) hypothesised that small mammals, particularly bats, could control insects in farm forests and therefore provide a health benefit to the trees. They installed bat boxes in small hardwood plantations in south-east Queensland and had success in attracting a small number of bats to their sites but better success with attracting feathertail gliders and squirrel gliders, which may have discouraged greater use of the boxes by the bats. The work in Organ Pipes National Park provides insight to the potential of strategic placement with regard to bats. In 2004–05, >100 bats were captured per round of box checks from 34 roost boxes scattered through 5 ha of regrowth forest (R. Bender, unpubl. data). Artificial hollows are being trialled in northern Australia to increase barn owl and masked owl numbers for the control of rodent pests in sugarcane crops (Gibbons and Lindenmayer 2002). Studies on the strategic use of artificial hollows in Australia are in their infancy. Further research is needed for this application. Conclusion In many landscapes across Australia the collapse of hollowbearing trees has outpaced the recruitment of replacement hollows and future shortages in this resource are inevitable (Saunders 1979; Lindenmayer et al. 1990, 1997; Saunders et al. 2003; Courtney and Debus 2006; Commonwealth of Australia 2007; Beyer et al. 2008). The provision of artificial hollows is likely to be the most appropriate interim solution to this shortage but existing information on the use of artificial hollows is too limited to enable this response to be effective and progress in this field has been slow. Applying our criteria of requiring more than a single record of artificial hollow use and some details of the hollows, we found information was documented for just 15 of 114 hollow-using bird species and 8 of 41 hollow-using microbat species. The lack of information is most stark for microbat species. The deployment of roost boxes for bats in Australia is clearly in its infancy. Progress has in part been hampered by the relatively infrequent use of artificial hollows by bats, which may be a consequence of positioning roost boxes without regard for creating suitable thermal environments. Selecting roosts to enable passive rewarming from daily torpor may be quite widespread among Australian tree-roosting bats (Turbill et al. 2003a, 2003b; Turbill 2006) and this will influence their ability to use artificial hollows. Correct positioning of artificial hollows poses a challenge because deployment in forested habitats may preclude positioning boxes in highly favourable microsites due to shading from canopy cover. Some natural roosts of bats in mature forest may be located high in trees (e.g. Herr and Klomp 1999; Lumsden et al. 2002a) and have low canopy cover (e.g. Campbell et al. 2005) to minimise shading. However, given that bats may commute over long distances where tree hollows are scarce (e.g. Lunney et al. 1985, 1988; Lumsden et al. 2002b), it may be appropriate to target edge and forest gap sites to place artificial hollows where sun exposure will be optimal. A key deficiency in deploying artificial hollows in research and management of hollow-using birds and bats is in understanding the most effective designs to use. Research 94 Wildlife Research suggests that most species favour artificial hollows with entrances just wide enough to enter. This enables avoidance of larger competitors and possibly predators. Other elements of design have not been properly investigated though guidance can be provided by research on natural hollows, which show that hollow depth is also important (e.g. Saunders et al. 1982; Gibbons et al. 2002). Some bird species will use artificial hollows made from salvaged tree limbs but it is not well documented whether constructed hollows that emulate these have been tried adequately. Bats may have different requirements during the year (breeding v. non-breeding periods) and this might require the provision of several designs. So few studies have been conducted of bats using roost boxes in Australia that suitable designs remain largely unknown. There is an obvious need for many field experiments