Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
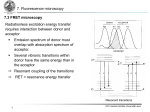
6. Fluorescence Spectroscopy 1 IPC Friedrich-Schiller-Universität Jena 6. Basic concepts in fluorescence spectroscopy 6.1 Stokes-Shift = Stokes-Shift due to vibrational energy relaxation within electronic excited state Energy differences between vibrational states which determine vibronic band intensities are very often the same for ground and electronic excited state Emission spectrum = mirror image of absorption spectrum Emission bands are shifted bathochromically i.e. to higher wavelengths 2 IPC Friedrich-Schiller-Universität Jena 6. Basic concepts in fluorescence spectroscopy 6.2 Fluorescence life-time The following transitions will be considered: S1 T1 S0 : rate constant for radiative S1S0 decay via fluorescence; : rate constant for internal conversion (S1S0); : rate constant for intersystem crossing; : rate constant for radiative decay via phosphorescence (T1S0); : rate constant for non-radiative decay (T1S0). Non radiative transitions originating from S1 are combined in: 3 IPC Friedrich-Schiller-Universität Jena 6. Basic concepts in fluorescence spectroscopy 6.2 Fluorescence life-time Dilute solution of fluorescent species A. Short d-laser pulse excites certain fraction of molecules A at t = 0. Decay rate of excited molecules A*: Integration: together with: number of molecules A promoted in the excited state at t = 0 and life-time of excited state S1: Fluorescence intensity is number of photons emitted per time and volume: 1A* 1A + Photon Fluorescence intensity IF at time t after excitation by a short light pulse: Part of molecules can end up in triplet state. Life-time of triplet state is defined as: 4 IPC Friedrich-Schiller-Universität Jena 6. Basic concepts in fluorescence spectroscopy 6.3 Fluorescence quantum yield Fluorescence quantum yield: Emitted Photons per Excitation events It follows: Integration over complete decay The quantum yields for ISC and phosphorescence can be expressed in analogy: bzw. 5 IPC Friedrich-Schiller-Universität Jena 6. Basic concepts in fluorescence spectroscopy 6.3 Fluorescence quantum yield Life-times & quantum yields Attention: Quantum yield is proportional to life-time but other non-radiative decay processes change lifetime radiative rate depends on refractive index of medium 6 IPC Friedrich-Schiller-Universität Jena 6. Basic concepts in fluorescence spectroscopy 6.4 Steady-State fluorescence emission It is advantageous to define the steady-state fluorescence per absorbed photon as photon flux in dependence of wavelength (Photon spectrum) : Emission photon spectrum expresses the probability distribution of the different transitions from the vibrational ground state of S1 down to the various vibrational states of S0. The normalized steady-state fluorescence IF(lF), recorded for the wavelength lF is proportional to as well as to the number of absorbed photons at the excitation wavelength lE. Number of absorbed photons is given by: Intensity transmitted irradiated 7 IPC Friedrich-Schiller-Universität Jena 6. Basic concepts in fluorescence spectroscopy 6.4 Steady-State fluorescence emission Fluorescence intensity can be expressed as follows: with k = proportionality constant dependent on numerous experimental values like e.g. collection angle, band width of monochromator, slid width, etc. Considering the intensity of the transmitted light by Lambert-Beer‘s law yields: Recording the intensity IF as function of the wavelength lF for a fixed excitation wavelength lE yields fluorescence spectrum. For low concentrations it follows: Higher terms can be neglected for diluted solutions. Thus it follows: A(lE) = absorbance at lE Proportionality between fluorescence intensity and concentration for diluted solutions only 8 IPC Friedrich-Schiller-Universität Jena 6. Basic concepts in fluorescence spectroscopy 6.4 Fluorescence excitation spectroscopy Recording fluorescence intensity as function of excitation wavelength lE for a fixed observation wavelength lF yields fluorescence excitation spectrum. According to: the fluorescence intensity recorded as a function of the excitation wavelength reflects the product In case the wavelength dependency of the incoming light can be compensated the fluorescence excitation spectrum depends only on what corresponds to the absorption spectrum. As long as only one ground state species exists the corrected excitation spectrum is identical to the absorption spectrum. Otherwise a comparison between fluorescence excitation and absorption spectrum yields valuable information about the sample species present. 