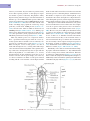
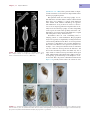
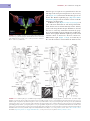
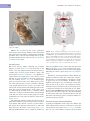
Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Chapter 13 Phylum Rotifera Robert L. Wallace Department of Biology, Ripon College, Ripon, Wisconsin, USA Terry W. Snell School of Biology, Georgia Institute of Technology, Atlanta, Georgia, USA Hilary A. Smith Department of Biological Sciences, University of Notre Dame, Notre Dame, Indiana, USA Chapter Outline Introduction to Rotifera 225 General Characteristics 225 Biogeography228 Evolutionary Relationships 230 General Biology 231 External Morphology 231 Organ System Structure and Function 232 Corona232 Trophi and Gut 232 Muscular System 235 Neural System 236 Excretory System: Protonephridium 236 Reproductive System 237 Environmental Physiology 238 Locomotion238 Physiological Ecology 238 Environmental Toxicology 240 Anhydrobiosis240 Generalized Stress Responses 242 Ecology and Evolution 242 Diversity and Distribution 242 Phenotypic Variation 242 INTRODUCTION TO ROTIFERA General Characteristics Phylum Rotifera comprises approximately 2000 species of unsegmented, bilaterally symmetrical invertebrates, most of which are found in freshwaters (Clément and Wurdak, 1991; Distribution and Population Movements 243 Colonial Rotifers 244 Sessile Rotifers 246 Reproduction and Life History 249 Reproduction249 Aging252 Population Dynamics 253 Ecological Interactions 258 Foraging Behavior 258 Functional Role in the Ecosystem 259 Competition with Other Zooplankton 261 Predator–Prey Interactions 261 Parasitism on Rotifers 264 Rotifers as Parasites 265 Collecting, Culturing, and Preparation for Identification 265 Collecting265 Culturing266 Laboratory Culture 266 Aquaculture267 Preparation for Identification 268 References269 Wallace et al., 2006; Segers, 2007). Their size ranges from 40 to 2000 μm, the smallest being only about 6 times the diameter of a human red blood cell. Because of their size, shape, and habitat, rotifers can be confused with protozoans (protists) (Chapter 7) and gastrotrichs (Chapter 12), but those taxa do not possess jaws and their ciliation is not distributed in the Thorp and Covich’s Freshwater Invertebrates. http://dx.doi.org/10.1016/B978-0-12-385026-3.00013-9 Copyright © 2015 Elsevier Inc. All rights reserved. 225 SECTION | III 226 same way as in rotifers. Very few rotifers are parasitic; nearly all are either raptorial predators or microphagous suspension feeders or grazers. Collectively this phylum is widely dispersed, being found in all types of freshwater habitats at densities up to about 1000 individuals per liter. With sufficient food, populations may surpass 5000 individuals per liter (Feike and Heerkloss, 2009). In some rather unusual water bodies, exceedingly large populations can develop; sewage ponds may contain about 12,000 per liter (Seaman et al., 1986), and soda water bodies in Chad can hold well over 100,000 per liter (Iltis and Riou-Duvat, 1971). Similar populations can be obtained in small chemostat systems (Boraas, 1983), but aquaculture systems population levels above 107 individuals per liter have been reported (Park et al., 2001). These tiny animals possess two conspicuous features. First, a specialized ciliated region called the corona (L., crown) caps the anterior end. The corona is commonly composed of two concentric rings of cilia (Figures 13.1–13.2). When viewing the corona of many species, one often is struck with the impression of a rotating wheel. This image comes from the metachronal (rhythmic and sequential) beating of their cilia, and inspired early microscopists with the name for the phylum (L., rota, wheel and L., ferre, to bear): the wheel-bearers. In free-swimming species the corona is used in locomotion, but all species employ it in some way to collect food. However, in adults of some species ciliation is lacking and the corona is funnel or bowl-shaped, with the Protozoa to Tardigrada mouth located at the bottom. The second obvious feature that all rotifers possess is a muscular pharynx, termed the mastax, that includes a complex set of jaws called trophi (G., troph, nourish). In some rotifers, the trophi are so unique that taxonomists distinguish species by critical morphological features of these minute structures (see the section “Trophi and Gut”). Although most rotifers inhabit freshwaters, some