that compare different hollow designs or which vary specific design elements for birds and bats. This should lead to better knowledge of designs preferred by different species. Another aspect that requires further investigation is the extent that competing species may pre-empt target species from artificial hollows. Many studies have described potential competition among hollow users but the seriousness of this to management is not well documented. In some cases competitors may favour a specific box design (e.g. arboreal mammals that displace birds or bats) and exert less interference if their hollow needs are catered for. This requires specific research to address. Furthermore, there are various issues that relate to the maintenance of artificial hollows, such as the occupation by feral honeybees and the collapse of boxes (Beyer and Goldingay 2006). This also must be addressed by research with particular attention given to the cost of maintenance (see also Harley 2006). One aspect of artificial hollow placement that is not well understood and in need of research is whether species show a preference for the height at which boxes are placed on trees. This is of considerable management relevance because this can have cost and safety implications in effectively employing artificial hollows. Available information suggests that heights >5 m are rarely needed (Table 1). The only examples where hollows have been placed very high is that of the Kangaroo Island glossy blackcockatoo with hollows at ~16 m above ground (Pedler 1996) and the red-tailed black-cockatoo with hollows at 12 m above ground (Emison 1996). It appears that artificial hollows have been placed at an equivalent height to natural tree hollows (Pedler 1996). Such hollows will require an enormous amount of time to check and maintain. Given that red-tailed black-cockatoos have used tree hollows at heights as low as 4.4 m (mean height 7 m) in Western Australia (Saunders et al. 1982), it is possible that very high hollows are not needed. Examples where bats select high roosts (i.e. >10 m) may reflect their preference for suitable thermal environments, which may be replicated by placing artificial hollows in canopy gaps (see above). Studies in Australia on the use of artificial hollows have lagged behind those conducted in Europe and North America (e.g. Boyd and Stebbings 1989; Newton 1994; Petty et al. 1994; Pogue and Schnell 1994; Brittingham and Williams 2000; Kerth et al. 2001), which is surprising given that a greater proportion of wildlife species in Australia compared to other continents is dependent on tree hollows for survival (see Saunders et al. 1982; Newton 1994; R. L. Goldingay and J. Stevens Gibbons and Lindenmayer 2002). The small number of Australian studies currently available is insufficient to allow many generalisations. Artificial hollows have enormous potential to become an important management tool for hollowusing species in Australia but this can only be realised by conducting many additional studies. This needs to occur across a range of species and landscapes to maximise our understanding of interactions between species and different environments. Furthermore, species of Australian birds and bats are likely to respond to similar factors in choosing artificial hollows as taxonomically equivalent species overseas (e.g. parrots, vespertilionid bats), so investigations can provide independent tests of current hypotheses concerning influences on use, or of management applications. This will allow greater generalisation and can also lead to work conducted in Australia informing research and management of hollow-using species in other countries. Acknowledgements The comments of Brendan Taylor, Geoff Smith and two anonymous referees helped improve this paper. This paper has been informed by several nest box projects conducted in Brisbane and an artificial hollow project conducted at Brunswick Heads, NSW. We thank Brisbane City Council for support of our research in Brisbane and Abigroup for assistance with the project at Brunswick Heads. Matthew Grimson and Geoff Smith are thanked for sharing ideas and field work that have