9 IPC Friedrich-Schiller-Universität Jena 6. Basic concepts in fluorescence spectroscopy 6.4 Fluorescence excitation spectroscopy Fluorescence excitation (lE = variabel, lF = 419 nm) Absorption-, 210 CH3 N O fluorescence- and fluorescenceexcitation spectra N O COOH 415 359.5 IF(lE=359 nm,lF = variable) 350 267.5 245 Absorbance, A(lE) O 250 300 350 400 450 500 550 Fluorescence intensity, IF(lE = 359 nm, lF) 259 Cinoxacin in H2O 600 Wavelength / nm 10 IPC Friedrich-Schiller-Universität Jena 7. Fluorescence microscopy 7.1 Fluorochromes Fluorescence microscopy differentiates between two kinds of fluorochromes: Primary fluorescence (autofluorescence) Secondary fluorescence (fluorochromation) Fluorescence dyes Immunofluorescence (using Antibodies) Molecular tags (SNAP Tag, ...) Fluorescent Proteins Applications of fluorochromes Identification of otherwise visible structures Localization and identification of otherwise invisible structures Monitoring of physiological processes Specific detection of a protein Using Photophysical properties of dyes (e.g. switching) for superresolution 11 IPC Friedrich-Schiller-Universität Jena 7. Fluorescence microscopy 7.1 Primary fluorescence (autofluorescence) Most samples fluoresce when excited with short-wave light Fluorescence very often occurs for systems containing many conjugated double bonds: e.g. chlorophyll exhibits dark red fluorescence when excited by blue or red light N H N N H N Porphyrin ring – central unit in Chlorophyll 12 Moss reeds – green excitation IPC Friedrich-Schiller-Universität Jena 7. Fluorescence microscopy http://en.wikipedia.org/wiki/File:Chlorophyll_ab_spectra2.PNG 13 IPC Friedrich-Schiller-Universität Jena 7. Fluorescence microscopy 7.1 Primary fluorescence (autofluorescence) Further examples: Riboflavine (550nm) NAD(P)H (460nm, 400ps) Elastin und Collagen (305-450nm) Retinol (500nm) Cuticula (blue) Lignin (> 590nm) Eucalyptus leaf section – UV excitation DNA (Ex @320nm, 390nm) Aminoacids: Tryptophane (348nm, 2.6ns) Tyrosin (303nm, 3.6ns, weak) Phenylalanine (282nm weak) Resins, Oils http://en.wikipedia.org/wiki/Autofluorescence 14 Nematode living sample – UV excitation IPC Friedrich-Schiller-Universität Jena 7. Fluorescence microscopy 7.1 Secondary fluorescence (fluorochromation) Staining (labeling) specific structures with fluorescent labels (dyes): fluorochromation Small dye concentrations are sufficient due to high fluorescence contrast fluorescence labels are superior than bright field dyes Single molecule sensitivity Fluorescence labels must selectively bind to structures or selectively accumulate in specific compartments e.g. DAPI (= 4',6-diamidino-2-phenylindole) to label DNA (cell nuclei) DAPI: lexc = 358 nm lem = 461 nm 15 Fluorescence image of Endothelium cells. Microtubili are labeld in green, while actin filaments are labeled red. DNA within cell nuclei are stained with DAPI. IPC Friedrich-Schiller-Universität Jena 7. Fluorescence microscopy 7.3 FRET microscopy Radiationless excitation energy transfer requires interaction between donor and acceptor Emission spectrum of donor must overlap with absorption spectrum of acceptor. Several vibronic transitions within donor have the same energy than in the acceptor Resonant coupling of the transitions RET = resonance energy transfer Resonant transitions 16 IPC Friedrich-Schiller-Universität Jena 7. Fluorescence microscopy 7.3 FRET microscopy Radiationless excitation energy transfer Assumption: 2 electrons one at the donor D and one at the acceptor A are involved in the transition: Antisymmetric wavefunction (Fermions) for initially excited state i (D excited, but not A) and final state f (A excited, but not D): Overall Hamiltonian: Interaction energy: Coulomb term UC 17 Exchange term Uex IPC Friedrich-Schiller-Universität Jena 7. Fluorescence microscopy 7.3 FRET microscopy Radiationless excitation energy transfer Coulomb Interaction (CI) Exchange Interaction 18 IPC Friedrich-Schiller-Universität Jena 7. Fluorescence microscopy 7.3 FRET microscopy Radiationless excitation energy transfer Different interaction mechanism lead to excitation energy transfer: Dipolar (Förster) „Long Range“ Coulomb interaction Multipolar Singlet energy transfer Inter molecular