genera also have members that occur in saline waters. For example, 21 of the 39 species in the genus Synchaeta are known to occur in brackish to full-strength marine waters (Segers, 2007). However, only about 100 species distributed among 22 genera in the phylum are found exclusively in marine habitats (Ricci and Fontaneto, 2003). In general, rotifers are not as diverse or as abundant in marine environments as microcrustaceans, but they occur in estuarine waters, as well as in interstitial, tide pool, and near-shore marine habitats. Occasionally rotifers comprise an important portion of the biomass of marine zooplankton (Dolan and Gallegos, 1992). Inland saline waters, termed athalassohaline, are also habitats for rotifers (Segers, 2007; Walsh et al., 2008). Remarkably, some rotifers are found at the interface bridging aquatic and terrestrial habitats, i.e., they inhabit the film of water covering mosses, lichens, and liverworts. This habitat, referred to as limnoterrestrial (Figure 13.3(a)–(b)), is also home to nematodes (Chapter 14) and tardigrades (Chapter 17). Additionally, rotifers are members of pitcher plant and treehole communities, the phytotelmata (Figure 13.3(c)–(d)), and can be FIGURE 13.1 Lateral view of a generalized rotifer. Modified with permission from Koste and Shiel (1987). Chapter | 13 227 Phylum Rotifera found in containers holding water, such as birdbaths, as well as in discarded cups and tires stored outdoors (Figure 13.3(e)–(g)). Furthermore, rotifers often are abundant in the interstitial water of soils and sediments (Pourriot, 1979) including peat (Błędzki and Ellison, 2002). Depending on the soil type and its moisture level estimates of their densities range from about 32,000 to more than 2 million per square meter. Because of their high population levels and rapid metabolism, rotifers probably play an important role in nutrient cycling in soils. In addition to the variation in their habitats, the diversity of rotifer life histories is remarkable. Most are motile, either swimming as members of the plankton or crawling over plants or within sediments. However, after a brief, free-swimming stage, juveniles in three families of sessile rotifers attach permanently to a substrate, usually a freshwater plant (Wallace, 1980). The vast majority of rotifers are solitary, but about 25 species form colonies of various sizes (Wallace, 1987). Higher taxonomic groups are largely known for their differences in reproductive strategies. Two classes of rotifers are recognized: class Pararotatoria, comprising a single small family Seisonidae; and class Eurotatoria, containing subclasses Bdelloidea and Monogononta (Segers, 2002; Wallace et al., 2006). Seisonids are exclusively marine and obligatorily sexual. They are not discussed in detail here. All bdelloids are exclusively parthenogenetic, whereas monogononts are intermittently sexual; that is, they are cyclical parthenogens (see “Reproduction and Life History”). However, a complication to these generalizations is that males have never been reported for some monogononts. The variety of form (Figures 13.4 and 13.5) and life histories within the phylum offers a rich field of study. Additional accounts of this phylum may be found in most texts of general and invertebrate zoology, in some specialized books about inland waters (Wallace and Ricci, 2002; Wallace and Smith, 2009), or in advanced texts (Edmondson, 1959; Ruttner-Kolisko, 1974; Wallace et al., 2006). In the 1800s, some beautifully illustrated works were published that still offer an excellent depiction of these animals, although the taxonomy of some species is out of date (Hudson and Gosse, 1886). The digital images provided by Jersabek et al. (2003) of permanent slides made by Frank J. Myers are instructive for the diversity of animals covered, as well as for their historic value. There is no single scientific journal or set of journals in which researchers publish their research on rotifers; the field simply is too FIGURE 13.2 Female and male Brachionus species. Modified with permission from Pourriot and Francez (1986). SECTION | III 228 (a) (b) (c) (d) (e) (f) Protozoa to Tardigrada (g) FIGURE 13.3 Little-known habitats for rotifers. Two limnoterrestrial habitats: (a) Sphagnum moss in a bog pond; (b) lichen on a granite outcrop. Two phytotelma: (c) Sarracenia purpurea, the northern pitcher plant); (d) treeholes. Three container