Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
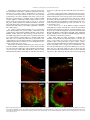
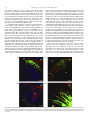
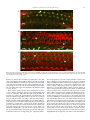
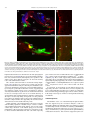
Neuroscience Letters 442 (2008) 134–139 Contents lists available at ScienceDirect Neuroscience Letters journal homepage: www.elsevier.com/locate/neulet Efficient and specific transduction of cochlear supporting cells by adeno-associated virus serotype 5 Ester Ballana a,1 , Jing Wang b , Frédéric Venail b , Xavier Estivill a,c , Jean-Luc Puel b , Maria L. Arbonès a,d , Assumpció Bosch e,∗ a Genes and Disease Programme, Center for Genomic Regulation (CRG), UPF, Barcelona, Catalonia, Spain INSERM U583, Montpellier, France Centre for Biomedical Research on Epidemiology and Public Health (CIBERESP), Barcelona, Catalonia, Spain d Centre for Biomedical Research on Rare Diseases (CIBERER), Barcelona, Catalonia, Spain e Department of Biochemistry and Molecular Biology, Center for Animal Biotechnology and Gene Therapy (CBATEG), Universitat Autònoma de Barcelona, E-08193 Bellaterra, Catalonia, Spain b c a r t i c l e i n f o Article history: Received 10 April 2008 Received in revised form 19 June 2008 Accepted 19 June 2008 Keywords: Gene transfer AAV5 serotype Inner ear Supporting cells a b s t r a c t Congenital deafness, affecting 1 in 1000 neonates, can lead to major problems in speech, cognitive and psychosocial development. Congenital deafness is mainly caused by mutations in connexins, hemi-channel proteins forming gap-junctions between supporting cells in the sensory epithelia. We describe a high tropism of AAV5 serotype for the supporting cells of the cochlea, both in vitro in postnatal day 4 mouse explants, and in vivo in the adult guinea-pig inner ear, through scala media perfusion. AAV5 transduction correlates with PDGFR␣ expression, previously reported as AAV5 receptor. This vector could be of major interest in addressing gene therapy approaches to deafness as well as for studying basic aspects of inner-ear development and hearing mechanisms. © 2008 Elsevier Ireland Ltd. All rights reserved. Hearing impairment is a clinical alteration affecting more than 250 million people worldwide. Among these, one child in a thousand before the age of two can suffer from congenital deafness, which may lead to significant problems in speech, cognitive and psychosocial development [21]. Sixty percent of these cases have a genetic cause and no efficient treatment has yet been found. Most of the pathologies leading to hearing impairment affect the inner ear, where only 15,000 neurosensorial cells – the hair cells – establish connections with 30,000 primary hearing neurons. However, the main cause of congenital deafness is due to mutations in connexins (CX), mainly expressed in the supporting cells of the cochlea [10,14]. Connexins are hemi-channel proteins forming gap-junctions between supporting cells in the sensory epithelia. In many populations, mutations in CX26 [14], CX30 [11] and in other CX genes [24] account for about half of the inherited prelingual non-syndromic cases of deafness. Despite recent surgical and technological advances, there is still no clinically efficient treatment for hearing disorders. Gene therapy could ∗ Corresponding author. Tel.: +34 935814203; fax: +34 935814200. E-mail address: [email protected] (A. Bosch). 1 Present address: IrsiCaixa Foundation, Hospital Universitari Germans Trias i Pujol, Badalona, Catalonia, Spain. 