orbital overlap Triplet Energy transfer 19 Electron exchange (Dexter) „Short Range“ Charge resonance interaction IPC Friedrich-Schiller-Universität Jena 7. Fluorescence microscopy 7.3 FRET microscopy Förster Resonance Energy Transfer (very weak coupling): D + hn1 D* + A A* D* Absorption A* + D Energy transfer A + hn2 Emission The following conditions must hold: D must be a fluorophore with sufficiently long life-time Partial spectral overlap between emission spectrum of D and absorption spectrum of A Transition dipole moments D and A must be oriented properly to each other; Distance between D and A shouldn‘t be too large 20 IPC Friedrich-Schiller-Universität Jena 7. Fluorescence microscopy 7.3 FRET microscopy Förster Resonance Energy Transfer (very weak coupling): Coulomb interaction can be developed in a multipole series in which the dipole term exhibits the term with the longest range Energy transfer via dipole-dipole transfer has been first calculated by Förster and is therefore called Förster process Energy transfer rate from molecule D to molecule A at a distance r: kD = radiative decay rate of donor tD0 = donor life-time in absence of energy transfer r-6-dependency as a result of dipole-dipole interaction R0 = critical distance or Förster-radius (distance at which intensity decrease caused by energy transfer and spontaneous decay are equal ( = kD)). 21 IPC Friedrich-Schiller-Universität Jena 7. Fluorescence microscopy 7.3 FRET microscopy Förster Resonance Energy Transfer (very weak coupling): R0 can be determined via spectroscopic values: Overlap between fluorescence of donor and absorption of acceptor k2 = orientational factor F0D = quantum yield of donor in absence of energy transfer n = average refractive index for wavelength area of spectral overlap ID(l) = normalized fluorescence spectrum of donor ( ) eA(l) = molar absorption coefficient of acceptor. For R0 in Å, l in nm, eA(l) in M-1 cm-1 (overlap integral in M-1 cm-1 nm4) Typical values for Förster-radii R0, i.e. for distances, over which energy transfer is important lie in the range of 15 -60 Å 22 IPC Friedrich-Schiller-Universität Jena 7. Fluorescence microscopy 7.3 FRET microscopy Förster Resonance Energy Transfer (very weak coupling): Transfer efficiency can be expressed by: In combination with changed lifetime: 1 D 1 D0 k Add D distance dependency: It follows: D und D0 are excited state life-times of donor in absence and presence of acceptor, respectively 23 IPC Friedrich-Schiller-Universität Jena 7. Fluorescence microscopy 7.3 FRET microscopy Förster Resonance Energy Transfer (very weak coupling): Besides the distance between the two chromophores also the relative orientation of the transition dipole moments of the donor D and acceptor A plays a crucial role for the energy transfer efficiency The orientation factor k2 is given by: A: angle between D-A connecting line and acceptor transition dipole moment D: angle between D-A connecting line and donor transition dipole moment T: angle between donor and acceptor transition dipole moment D A 24 IPC Friedrich-Schiller-Universität Jena 7. Fluorescence microscopy 7.3 FRET microscopy Förster Resonance Energy Transfer (very weak coupling): For systems where the orientation stays constant during the energy transfer (e.g. usage of highly viscose solvents or rigid coupling of chromophores to large and stiff molecules) k2 can reach values between 0 (transition dipole moments are orthogonal) and 4 (collinear arrangement); k2 = 1, for a parallel arrangement If both acceptor and donor can rotate the orientational factor k2 must be replaced by an average value: In case both chromophores undergo a fast isotropic rotation i.e. the rotation is considerably faster than the energy transfer rate the average orientation factor is given by k2 = 2/3 In case donor and acceptor are freely movable but the rotation is significantly slower than the energy transfer the orientation factor results in: k2 = 0.476 25 IPC Friedrich-Schiller-Universität Jena 7. Fluorescence microscopy 7.3 FRET microscopy Förster Resonance Energy Transfer (very weak coupling): RET is utilized as „optical nano ruler“ (10 – 100 Å) in biochemistry and cell biology Distance between donor and acceptor should be in the range of: because R0 is a benchmark for donor-acceptor distances which can be determined by FRET. 26 IPC Friedrich-Schiller-Universität Jena 7. Fluorescence microscopy 7.3 FRET microscopy