provided background to this paper. We thank Robert Bender and Robert Irvine for their pioneering work with bat boxes in Organ Pipes National Park. References Ambrose, G. J. (1982). An ecological and behavioural study of vertebrates using hollows in eucalypt branches. Ph.D. Thesis, La Trobe University, Melbourne. Ardia, D. R., Perez, J. H., and Clotfelter, E. D. (2006). Nest box orientation affects internal temperature and nest site selection by tree swallows. Journal of Field Ornithology 77, 339–344. doi: 10.1111/j.15579263.2006.00064.x Baltz, A. P., and Clark, A. B. (1999). Does conspecific attraction affect nest choice in budgerigars (Melopsittacus undulates: Psittacidae: Aves)? Ethology 105, 583–594. doi: 10.1046/j.1439-0310.1999.00440.x Bennett, A. F., Lumsden, L. F., and Nicholls, A. O. (1994). Tree hollows as a resource for wildlife in remnant woodlands: spatial and temporal patterns across the northern plains of Victoria, Australia. Pacific Conservation Biology 1, 222–235. Beyer, G. L., and Goldingay, R. L. (2006). The value of nest boxes in the research and management of Australian hollow-using arboreal marsupials. Wildlife Research 33, 161–174. doi: 10.1071/WR04109 Beyer, G. L., Goldingay, R. L., and Sharpe, D. J. (2008). The characteristics of squirrel glider (Petaurus norfolcensis) den trees in subtropical Australia. Australian Journal of Zoology 56, 13–21. doi: 10.1071/ZO08053 Boyd, I. L., and Stebbings, R. E. (1989). Population changes of brown longeared bats (Plecotus auritus) in bat boxes at Thetford Forest. Journal of Animal Ecology 26, 101–112. Boyles, J. G. (2007). Describing roosts used by forest bats: the importance of microclimate. Acta Chiropterologica 9, 297–303. doi: 10.3161/17335329(2007)9[297:DRUBFB]2.0.CO;2 Briggs, S. V. (1991). Intraspecific nest parasitism in maned ducks Chenonetta jubata. Emu 91, 230–235. Brittingham, M. C., and Williams, L. M. (2000). Bat boxes as alternative roosts for displaced bat maternity colonies. Wildlife Society Bulletin 28, 197–207. Use of artificial tree hollows by Australian birds and bats Burton, N. H. K. (2007). Intraspecific latitudinal variation in nest orientation among ground-nesting passerines: a study using published data. The Condor 109, 441–446. doi: 10.1650/0010-5422(2007)109[441: ILVINO]2.0.CO;2 Calder, T. G., Golding, B. G., and Manderson, A. D. (1983). Management for arboreal species in the Wombat State Forest. Environmental Report No. 16, Graduate School of Environmental Science, Monash University, Melbourne. Calver, M. C., and King, D. R. (1999). Why publication matters in conservation biology. Pacific Conservation Biology 6, 2–8. Campbell, S., Lumsden, L. F., Kirkwood, R., and Coulson, G. (2005). Day roost selection by female little forest bats (Vespadelus vulturnus) within remnant woodland on Phillip Island, Victoria. Wildlife Research 32, 183–191. doi: 10.1071/WR04039 Churchill, S. (1998). ‘Australian Bats.’ (New Holland: Sydney.) Commonwealth of Australia (2006). Background and implementation information for the South-eastern Red-tailed Black-Cockatoo, Calyptorhynchus banksii graptogyne recovery plan. Department of the Environment and Water Resources, Canberra. Commonwealth of Australia (2007). National recovery plan for the Southeastern Red-tailed Black-Cockatoo Calyptorhynchus banksii graptogyne. Department of the Environment and Water Resources, Canberra. Courtney, J., and Debus, S. J. S. (2006). Breeding habits and conservation status of the musk lorikeet Glossopsitta concinna and the little lorikeet G. pusilla in northern New South Wales. Australian Field Ornithology 23, 109–124. Dawson, R. D., Lawrie, C. C., and O’Brien, E. L. (2005). The importance of microcimalte variation in determining size, growth and survival of avian offspring: experimental evidence from a cavity nesting passerine. Oecologia 144, 499–507. doi: 10.1007/s00442-005-0075-7 Eadie, J., Sherman, P., and Semel, B. (1998). Conspecific brood parasitism, population dynamics, and the conservation of cavity-nesting birds. In ‘Behavioral Ecology and Conservation Biology’. (Ed. T. Caro.) pp. 306–340. (Oxford University Press: New York.) Emison, W. B. (1996). Use