0304-3940/$ – see front matter © 2008 Elsevier Ireland Ltd. All rights reserved. doi:10.1016/j.neulet.2008.06.060 potentially offer a beneficial approach to solving this type of deafness. The inner ear offers several advantages as a model for gene therapy, as it is a small and well-compartmentalized receptacle that is easily accessible through retroauricular injection; additionally, cochlear endolymph and perilymph volumes are well characterized in humans and in animal models, so adverse effects of high volume and pressure can be avoided. Several publications have reported viral and non-viral transduction of the inner ear in rodents through different routes of administration, targeting a range of cell types with variable efficiencies [3,7,15,16,20,25]. Among the viral vectors available, adeno-associated virus (AAV) is one of the most promising gene therapy vectors for human clinical trials. Different serotypes of AAV vectors have shown high transduction specificity for certain cell types. For example, AAV1 and AAV5 mostly infect neurons in the central nervous system, and are able to diffuse into the brain parenchyma for several millimetres, while AAV4 infects ependymal cells [4,6]. In order to ascertain an AAV serotype with a high and specific tropism for supporting cells in the cochlea, we tested the infection capacity of different pseudotyped AAV vectors in mouse organotypic cochlear cultures, and in vivo in the adult guinea-pig inner ear and we characterized the cell types transduced in both models. E. Ballana et al. / Neuroscience Letters 442 (2008) 134–139 All animal procedures involved were previously approved by the Animal Ethics Committees of Catalonia and France and were performed in accordance with recommendations for the proper care and use of laboratory animals. Cochleae were dissected from postnatal day 3 CD-1 mice. Epithelia of the entire cochleae were infected in eppendorf tubes containing 20 l of culture medium and virus particles of AAV1, AAV2, AAV4 or AAV5 at a final concentration of 109 viral genomes/ml (vg/ml) for 12 h, at 37 ◦ C and 5% CO2 . Subsequently, explants were cultured on laminin-coated LabTek 8-well culture slides (Nalge Nunc) for 5, 10 or 15 days in DMEM supplemented with d-glucose (8.25 mM), l-glutamine (1 mM) and N1 (1:100) (Sigma). For cochlear explant immunostaining, we used antibodies against glial fibrillary acidic protein (GFAP) (1:400; Dako), Cx26 (1:200; Zymed), Neurofilament 200 kDa (NF200) (1:200, Sigma), PDGFR␣ (1:200; Santa Cruz Biotechnology), myelin basic protein (MBP) and peripherin (1:200; Chemicon). Primary antibodies were detected with secondary antibodies conjugated with Alexa Fluor 488 or 633 (1:1000; Molecular Probes) and cultures were counterstained for F-actin with rhodamine-conjugated phalloidin (100 mM; Sigma). Whole mounts were observed using fluorescence and/or confocal microscopy (Leica). Immunohistochemistry on adult guinea-pig cochleae was performed for PDGFR␣ on 10 m-thick cryo-sections. Double labeling was performed using antibodies against calbindin (1:100, Chemicon), myosin VIIa (1:200, ABR), and NF200 (1:800). Secondary labeling with Alexa Fluor 594 and 647 (1:500, Molecular Probes) 135 was used to localize these proteins with a Bio-Rad confocal microscope. For in vivo cochlear gene transfer, adult guinea-pigs were anaesthetized and endolymphatic perfusion was performed by shaving a small fenestrate in the cochlear bony capsule of the basal turn over the stria vascularis. A second aperture was made at the cochlear apex. Endocochlear potentials were recorded to ensure the correct positioning of the aperture. Only those animals with normal endocochlear potentials following this procedure (95.4 mV ± 1.1, n = 5) received AAV5 vectors. Vectors at titers of 3.7 × 109 vg diluted in 100 l of artificial endolymph were perfused through the scala media fenestration in the basal turn, and flowed out of the cochlea through the opening at the apex. Artificial endolymph composition was NaCl 1 mM, KCl 126 mM, KHCO3 25 mM, MgCl2 0.025 mM, CaCl2 0.025 mM, K2 HPO4 1.4 mM (pH 7.4; osmolarity = 300 mOsm/kg H2 O). Animals were euthanized 1 month after perfusion. AAV1, AAV2, AAV4 and AAV5 pseudotypes