Förster Resonance Energy Transfer (very weak coupling): RET as „optical nano ruler“ in biochemistry and cell biology 27 IPC Friedrich-Schiller-Universität Jena 7. Fluorescence microscopy 7.3 FRET microscopy Förster Resonance Energy Transfer (very weak coupling): RET as „optical nano ruler“ in biochemistry and cell biology 28 IPC Friedrich-Schiller-Universität Jena 7. Fluorescence microscopy 7.3 FRET microscopy Förster Resonance Energy Transfer (very weak coupling): RET as „optical nano ruler“ in biochemistry and cell biology One requires appropriate method to label specific intracellular proteins with suitable fluorophores (fluorescent proteins genetics): Green Fluorescent Protein (GFP) first isolated from the jellyfish Aequorea victoria GFP can be combined with just about any other protein by attaching its gene to the gene of a target protein, thereby introducing it into a cell. Thus by recording the GFP fluorescence the spatial and temporal distribution of this target protein can be directly monitored in living cells, tissue and organism. Several GFP mutants with altered fluorescence spectra exist. These mutants are named according to their color e.g. CFP (cyan) or YFP (yellow) Excitation maxima at 395 und 475 nm Emission wavelength at 509 nm 29 IPC Friedrich-Schiller-Universität Jena 7. Fluorescence microscopy 7.3 FRET microscopy Agar plate of fluorescent bacteria colonies 30 IPC Friedrich-Schiller-Universität Jena 7. Fluorescence microscopy 7.3 FRET microscopy Förster Resonance Energy Transfer (very weak coupling): RET as „optical nano ruler“ in biochemistry and cell biology : GFP-mutants FRET R0 = 4.7 – 4.9 nm no FRET protein folding 31 protein-protein interaction IPC Friedrich-Schiller-Universität Jena 7. Fluorescence microscopy 7.3 FRET microscopy Resolution of a light microscope is limited to several hundred nanometers (< organelles) FRET allows detection of molecule-molecule interactions on a nanometer scale by means of a light microscope Decrease of donoremission Increase of acceptor emission Reduction of donor fluorescence life-time Energy transfer (FRET-efficiency) depends strongly on donor-acceptor distance R0 = Förster-radius (distance for which energy transfer is half maximal) sensitized emission 32 IPC Friedrich-Schiller-Universität Jena 7. Fluorescence microscopy 7.3 FRET microscopy FRET ratio imaging = acceptor emission at donor excitation (sensitized emission SAkzeptor) divided by donor emission at donor excitation (SDonor) SDonor SAkzeptor Advantages: Since both donor decrease as well as acceptor increase contribute to the signal the signal-to-noise ratio is better than for solely recording the acceptor fluorescence 33 IPC Friedrich-Schiller-Universität Jena 7. Fluorescence microscopy 7.3 FRET microscopy FRET ratio imaging – problems: Excitation wavelength FRET-detection channel Correction for direct excitation of Correction for bleedthrough : the acceptor when exciting donor Portion of CFP in yellow channel for blue excitation in absence of FRET (acceptor) (control measurement with YFP = bleedthrough of CFP in YFP-channel (rBT,CY) only) = correction factor rDE or bleedthrough of YFP in CFP-channel (rBT,YC) 34 IPC Friedrich-Schiller-Universität Jena 7. Fluorescence microscopy 7.3 FRET microscopy FRET ratio imaging – 3-filter-set: 1. Donor excitation and emission (ICFP,430) 2. Acceptor excitation and emission (IYFP,514) 3. Donor excitation and acceptor emission (IYFP,430) 35 IPC Friedrich-Schiller-Universität Jena 7. Fluorescence microscopy 7.3 FRET microscopy FRET ratio imaging – 3-filter-set: Model of FRET-detection of SrcCsk protein interaction (Src = protein tyrosine kinase Csk = C-terminal Src kinase) Important signal transduction step during blood coagulation 36 No FRET IPC Friedrich-Schiller-Universität Jena 7. Fluorescence microscopy 7.3 FRET microscopy FRET ratio imaging – 3-filter-set: Visualization of Src-Csk-interaction during aIIbß3-induced fibrinogen adhesion in a thrombocyte model cell line (A5-CHO) by means of FRET superposition 37 IPC Friedrich-Schiller-Universität Jena 7. Fluorescence microscopy 7.3 FRET microscopy FRET ratio imaging – 3-filter-set: FRET for displaying Ca2+ in living cells via Yellow-Cameleon-2 (YC2) sensor FRET-ratio image of HeLa-cells, expressing the YC2-sensor before and after adding ionomycin FRET response of HEK/293 cells expressing YC2-seonsor after adding 1nM ionomycin and additional extracellular