habitats: (e) birdbath; (f) discarded cup; (g) discarded tires. ((a)–(c), (e)–(g), R.L. Wallace; (d), courtesy of Christian Jersabek, University of Salzburg. diverse. However, since 1976, a small group of researchers (ca. 35–135) has gathered every 3 years to hold the International Rotifer Symposium. (At the time of this writing 13 such meetings have been held and their symposia volumes published.) Some of the papers discussed in this chapter were presented at the rotifer meetings. Several Internet sites describe rotifers and post stunning photomicrographs. However, we urge care in using keys posted on the Internet, as they are commonly based on regional samples, and identification of specimens based on photographs or line drawings alone is unwise. A detailed coverage of the phylum and of specific taxonomic groups is available in the Series Guides to the Identification of Microinvertebrates of the Continental Waters of the World. Biogeography Early studies of rotifer biogeography were dominated by the certainty that all species could be dispersed worldwide, an idea supported by the fact that the diapausing embryos of monogononts and the xerosomes and xeroova of bdelloids may be transported by insects, birds, and mammals (zoochory) and wind (anemochory). (See the later sections on “Reproduction and Life History” and “Environmental Physiology: anhydrobiosis”) Such passive dispersal, it was argued, is very effective at ensuring that most species become globally distributed. However, data have accumulated suggesting that this conclusion is mistaken. In fact, according to the last comprehensive survey of the literature (Segers, 2007), few species have a cosmopolitan distribution (Figure 13.6). Therefore, rotifers may not be as easily dispersed as previously thought. Moreover, once a dispersal stage arrives at a site, local abiotic conditions (e.g., pH, salinity, temperature) may be unsuitable for hatching. Additionally, successful colonization requires hatchlings compete for resources against competitors that have already become adapted to the idiosyncrasies of the habitat. This concept has been termed the Monopolization Hypothesis Chapter | 13 229 Phylum Rotifera FIGURE 13.4 Variation in morphology of bdelloid rotifers. Scale bars ca. 50 μm. Scanning electron photomicrographs courtesy of Diego Fontaneto and Giulio Melone, University of Milan, Italy. (a) (d) (b) (e) (De Meester et al., 2002). Thus, given the limits on dispersal and colonization, we should not be surprised that rotifers show biogeographical patterns. Recognizable trends in rotifer biogeography are evident. The fauna of Central America and the southern United States have close affinities to tropical South American fauna. Farther north, affinities with Europe are apparent, and endemism is strong in Keratella, Notholca, and Synchaeta. Yet, no single hypothesis best explains what we currently know about the biogeography of rotifers. Depending on the region, the rotifer fauna probably reflects the combined effects of recent glaciations and aridification, coupled with subsequent zoochory and anemochory. Nevertheless, there are some confounding issues to studying patterns of rotifer distribution. Biogeographical analyses are typically accomplished by collating information collected from published works. Thus, one obvious problem to achieving a comprehensive view of rotifer distribution is that rotifer researchers have not explored everywhere. For example, ∼52% of the species that have been recorded from only one realm have been reported in the Palearctic, and most of those from Europe. On the other hand, only ∼6% of single-realm records have been reported in the Afrotropical realm, which includes sub-Saharan Africa. These results are not surprising, given that Europe is the birthplace of rotifer research. Thus, the pattern of distribution illustrated in Figure 13.6 probably reflects where the collections were (c) (f) FIGURE 13.5 Variation in morphology of monogonont rotifers. (a) Asplanchna (foot absent), (b) Euchlanis (short foot with toes), (c) Epiphanes (prominent foot with toes), (d) Lecane (animal contracted into the lorica; short foot with prominent toes), (e) Testudinella (telescoping foot contracted into body), (f) Cephalodella (animal somewhat flattened laterally by the preparation; toe prominent). 