containing the CMVeGFP cassette flanked by AAV2-ITR sequences and the capside for each serotype were used to infect P4 mouse inner-ear explants at titers of 109 vg/ml. After 15 days in culture, cochleae counter-stained with phalloidin showed different patterns of transduction, depending on the serotype (Fig. 1). Fibroblast-like cells were infected with AAV2 (not shown) and AAV4 (Fig. 1C, arrows). AAV1 and, more efficiently, AAV2 were able to infect hair cells (Fig. 1A and B, arrowheads). In addition, and in accordance with that reported by other authors [16,20], AAV1 also infected supporting Fig. 1. Differential pattern of AAV pseudotyped-mediated transduction of mouse cochlea. Mouse P4 explants infected with AAV2/1 (A), AAV2/2 (B), AAV2/4 (C), or AAV2/5 (D) containing a CMVeGFP cassette (green) and counterstained with rhodamine-conjugated phalloidin (red) after 15 days in culture. Arrowheads indicate infected hair cells, arrows indicate fibroblast-like cells and asterisks indicate position of outer and inner hair cell rows at low magnification. OHC, outer hair cells; IHC, inner hair cells; HC, hair cells. Scale bar: 25 m. 136 E. Ballana et al. / Neuroscience Letters 442 (2008) 134–139 cells in the cochlea (Fig. 1A). In contrast, AAV5 showed hightransduction specificity for a particular structure located between the hair and the ganglion cells (Fig. 1D). Additional experiments performed with half-log lower titer AAV5 showed the same pattern of transduction at a similar efficiency (data not shown). With shorter virus incubation times (10 days), we obtained the exact transduction pattern. At 5 days, however, the level of GFP expression was too low to identify this structure (data not shown). Up to 40 cochleae were infected, with similar results. To identify the AAV5-transduced cells, we performed immunohistochemistry with antibodies targeting different types of inner-ear cell. We did not observe expression of GFP in immunoreactive cells for anti-peripherin (Fig. 2A) or anti-NF200 (Fig. 2B), which label type II and type I neurons, respectively; nor for anti-myosin VIIa (data not shown), which labels hair cells. GFPexpressing cells in the cochlea were always detected in the same structure in which GFAP-expressing supporting cells are located (Fig. 2C and D). In the mouse cochlea, all supporting cells express GFAP at P3 in a gradient decreasing in intensity from base to apex [19]. Interestingly, AAV5-transduced cells are also more numerous in the base than in the apex of the cochlea. To identify the type of supporting cells expressing GFP, we used anti-MBP and anti-Cx26 antibodies. These antibodies labeled most of the supporting cells interdigitated with hair cells in the sensory epithelia of the cochleae at time of infection (P3–P4) (Fig. 3A and B). Unfortunately, neither of these two antibodies worked in explants that had been in culture for several days, when GFP was evident. As many other viruses, AAV5 requires charged carbohydrates, in particular ␣-2,3-N-linked sialic acid, for efficient binding and transduction [13]. However, the only membrane receptor reported so far for AAV5 is PDGFR␣, described in the neurons of the rat hippocampus [8] and in the rods of the human retina [17]. Immunohistochemistry for PDGFR␣ performed in P4 mouse cochleae showed immunoreactivity in MBP-positive and Cx26-positive cells, consistent with the expression of the AAV5 receptor in most of the cochlear supporting cells (Fig. 3C). Our results therefore provide evidence that AAV5 is able to specifically infect the supporting cells of the cochlea in postnatal mice, probably through the PDGFR␣. Receptor expression can be speciesdependent, and can be modified throughout cochlear development; or it may be altered when a tissue is cultured for several days. To address these possibilities, we characterized the expression of