of supplementary nest hollows by an endangered subspecies of Red-tailed Black-cockatoo. Victorian Naturalist 113, 262–263. Evans, M. R., Lank, D. B., Boyd, W. S., and Cooke, F. (2002). A comparison of the characteristics and fate of Barren’s goldeneye and bufflehead nests in nest boxes and natural cavities. The Condor 104, 610–619. doi: 10.1650/0010-5422(2002)104[0610:ACOTCA]2.0.CO;2 Eyre, T. J. (2005). Hollow-bearing trees in large glider habitat in south-east Queensland, Australia: abundance, special distribution and management. Pacific Conservation Biology 11, 23–37. Flaquer, C., Torre, I., and Ruiz-Jarillo, R. (2006). The value of bat-boxes in the conservation of Pipistrellus pygmaeus in wetland rice paddies. Biological Conservation 128, 223–230. doi: 10.1016/j.biocon.2005. 09.030 Flaquer, C., Torre, I., and Arrizabalaga, A. (2007). Comparison of sampling methods for inventory of bat communities. Journal of Mammalogy 88, 526–533. doi: 10.1644/06-MAMM-A-135R1.1 Franks, A., and Franks, S. (2003). ‘Nest Boxes for Wildlife: A Practical Guide.’ (Bloomings Books: Melbourne.) Garnett, S. T., Pedler, L. P., and Crowley, G. M. (1999). The breeding biology of the glossy black-cockatoo Calyptorhynchus lathami on Kangaroo Island, South Australia. Emu 99, 262–279. doi: 10.1071/MU99032 Gibbons, P., and Lindenmayer, D. (2002). ‘Tree Hollows and Wildlife Conservation in Australia.’ (CSIRO Publishing: Melbourne.) Gibbons, P., Lindenmayer, D. B., Barry, S. C., and Tanton, M. T. (2002). Hollow selection by vertebrate fauna in forests of southeastern Australia and implications for forest management. Biological Conservation 103, 1–12. doi: 10.1016/S0006-3207(01)00109-4 Gleeson, J. A. (1999). Breeding observations of crimson and eastern rosellas in suburban nestboxes. Canberra Bird Notes 24, 167–172. Wildlife Research 95 Golding, B. G. (1979). Use of artificial hollows by mammals and birds in the Wombat Forest, Daylesford, Victoria. M.Sc. Thesis, Monash University, Melbourne. Goldingay, R. L., Grimson, M. J., and Smith, G. C. (2007). Do feathertail gliders show a preference for nest box design? Wildlife Research 34, 484–490. doi: 10.1071/WR06174 Grant, J. (1997). ‘The Nestbox Book.’ (Gould League: Melbourne.) Harley, D. K. P. (2004). Patterns of nest box use by Leadbeater’s possum (Gymnobelideus leadbeateri): applications to research and conservation. In ‘The Biology of Australian Possums and Gliders’. (Eds R. L. Goldingay and S. M. Jackson.) pp. 318–329. (Surrey Beatty and Sons: Sydney.) Harley, D. K. P. (2006). A role for nest boxes in the conservation of Leadbeater’s possum (Gymnobelideus leadbeateri). Wildlife Research 33, 385–395. doi: 10.1071/WR04038 Harper, M. J., McCarthy, M. A., and van der Ree, R. (2005a). The abundance of hollow-bearing trees in urban dry sclerophyll forest and the effect of wind on hollow development. Biological Conservation 122, 181–192. doi: 10.1016/j.biocon.2004.07.003 Harper, M. J., McCarthy, M. A., and van der Ree, R. (2005b). The use of nest boxes in urban natural vegetation remnants by vertebrate fauna. Wildlife Research 32, 509–516. doi: 10.1071/WR04106 Heinsohn, R., Murphy, S., and Legge, S. (2003). Overlap and competition for nest holes among eclectus parrots, palm cockatoos and sulphur-crested cockatoos. Australian Journal of Zoology 51, 81–94. doi: 10.1071/ ZO02003 Herr, A., and Klomp, N. I. (1999). Preliminary investigation of the roosting habitat preferences of the large forest bat Vespadelus darlingtoni (Chiroptera, Vespertilionidae). Pacific Conservation Biology 5, 208–213. Hill, R. (2002). Recovery plan for the Norfolk Island Green Parrot (Cyanoramphus novaezelandiae cookii). Environment Australia, Canberra. Homan, P. (1999). A fauna survey of riparian and other revegetation sites in Eltham, Victoria. Victorian Naturalist 116, 19–25. Homan, P. (2000). Excluding the common myna Acridotheres tristis from artificial nest boxes using a baffle. Victorian Nauralist 117, 75. Irvine, R., and Bender, R. (1995). Initial results from bat roosting boxes at Organ Pipes National Park. Victorian Naturalist 112, 212–217. Joseph, L., Emison, W. B., and Bren, W. M. (1991). Critical