Ca21 (30 mM) 38 IPC Friedrich-Schiller-Universität Jena 7. Fluorescence microscopy 7.3 FRET microscopy FRET fluorescence life-time microscopy: In case of FRET the donor fluorescence life-time is reduced. Determination of this donor life time reduction yields a quantitative FRET measurement which is independent of dye concentration or spectral contamination (crosstalk, bleedthrough). Dimerization of C/EBP® – proteins in GHFT1-5 cell nuclei (donor/acceptor CFP/YFP-C/EBP®) 39 IPC Friedrich-Schiller-Universität Jena 7. Fluorescence microscopy 7.4 Fluorescence Life-Time Imaging Microscopy (FLIM) Methods of time-resolved fluorescence diagnostics Time-resolved measurements Sample is excited by a short laser pulse Laser pulse Longer fluorescence life-time Shorter fluorescence life-time 40 Sample molecules relax individually according to the transition probability of the different relaxation pathways to the ground state Fluorescence intensity exhibits mono-, multi or non-exponential decay depending on nature and number of fluorescence contributions. IPC Friedrich-Schiller-Universität Jena 7. Fluorescence microscopy 7.4 Fluorescence Life-Time Imaging Microscopy (FLIM) Methods of time-resolved fluorescence diagnostics Time-resolved measurements Intensity integrating measurements The determination of the fluorescence decay time ¿ or times ¿i and relative amplitudes ®i in case of multiple contributions is possible by recording the fluorescence signal for several measurement points after the excitation pulse. For a mono-exponential decay behavior or to determine the average decay time ¿ two sampling points are sufficient 41 For two times t1 and t2 after the excitation pulse the detector signal is integrated for a sampling window ¢ T. The ratio of the measurement signals D1 und D2 can be used to calculate the decay-time ¿ or the average decay-time ¿ : IPC Friedrich-Schiller-Universität Jena 7. Fluorescence microscopy 7.4 Fluorescence Life-Time Imaging Microscopy (FLIM) Methods of time-resolved fluorescence diagnostics Time-resolved measurements Gated fluorescence detection Gated optical image intensifiers (GOI) are capable of taking pictures with high (subnanosecond) time resolution i.e. camera with ultrafast shutter (gate < 100 ps) which can be opened and closed for different delay-times after the sample has been excited with an ultrashort laser-pulse. By collecting a series of time-scanned fluorescence intensity images for different delay-times after excitation the fluorescence decay profile for every pixel in the field of view can be accessed and displayed as false color plot = fluorescence life-time image 42 IPC Friedrich-Schiller-Universität Jena 7. Fluorescence microscopy 7.4 Fluorescence Life-Time Imaging Microscopy (FLIM) Methods of time-resolved fluorescence diagnostics Time-resolved measurements Gated fluorescence detection (a) (b) (c) Tissue section of a rat ear: (a) Brightfield microscopy image stained with orcein (b) Fluorescence intensity- and (c) FLIM images of an unstained parallel sample (tissue autofluorescence) (excitation 410 nm; FLIM false color plot from 200 ps (blue) to 1800 ps (red) 43 (Top) FLIM image of an unstained human pancreas section (tissue autofluorescence) with an endocrine tumor (below) Brightfield image of the same section after conventional histopathological staining IPC Friedrich-Schiller-Universität Jena 7. Fluorescence microscopy 7.4 Fluorescence Life-Time Imaging Microscopy (FLIM) Methods of time-resolved fluorescence diagnostics Time-resolved measurements TCSPC = time-correlated single photon counting In case of intensive excitation light many electrons of the dye are getting excited for every laser pulse i.e. the average life-time can be deduced from the fluorescence decay-time after every pulse (multiple photon emission). A common FLIM method is the measurement of the life-time for single fluorescence photons. In doing so the dye is excited by light pulses of extremely low intensity in a way that at most one electron per pulse gets excited. The individual lifetime of every photon is measured and the average life-time is determined staistically. 