PDGFR␣ in the adult guinea-pig, an animal model extensively used for inner-ear studies. We found PDGFR␣ immunoreactivity in different types of supporting cells, including Deiter’s, Hensen’s, pillar and the phalangeal cells of the guinea-pig organ of Corti (Fig. 4A). This result correlated with the in vitro studies carried out on mouse P4 inner ear. In addition, PDGFR␣ immunoreactivity was detected in spiral ganglion neurons and in the marginal cells of the stria vascularis in the adult guinea-pig (Fig. 4A and B). Expression of viral receptors is needed for vector-cell transduction; we therefore hypothesize that if we could access the supporting cells in the guinea-pig cochlea we would be able to transduce them with AAV5. Scala media perfusion in adult guinea-pigs of 3.7 × 109 vg/cochlea showed GFP expression in the supporting cells of the cochlea, mainly in Deiter’s, pillar and inner border cells, correlating with PDGFR␣ immunoreactivity (Fig. 4C). Fig. 2. AAV2/5 specifically transduces supporting cells of the mouse cochlea. Confocal images of cochlear explants processed for immunofluorescence at 10 days post-infection with AAV2/5-CMVGFP and labeled with anti-peripherin, labeling type II neurons (A), anti-NF200, labeling type I neurons (B) (both in blue) or anti-GFAP, labeling supporting cells (red) (C and D). A higher magnification of (C) is shown in (D). (A and B) were counterstained for F-actin with rhodamine-conjugated phalloidin (red). HC, hair cells. Scale bar: 25 m. E. Ballana et al. / Neuroscience Letters 442 (2008) 134–139 137 Fig. 3. Supporting cells in the neonatal mouse cochlea expressed PDGFR␣. Freshly isolated cochlea from P4 mice were processed for immunofluorescence using anti-MBP (A), anti-Cx26 (B) and anti-PDGFR␣ (C). All tissues were counterstained with rhodamine-conjugated phalloidin (red). Arrowheads indicate supporting cells. OHC, outer hair cells; IHC, inner hair cells. Scale bar: 25 m. However, no neurons were transduced, probably due to the difficulty of reaching the spiral ganglion neurons through scala media perfusion. In addition, some GFP expression was seen in marginal cells of the stria vascularis that also expressed PDGFR␣ (Fig. 4D). Sporadically, we were also able to target PDGFR␣-negative inner hair cells which may express an alternative, still unknown, AAV5 receptor (Fig. 4C). Other authors have reported AAV5 transduction in rodent cochleae, both in organotypic cultures from postnatal animals and in vivo, in adult mice, with different results [9,16,20]. While Di Pasquale et al. found only 1% transduction of vestibular hair cells in P2 rat explants using a CMV promoter [9], Stone et al. reported very rare transduced-hair cells using a chicken -actin promoter and rare transduced-interdental and Hensen cells with a GFAP promoter in mouse P1 organotypic cultures [20]. In vivo round window transduction of adult mouse cochleae using another ubiquitous promoter, chicken -actin, mostly showed infection of inner hair and spiral ganglion cells, and at lower levels, of inner sulcus, spiral limbus, and Claudius’ cells [16]. In these reports virus load was at least 10 times higher than in the present work with lower transduction efficiencies in supporting cells, suggesting that the increased tropism we report here for this particular cell type is not due to saturation of viral load. Moreover, we showed correlation between PDGFR␣ and AAV5 transduction, indicating specificity of AAV5 infection through its receptor in the supporting cells of the cochlea. Length of expression could also account for some of the differences reported. Other authors analyzed rodent explants at day 5 or at day 8 post-infection [9,20]; in contrast, we waited until day 15, described as the maximum for GFP expression in organotypic cultures of human airway epithelia [26]. We did not observe fluorescence