230 FIGURE 13.6 Occurrences of rotifers among the eight biogeographical realms. (Data compiled from Segers (2007).) Most species have been found in a single biogeographic realm. Yet the question remains: Is this a matter of uneven distribution of species (i.e., a biogeography) or vastly uneven sampling or both? made more than the actual distribution of species (Fontaneto et al., 2012). A second problem is that some groups, such as bdelloids, are particularly understudied—some rightfully say nearly ignored. A separate issue is the phenomenon of seasonal polymorphism, which, in the past, confused workers who considered each morphotype to be a new species. A further question that complicates these studies is the dilemma of what actually constitutes a rotifer species; application of the biological species concept is problematic because males have never been described for many taxa. In practical terms, rotiferologists have been applying morphological criteria from the beginning of their discipline, but the morphospecies concept has its own issues. Chief among these is the fact that there are limited opportunities for workers to receive adequate training in rotifer identification. Moreover, several workers have shown that cryptic speciation is an important phenomenon in rotifers, including species in the genera Brachionus, Epiphanes, Keratella, Notholca, Philodina, and Synchaeta. Thus, minor variations in morphology, physiology, and/ or behavior that may be overlooked in identification may actually reflect differences important enough to warrant a new species designation. Efforts to clarify these matters will be greatly aided by modern genetic techniques, which include microsatellite analysis and sequencing molecular markers (e.g., CO1). Improved resolution of species identity is essential to a complete understanding of rotifer biogeography, a topic that Segers (2008) has explored in detail. Evolutionary Relationships The phylogenetic position of rotifers is still an open question. The current view suggests that rotifers are somehow related to the phylum Platyhelminthes, having been derived either from an ancestral flatworm or as a sister group to SECTION | III Protozoa to Tardigrada the flatworms. Another issue regarding the evolution of the Rotifera is the question of its relationship to the parasitic phylum Acanthocephala. The presence of the intracytoplasmic lamina (ICL) within a syncytial epidermis in both taxa, along with similarities in the ultrastructure of their spermatozoa, suggests that acanthocephalans and rotifers are closely related, even though acanthocephalans are 5–1500 times larger than the largest rotifer. Furthermore, detailed molecular analysis aligns acanthocephalans firmly with the rotifers, but exactly how they are related remains in doubt. Thus, some researchers argue that similarities in structure along with molecular evidence indicate that these two taxa are sufficiently related to be recognized as forming their own clade, the Syndermata (Sørensen and Giribet, 2006). Within this clade, several possible permutations of rotifer phylogeny have been postulated, including the following: acanthocephalans are: (1) highly modified bdelloids; (2) related to the group bdelloids + monogononts; (3) related to the group seisonids + bdelloids; or (4) a sister group to all rotifers. Nonetheless, for the present, taxonomists in both fields have retained the names Acanthocephala and Rotifera, ignoring the question of their phylogeny. Ultimately, integrative studies using total evidence (morphological and molecular data) should help to resolve issues of phylogenetic affinity. This goal is made more feasible perhaps by studies of the transcriptome of Brachionus plicatilis Müller, 1786 and discovery that the mitochondrial genome of this species is composed of two circular chromosomes of unequal copy number. Indeed, much remains to be accomplished, including a detailed analysis of a proposal to group a suite of small, jawed taxa—Gnathostomulida, Micrognathozoa, and Acanthocephala + Rotifera—into a superphylum Gnathifera (G., gnath, jaw and L., fera, bearing). Of course, to determine the phylogenetic relationships among these taxa, more molecular and morphological data, as well as a better sampling of taxa will be necessary. Evolution of bdelloids offers its own problems, but research has supplied some exciting new insights into these remarkable animals. Chief among the issues in bdelloid evolution are the following: How did asexuality arise