44 IPC Friedrich-Schiller-Universität Jena 7. Fluorescence microscopy 7.4 Fluorescence Life-Time Imaging Microscopy (FLIM) Methods of time-resolved fluorescence diagnostics Time-resolved measurements TCSPC = time-correlated single photon counting Detection of single photons of a periodic light signal Light intensity is so weak, that the probability to detect a photon within one period is very small. Periods with more than one photon are extremely rare For every detected photon its delay time with respect to the excitation pulse is determined A delay distribution builds up over many pulse Time resolution up to 25 ps 45 IPC Friedrich-Schiller-Universität Jena 7. Fluorescence microscopy 7.4 Fluorescence Life-Time Imaging Microscopy (FLIM) Methods of time-resolved fluorescence diagnostics Time-resolved measurements TCSPC = time-correlated single photon counting Probe Stop watch = TAC: Time-to-Amplitude Converter converts time between a start and a stop pulse by charging a capacitor with constant current Start can be reference (from laser) and photon is stop -> Problem is loss of much time (due to reset time) -> Reverse counting (start = photon, stop=next laser pulse) Histogram of arrival times after excitation -> fluorescence life-time. 46 IPC Friedrich-Schiller-Universität Jena 7. Fluorescence microscopy 7.4 Fluorescence Life-Time Imaging Microscopy (FLIM) Methods of time-resolved fluorescence diagnostics Time-resolved measurements TCSPC = time-correlated single photon counting Fluorescence intensity image of a vacuole which is labeled by fluorescent phospholipids 47 FLIM image and corresponding distribution of life-times. Long life-times (red) are found in the cell membrane while the cytoplasma exhibits shorter life-times pointing towards a less ordered environment. IPC Friedrich-Schiller-Universität Jena 7. Fluorescence microscopy 7.4 Fluorescence Life-Time Imaging Microscopy (FLIM) Methods of time-resolved fluorescence diagnostics Steady state measurements Phase modulation Intensity of a continuous wave (CW) source is modulated at high frequency by a standing wave acousto-optic modulator (n 2 50 MHz) which will modulate the excitation intensity at double frequency. Detected fluorescence is modulated at the same frequency. The observed phase shift with respect to the excitation and the modulation depth M (ratio of Ac signal to DC signal) depends on the fluorescence life-time of the excited fluorophores. Fluorescence lifetimes phase and phase can be calculated and should be identical for single exponential decays. 48 IPC Friedrich-Schiller-Universität Jena 7. Fluorescence microscopy 7.4 Fluorescence Life-Time Imaging Microscopy (FLIM) Methods of time-resolved fluorescence diagnostics Intensity Steady state measurements Phase modulation Excitation light Measurement values: Demodulation (modulation depth) M Phase shift DF Modulation of excitation light with n, which is characterized by modulation depth ME = a/d and FE : Fluorescence Time 49 leads to an accordingly modulated fluorescence signal F(t) with demodulation MF = A/D und phase FF IPC Friedrich-Schiller-Universität Jena 7. Fluorescence microscopy 7.4 Fluorescence Life-Time Imaging Microscopy (FLIM) Methods of time-resolved fluorescence diagnostics Steady state measurements Phase modulation Rate equation of change of number of excited molecules Absorption rate: F(t) ~ N(t) Relationship between fluorescence life-time and fluorescence emission behavior upon intensity modulated excitation light Relationship between measurement parameters: M = MF / ME as well as DF = FE - FE and life-time : 50 IPC Friedrich-Schiller-Universität Jena 7. Fluorescence microscopy 7.4 Fluorescence Life-Time Imaging Microscopy (FLIM) Methods of time-resolved fluorescence diagnostics Steady state measurements Phase modulation Continuous intensity modulated excitation of fluorescence transforms the determination of fluorescence decay-times to measurements of phase shifts and demodulation of the fluorescence signal Demodulation M and phase shift DF of the fluorescence depend on the fluorescence lifetime as well as on the modulation frequency w = 2p n of the excitation light. The simulation shows the dependency of M and DF for a decay time of = 4 ns and a frequency of n = 40 MHz Choosing frequency at w 1 51 IPC Friedrich-Schiller-Universität Jena 7. Fluorescence microscopy 7.4 Fluorescence Life-Time Imaging Microscopy (FLIM) Methods of time-resolved fluorescence diagnostics Steady state measurements Phase modulation Frozen section of portio biopsies in the spectral region of lEm>500 nm (lexc = 457 nm; n = 40 MHz) Top right: Fluorescence intensity Bottom right: corresponding HE stain image. 52 IPC Friedrich-Schiller-Universität Jena