in explants 5 days post-infection, indicating that supporting cells may need longer exposures to show AAV5-driven GFP expression. Similarly, we evaluated the in vivo experiments 4 weeks after transduction, whereas Liu et al. analyzed AAV5 expression at day 10 and recorded few transduced supporting cells [16]. Most of the in vivo experiments performed with AAV5 were evaluated at least 3 weeks after infection in a wide variety of tissues, including the CNS [4,6]. In the retina, AAV5 is one of the most efficient AAV serotypes and its expression starts at 2 weeks, although it peaks between 3 and 4 weeks [2,18]. Thus, it is possible that tissues processed at 10 days did not show enough reporter protein to highlight transduced supporting cells in the cochlea. Nevertheless, 138 E. Ballana et al. / Neuroscience Letters 442 (2008) 134–139 Fig. 4. Localization of PDGFR␣ in adult guinea-pig cochlea and transgene distribution after in vivo delivery of AAV2/5 in the inner ear target supporting cells. Immunohistochemistry for PDGFR␣ on adult guinea-pig cochlea. (A and B) Confocal images of transverse (midmodiolar) section through the medial turn in the adult guinea-pig cochlea, showing immunoreactivity to PDGFR␣ (red). In (A), calbindin counterstains hair cells in blue and NF200 counterstains nerve fibers of spiral ganglion neurons in green. PDGFR␣ was localized in the Deiter’s cells (DCs), Hensen’s cells (HCs), inner pillar cell (ip), outer pillar cell (op) and phalangeal and border cells (asterisks). PDGFR␣ immunoreactivity was also present in the marginal cells of the stria vascularis (sv) (B). (C and D) Confocal microscopy of sections of an AAV5-perfused adult guinea-pig cochlea at 1 month. GFP expression (green) was colocalized with myosin VIIa (red) in the inner hair cell (IHC) (C). GFP expression was also observed in the supporting cells of the organ of Corti such as Deiter’s cells (DCs) (C). GFP expression also occurred in the marginal cells of the stria vascularis (sv) (D). IHC, inner hair cell; OHCs, outer hair cells; sl, spiral ligament; nf, nerve fibers; isb, inner spiral bundles; tC, tunnel of Corti. Scale bar: 10 m. high transduction efficiencies in inner hair cells and spiral ganglion neurons were reported [16]. We also observed transduction in inner hair cells, but not in neurons, although they were positive for the AAV5 receptor. These differences may also be due to the delivery route or the administration volume. Mice models deficient in Cx26 or Cx30 show hearing impairment [5,22] and they are of interest for assaying gene therapy approaches with AAV5 coding for connexin genes. In addition, overexpression of Cx26 has been shown to correct Cx30-linked deafness by crossing two genetically modified mice models, suggesting that up-regulation of Cx26 could be used as treatment for deafness caused by Cx30 mutations [1]. However, it is not known when Cx26 needs to be overexpressed in order to correct Cx30 deficiency. A question that requires further attention is whether connexins may already be needed early during development or in the postnatal period. This issue might be addressed by specific administration of an AAV5 coding for Cx26 to the supporting cells of Cx26 or Cx30deficient mouse models at different time points. Unlike avian hair cells, mammalian hair cells do not regenerate and the initial population of 15,000 cells per cochlea is continuously diminished through frequent damage such as loud sounds, certain drugs, infections or the natural aging process. Recently, mammalian cochlear supporting cells have been shown to be able to divide and trans-differentiate into hair cells when expressing genes involved in hair-cell differentiation such as Math1/Atoh1 [12], or when cyclin-dependent kinase inhibitor p27Kip1 is downregulated [23]. These results suggest that postnatal mammalian supporting