assessment of the conservation status of red-tailed black-cockatoos in south-eastern Australia with special reference to nesting requirements. Emu 91, 46–50. nig, B. (2001). Day roost selection in female Kerth, G., Weissmann, K., and Ko Bechstein’s bats (Myotis bechsteinii): a field experiment to determine the influence of roost temperature. Oecologia 126, 1–9. doi: 10.1007/ s004420000489 Koenig, W. D. (2003). European starlings and their effect on native cavitynesting birds. Conservation Biology 17, 1134–1140. doi: 10.1046/j.15231739.2003.02262.x Koenig, W. D., Gowaty, P. A., and Dickinson, J. L. (1992). Boxes, barns and bridges: confounding factors or exceptional opportunities in ecological studies. Oikos 63, 305–308. doi: 10.2307/3545392 Krebs, E. A. (1998). Breeding biology of crimson rosellas (Platycercus elegans) on Black Mountain, Australian Capital Territory. Australian Journal of Zoology 46, 119–136. doi: 10.1071/ZO97040 Krebs, E. A. (1999). Last but not least: nestling growth and survival in asynchronously hatching crimson rosellas. Journal of Animal Ecology 68, 266–281. doi: 10.1046/j.1365-2656.1999.00284.x Krebs, E. A. (2001). Begging and food distribution in crimson rosella (Platycercus elegans) broods: why don’t hungry chicks beg more? Behavioral Ecology and Sociobiology 50, 20–30. doi: 10.1007/ s002650100339 Krebs, E. A., and McGrath, R. D. (2000). Food allocation in crimson rosella broods: parents differ in their responses to chick hunger. Animal Behaviour 59, 739–751. doi: 10.1006/anbe.1999.1375 96 Wildlife Research Krebs, E. A., Cunningham, R., and Donnelly, C. F. (1999). Complex patterns of food allocation in asynchronously hatching broods of crimson rosellas. Animal Behaviour 57, 753–763. doi: 10.1006/ anbe.1998.1029 Krebs, E. A., Green, D. J., Double, M. C., and Griffiths, R. (2002). Laying date and laying sequence influence the sex ratio of crimson rosella broods. Behavioral Ecology and Sociobiology 51, 447–454. doi: 10.1007/s00265002-0459-1 Kunz, T. H., and Lumsden, L. F. (2003). Ecology of cavity and foliage roosting bats. In ‘Bat Ecology’. (Eds T. H. Kunz and M. B. Fenton.) pp. 3–89. (University of Chicago Press: Chicago, IL.) Lindenmayer, D. B., Cunningham, R. B., Tanton, M. T., and Smith, A. P. (1990). The conservation of arboreal marsupials in the montane ash forests of the Central Highlands of Victoria, south-east Australia: II. The loss of trees with hollows and its implications for the conservation of Leadbeater’s possum, Gymnobelideus leadbeateri McCoy (Marsupialia: Petauridae). Biological Conservation 54, 133–145. doi: 10.1016/0006-3207(90)90138-F Lindenmayer, D. B., Cunningham, R. B., and Donnelly, C. F. (1997). Decay and collapse of trees with hollows in eastern Australian forests: impacts on arboreal marsupials. Ecological Applications 7, 625–641. doi: 10.1890/ 1051-0761(1997)007[0625:DACOTW]2.0.CO;2 Lindenmayer, D. B., MacGregor, C. I., Cunningham, R. B., Incoll, R. D., Crane, M., Rawlins, D., and Michael, D. R. (2003). The use of nest boxes by arboreal marsupials in the forests of the central highlands of Victoria. Wildlife Research 30, 259–264. doi: 10.1071/WR02047 Loeb, S. C., and Hooper, R. G. (1997). An experimental test of interspecific competition for red-cockaded woodpecker cavities. The Journal of Wildlife Management 61, 1268–1280. doi: 10.2307/3802126 Lourenço, S. I., and Palmeirim, J. M. (2004). Influence of temperature in roost selection by Pipistrellus pygmaeus (Chiroptera): relevance for the design of bat boxes. Biological Conservation 119, 237–243. doi: 10.1016/ j.biocon.2003.11.006 Lumsden, H. G. (1989). Test of nest box preferences of eastern bluebirds, Sialia sialis, and tree swallows, Tachycineta bicolor. Canadian Field Naturalist 103, 595–597. Lumsden, L. F., Bennett, A. F., and Silins, J. E. (2002a). Selection of roost sites by the lesser long-eared bat (Nyctophilus geoffroyi) and Gould’s wattled bat (Chalinolobus gouldii) in south-eastern Australia. Journal of Zoology 257, 207–218. doi: 10.1017/S095283690200081X Lumsden, L. F., Bennett, A. F., and Silins, J. E. (2002b). Location of roosts of the lesser long-eared bat Nyctophilus geoffroyi