cells are potential targets for therapeutic manipulation. Specifically targeting supporting cells in the cochlea with AAV5 coding for Math1 or containing RNA interference sequences against p27Kip1 may therefore restore some of the lost hair cells in the cochlea. In conclusion, we demonstrate specific AAV5 tropism for the supporting cells of the cochlea both in vitro, in mouse explants, and in vivo, in the adult guinea-pig. This vector could be of considerable interest in addressing gene therapy approaches to deafness, but also in the study of basic aspects of inner-ear development and hearing mechanisms. Acknowledgements We thank the vector core of the University Hospital of Nantes that was supported by the Association Française contre les Myopathies (AFM) for providing the AAV vectors. EB was a recipient of a fellowship from the Generalitat de Catalunya (2003FI00066) and AB was an investigator of the Reincorporación de Doctores y Tecnólogos and the Ramon y Cajal Programs from the Spanish Ministry of Education and Science. This work was supported by the Gen- E. Ballana et al. / Neuroscience Letters 442 (2008) 134–139 eralitat de Catalunya (ACI2001/8), the Instituto de Salud Carlos III (G03/203, PI05-2347, PI05-1705) and the Fundació Marató de TV3 (98/1710). References [1] S. Ahmad, W. Tang, Q. Chang, Y. Qu, J. Hibshman, Y. Li, G. Sohl, K. Willecke, P. Chen, X. Lin, Restoration of connexin26 protein level in the cochlea completely rescues hearing in a mouse model of human connexin30-linked deafness, Proc. Natl. Acad. Sci. U.S.A. 104 (2007) 1337–1341. [2] A. Auricchio, G. Kobinger, V. Anand, M. Hildinger, E. O’Connor, A.M. Maguire, J.M. Wilson, J. Bennett, Exchange of surface proteins impacts on viral vector cellular specificity and transduction characteristics: the retina as a model, Hum. Mol. Genet. 10 (2001) 3075–3081. [3] J.C. Bedrosian, M.A. Gratton, J.V. Brigande, W. Tang, J. Landau, J. Bennett, In vivo delivery of recombinant viruses to the fetal murine cochlea: transduction characteristics and long-term effects on auditory function, Mol. Ther. 14 (2006) 328–335. [4] C. Burger, O.S. Gorbatyuk, M.J. Velardo, C.S. Peden, P. Williams, S. Zolotukhin, P.J. Reier, R.J. Mandel, N. Muzyczka, Recombinant AAV viral vectors pseudotyped with viral capsids from serotypes 1, 2, and 5 display differential efficiency and cell tropism after delivery to different regions of the central nervous system, Mol. Ther. 10 (2004) 302–317. [5] M. Cohen-Salmon, T. Ott, V. Michel, J.P. Hardelin, I. Perfettini, M. Eybalin, T. Wu, D.C. Marcus, P. Wangemann, K. Willecke, C. Petit, Targeted ablation of connexin26 in the inner ear epithelial gap junction network causes hearing impairment and cell death, Curr. Biol. 12 (2002) 1106–1111. [6] B.L. Davidson, C.S. Stein, J.A. Heth, I. Martins, R.M. Kotin, T.A. Derksen, J. Zabner, A. Ghodsi, J.A. Chiorini, Recombinant adeno-associated virus type 2, 4, and 5 vectors: transduction of variant cell types and regions in the mammalian central nervous system, Proc. Natl. Acad. Sci. U.S.A. 97 (2000) 3428–3432. [7] M.L. Derby, M. Sena-Esteves, X.O. Breakefield, D.P. Corey, Gene transfer into the mammalian inner ear using HSV-1 and vaccinia virus vectors, Hear Res. 134 (1999) 1–8. [8] G. Di Pasquale, B.L. Davidson, C.S. Stein, I. Martins, D. Scudiero, A. Monks, J.A. Chiorini, Identification of PDGFR as a receptor for AAV-5 transduction, Nat. Med. 9 (2003) 1306–1312. [9] G. Di Pasquale, A. Rzadzinska, M.E. Schneider, I. Bossis, J.A. Chiorini, B. Kachar, A novel bovine virus efficiently transduces inner ear neuroepithelial cells, Mol. Ther. 11 (2005) 849–855. [10] X. Estivill, P. Fortina, S. Surrey, R. Rabionet, S. Melchionda, L. D’Agruma, E. Mansfield, E. Rappaport, N. Govea, M. Mila, L. Zelante, P. Gasparini, Connexin-26 mutations in sporadic and inherited sensorineural deafness, Lancet 351 (1998) 394–398. [11] A. Grifa, C.A. Wagner, L. D’Ambrosio, S. Melchionda, F. Bernardi, N. Lopez-Bigas, R. Rabionet, M. Arbones, M.D. Monica, X. Estivill, L. Zelante, F. Lang, P. Gasparini, Mutations in GJB6 cause nonsyndromic autosomal dominant deafness at DFNA3 locus, Nat. Genet. 