and Gould’s wattled bat Chalinolobus gouldii in a fragmented landscape in southeastern Australia. Biological Conservation 106, 237–249. doi: 10.1016/ S0006-3207(01)00250-6 Lunney, D., Barker, J., and Priddel, D. (1985). Movements and day roosts of the chocolate wattled bat Chalinolobus morio (Gray) (Microchiroptera: Vespertilionidae) in a logged forest. Australian Mammalogy 8, 313–317. Lunney, D., Barker, J., and Leary, T. (1988). Movements of banded bats (Microchiroptera: Vespertilionidae) in Mumbulla State Forest near Bega, New South Wales. Australian Mammalogy 11, 167–169. Mackowski, C. M. (1984). The ontogeny of hollows in blackbutt, Eucalyptus pilularis, and its relevance to the management of forests for possums, gliders and timber. In ‘Possums and Gliders’. (Eds A. P. Smith and I. D. Hume.) pp. 517–525. (Australian Mammal Society: Sydney.) Marchant, S., and Higgins, P. J. (Eds) (1990). ‘Handbook of Australian, New Zealand and Antarctic Birds. Vol. 1. Ratites to Ducks.’ (Oxford University Press: Melbourne.) Mawson, P. R., and Long, J. L. (1994). Size and age parameters of nest trees used by four species of parrot and one species of cockatoo in south-western Australia. Emu 94, 149–155. McCulloch, E., and Thomas, R. (1986). ‘Nest Boxes for Australian Birds.’ (Bird Observers Club of Australia: Melbourne.) R. L. Goldingay and J. Stevens Menkhorst, P. W. (1984). Use of nest boxes by forest vertebrates in Gippsland: acceptance, preference and demand. Australian Wildlife Research 11, 255–264. doi: 10.1071/WR9840255 Milledge, D. (1978). A nest box used by the striated pardalote. Corella 2, 10–11. Mols, C. M. M., and Visser, M. E. (2002). Great tits can reduce caterpillar damage in apple orchards. Journal of Applied Ecology 39, 888–899. doi: 10.1046/j.1365-2664.2002.00761.x Mooney, P. A., and Pedler, L. P. (2005). Recovery plan for the South Australian subspecies of the Glossy Black-Cockatoo (Calyptorhynchus lathami halmaturinus): 2005–2010. South Australian Department for the Environment and Heritage, Adelaide. Neilson, A. L., and Fenton, M. B. (1994). Responses of little brown myotis to exclusion and to bat houses. Wildlife Society Bulletin 22, 8–14. Newton, I. (1994). The role of nest sites in limiting the numbers of hole-nesting birds: a review. Biological Conservation 70, 265–276. doi: 10.1016/00063207(94)90172-4 Norman, F. I. (1982). Eggs, egg-laying and incubation in the chestnut teal. Emu 82, 195–198. Norman, F. I., and Riggert, T. L. (1977). Nest boxes as nest sites for Australian waterfowl. The Journal of Wildlife Management 41, 643–649. doi: 10.2307/3799984 Nowak, R. M. (1999). ‘Walker’s Mammals of the World.’ 6th edn. (Johns Hopkins University Press: Baltimore, MD.) Olsen, P. D. (1996). Re-establishment of an endangered subspecies: the Norfolk Island boobook owl Ninox novaeseelandiae undulata. Bird Conservation International 6, 63–80. Orange-bellied Parrot Recovery Team (2006). Background and implementation information for the Orange-bellied Parrot recovery plan. Department of Primary Industries and Water, Hobart. Pedler, L. (1996). Artificial nest hollows for black-cockatoos. Eclectus 1, 13. Pell, A. S., and Tidemann, C. R. (1997a). The impact of two exotic hollow-nesting birds on two native parrots in savannah and woodland in eastern Australia. Biological Conservation 79, 145–153. doi: 10.1016/ S0006-3207(96)00112-7 Pell, A. S., and Tidemann, C. R. (1997b). The ecology of the common myna in urban nature reserves in the Australian Capital Territory. Emu 97, 141–149. doi: 10.1071/MU97018 Petty, S. J., Shaw, G., and Anderson, D. I. K. (1994). Value of nest boxes for population studies and conservation of owls in coniferous forests in Britain. The Journal of Raptor Research 28, 134–142. Pogue, D. W., and Schnell, G. D. (1994). Habitat characterization of secondary cavity-nesting birds in Oklahoma. The Wilson Bulletin 106, 203–226. Quin, B. R., and Baker-Gabb, D. J. (1993). Conservation and management of the turquoise parrot Neophema pulchella in north-east Victoria. Arthur Rylah Institute Technical Report Series No. 125, Victorian Department of Conservation and Environment, Melbourne. Radunzel, L. A., Muschitz, D. M., Bauldry, V. M., and Arcese, P. (1997). A long-term study of the breeding success of eastern bluebirds by year and cavity type. Journal of Field Ornithology 68, 7–18. Ruczynski, I. (2006). Influence of temperature on maternity roost selection by noctule bats (Nyctalus noctula) and Leisler’s bats (N. leisleri) in Bialowieza Primeval Forest, Poland. Canadian Journal of Zoology 84, 900–907. doi: 10.1139/Z06-060 Saunders, D. A. (1979). The availability of tree hollows for use as nest sites by white-tailed black cockatoos. Australian Wildlife Research 6, 205–216. doi: 10.1071/WR9790205 Saunders, D. A., Smith, G. T., and Rowley, I. (1982). The availability and dimensions of tree hollows that provide nest sites for cockatoos (Psittaciformes) in Western Australia. Australian Wildlife Research 9, 541–556. doi: 10.1071/WR9820541 Use of artificial tree hollows by Australian birds and bats Saunders, D. A., Smith, G. T., Ingram, J. A., and Forrester, R. I. (2003). Changes in a remnant of salmon gum Eucalyptus salmonophloia and York gum E. loxophleba woodland, 1978 to 1997. Implications for woodland conservation in the wheat-sheep regions of Australia. Biological Conservation 110, 245–256. doi: 10.1016/S0006-3207(02)00223-9 Schulz, M., and Lumsden, L. F. (2004). National recovery plan for the Christmas Island Pipistrelle Pipistrellus murrayi. Commonwealth of Australia, Canberra. Simpson, K., and Day, N. (1993). ‘Field Guide to the Birds of Australia.’ (Viking Penguin: Melbourne.) Smith, G. C., and Agnew, G. (2002). The value of ‘bat boxes’ for attracting hollow-dependent fauna to farm forestry plantations in southeast Queensland. Ecological Management & Restoration 3, 37–46. doi: 10.1046/j.1442-8903.2002.00088.x Soderquist, T. R., Traill, B. J., Faris, F., and Beasley, K. (1996). Using nestboxes to survey for the Brush-tailed Phascogale Phascogale tapoatafa. Victorian Naturalist 113, 256–261. Stebbings, R. E., and Walsh, S. T. (1991). ‘Bat-boxes: a Guide to the History, Function, Construction and Use in the Conservation of Bats.’ (The Bat Conservation Trust: London.) Threatened Species Section (2006). Fauna recovery plan: forty-spotted pardalote 2006–2010. Department of Primary Industries and Water, Hobart. Trainor, R. (1995a). Artificial nest-hollows: report on the use of artificial hollows in the box-ironbark forest surrounding Maryborough, Central Victoria 1982–1995. Bird Observer 759, 5–7. Trainor, R. (1995b). Sweet danger: how feral bees compete with hollow-using birds and mammals. Bird Observer 751, 7–9. Wildlife Research 97 Turbill, C. (2006). Roosting and thermoregulatory behaviour of male Gould’s long-eared bats, Nyctophilus gouldi: energetic benefits of thermally unstable tree roosts. Australian Journal of Zoology 54, 57–60. doi: 10.1071/ZO05068 Turbill, C., Kortner, G., and Geiser, F. (2003a). Natural use of heterothermy by a small, tree-roosting bat during summer. Physiological and Biochemical Zoology 76, 868–876. doi: 10.1086/378915 Turbill, C., Law, B. S., and Geiser, F. (2003b). Summer torpor in a free-ranging bat from subtropical Australia. Journal of Thermal Biology 28, 223–226. doi: 10.1016/S0306-4565(02)00067-0 Ward, S. J. (2000). The efficacy of nest boxes versus spotlighting for detecting feathertail gliders. Wildlife Research 27, 75–79. doi: 10.1071/WR99018 Willner, G. R., Gates, J. E., and Devlin, W. J. (1983). Nest box use by cavity nesting birds. American Midland Naturalist 109, 194–201. doi: 10.2307/ 2425530 Woinarski, J. C. Z., and Bulman, C. (1985). Ecology and breeding biology of the forty-spotted pardalote and other pardalotes on North Bruny Island. Emu 85, 106–120. Wood, M. S., and Wallis, R. L. (1998). Potential competition for nest boxes between feral honeybees and sugar gliders at Tower Hill State Game Reserve. Victorian Naturalist 115, 78–80. Zinner, D., Hilgartner, R. D., Kappeler, P. M., Pietsch, T., and Ganzhorn, J. U. (2003). Social organization of Lepilemur ruficaudatus. International Journal of Primatology 24, 869–888. doi: 10.1023/A:1024684907250 Manuscript received 1 May 2008, accepted 9 October 2008 http://www.publish.csiro.au/journals/wr