23 (1999) 16–18. [12] M. Izumikawa, R. Minoda, K. Kawamoto, K.A. Abrashkin, D.L. Swiderski, D.F. Dolan, D.E. Brough, Y. Raphael, Auditory hair cell replacement and hearing [13] [14] [15] [16] [17] [18] [19] [20] [21] [22] [23] [24] [25] [26] 139 improvement by Atoh1 gene therapy in deaf mammals, Nat. Med. 11 (2005) 271–276. N. Kaludov, K.E. Brown, R.W. Walters, J. Zabner, J.A. Chiorini, Adeno-associated virus serotype 4 (AAV4) and AAV5 both require sialic acid binding for hemagglutination and efficient transduction but differ in sialic acid linkage specificity, J. Virol. 75 (2001) 6884–6893. D.P. Kelsell, J. Dunlop, H.P. Stevens, N.J. Lench, J.N. Liang, G. Parry, R.F. Mueller, I.M. Leigh, Connexin 26 mutations in hereditary non-syndromic sensorineural deafness, Nature 387 (1997) 80–83. A.K. Lalwani, B.J. Walsh, P.G. Reilly, N. Muzyczka, A.N. Mhatre, Development of in vivo gene therapy for hearing disorders: introduction of adenoassociated virus into the cochlea of the guinea pig, Gene Ther. 3 (1996) 588– 592. Y. Liu, T. Okada, K. Sheykholeslami, K. Shimazaki, T. Nomoto, S. Muramatsu, T. Kanazawa, K. Takeuchi, R. Ajalli, H. Mizukami, A. Kume, K. Ichimura, K. Ozawa, Specific and efficient transduction of Cochlear inner hair cells with recombinant adeno-associated virus type 3 vector, Mol. Ther. 12 (2005) 725–733. A.J. Lotery, G.S. Yang, R.F. Mullins, S.R. Russell, M. Schmidt, E.M. Stone, J.D. Lindbloom, J.A. Chiorini, R.M. Kotin, B.L. Davidson, Adeno-associated virus type 5: transduction efficiency and cell-type specificity in the primate retina, Hum. Gene Ther. 14 (2003) 1663–1671. J.E. Rabinowitz, F. Rolling, C. Li, H. Conrath, W. Xiao, X. Xiao, R.J. Samulski, Crosspackaging of a single adeno-associated virus (AAV) type 2 vector genome into multiple AAV serotypes enables transduction with broad specificity, J. Virol. 76 (2002) 791–801. C. Rio, P. Dikkes, M.C. Liberman, G. Corfas, Glial fibrillary acidic protein expression and promoter activity in the inner ear of developing and adult mice, J. Comp. Neurol. 442 (2002) 156–162. I.M. Stone, D.I. Lurie, M.W. Kelley, D.J. Poulsen, Adeno-associated virusmediated gene transfer to hair cells and support cells of the murine cochlea, Mol. Ther. 11 (2005) 843–848. M. Tekin, K.S. Arnos, A. Pandya, Advances in hereditary deafness, Lancet 358 (2001) 1082–1090. B. Teubner, V. Michel, J. Pesch, J. Lautermann, M. Cohen-Salmon, G. Sohl, K. Jahnke, E. Winterhager, C. Herberhold, J.P. Hardelin, C. Petit, K. Willecke, Connexin30 (Gjb6)-deficiency causes severe hearing impairment and lack of endocochlear potential, Hum. Mol. Genet. 12 (2003) 13–21. P.M. White, A. Doetzlhofer, Y.S. Lee, A.K. Groves, N. Segil, Mammalian cochlear supporting cells can divide and trans-differentiate into hair cells, Nature 441 (2006) 984–987. J.H. Xia, C.Y. Liu, B.S. Tang, Q. Pan, L. Huang, H.P. Dai, B.R. Zhang, W. Xie, D.X. Hu, D. Zheng, X.L. Shi, D.A. Wang, K. Xia, K.P. Yu, X.D. Liao, Y. Feng, Y.F. Yang, J.Y. Xiao, D.H. Xie, J.Z. Huang, Mutations in the gene encoding gap junction protein beta-3 associated with autosomal dominant hearing impairment, Nat. Genet. 20 (1998) 370–373. T. Yamasoba, M. Yagi, B.J. Roessler, J.M. Miller, Y. Raphael, Inner ear transgene expression after adenoviral vector inoculation in the endolymphatic sac, Hum. Gene Ther. 10 (1999) 769–774. J. Zabner, M. Seiler, R. Walters, R.M. Kotin, W. Fulgeras, B.L. Davidson, J.A. Chiorini, Adeno-associated virus type 5 (AAV5) but not AAV2 binds to the apical surfaces of airway epithelia and facilitates gene transfer, J. Virol. 74 (2000) 3852–3858.