in this group; how did their ability to become anhydrobiotic appear; and how are deleterious mutations eliminated from the population? Answers to these questions probably lie in the fact that bdelloids have undergone genome duplication, with the resulting gene copies evolving independent functions (i.e., they are degenerate tetraploids), as well as horizontal gene transfer, which has resulted in the acquisition of a diverse array of genes from viruses, bacteria, and other Metazoa (Ramulu et al., 2012). Current thinking on the evolution of the bdelloids and monogononts is that these two groups separated at least 100 million years ago. Chapter | 13 231 Phylum Rotifera GENERAL BIOLOGY External Morphology Rotifers exhibit a wide variety of morphologies; they are saccate to cylindrical in shape, sometimes appearing wormlike. Typically, the body is composed of three or four regions: head (corona), neck (in some forms), body (trunk), and foot, although the foot often is absent in planktonic forms. Although folds in the body may mark these regions, they do not represent segments; thus rotifers are not metameric (segmented). In many species, especially bdelloids, these creases function like joints or allow the body to collapse telescopically, thereby bending or shortening the animal, respectively (Figure 13.4). The generalizations noted here do not describe all rotifers well. In some forms, the neck and foot may be quite prominent, whereas in others they are absent. When present, the foot often extends from the body ventrally (Figure 13.1–13.3, 13.5). It usually possesses two toes, but this number may vary from 0 to 4. The foot also may possess pedal glands the ducts of which exit near the toes. These glands secrete an adhesive for temporary attachment to surfaces. On the other hand, juveniles of sessile rotifers release cement that forms a bond with the substrate that is not easily detached. If dislodged, the adults of sessile species do not reattach. In the subclass Monogononta, male rotifers (Figure 13.2) and juveniles of sessile rotifers (Figure 13.7) usually are much smaller and have a morphology different from that of adult females (Wallace, 1980; Wallace et al., 2006). In addition, male rotifers are often structurally simpler (see below: “Reproductive Systems”). Whenever males or juveniles are found in samples, their strikingly different morphologies may lead to improper identification. Rotifers possess a body wall (integument) containing a filament layer of varying thickness called the intracytoplasmic lamina (ICL) (Wallace et al., 2006; Wallace and Smith, 2009). This feature is shared only with the Acanthocephala, and thus is thought to be synapomorphic. The ICL of B. plicatilis, a halophilic rotifer, is composed of two filamentous, keratin-like proteins (Mr 39,000 and 47,000 Da) crosslinked by disulfide bonds (Wallace et al., 2006). Species in which major portions of the integument are strengthened by a thick ICL are termed loricate, and the thickened body wall is simply referred to as the lorica (L., armor). The integument of species without an extensive ICL remains thin and flexible. These rotifers are termed illoricate (L., il, without). In general, thickness of the body wall is of little taxonomic significance because extremes may be found within a single family or genus (e.g., Cephalodella). Even in loricate species, regions of the integument possess fewer ICL filaments, making the body wall flexible in that area. For example, flexibility is found in the coronal field, in the foot or at its junction with the body, and at articulations between movable spines and the trunk. In some rotifers, the surface of (b) (a) (c) (d) FIGURE 13.7 Adults and juveniles (larvae) of some sessile species. Family Flosculariidae: colony of adult Lacinularia flosculosa (a) and a juvenile somewhat compressed by the cover glass (b); Family Collothecidae: solitary adult (c) and a newly hatched juvenile. Bars ∼100 μm. (a) and (b), R.L. Wallace; (c) and (d), courtesy of Rick Hochberg, University of Massachusetts at Lowell. the body has various small projections (e.g., minute bumps or ridges). Many rotifers possess assorted fixed or movable spines, usually at the anterior and posterior ends. These serve as protection from predators. The morphology of rotifers is being better resolved by applying the techniques of electron microscopy and confocal laser scanning microscopy, coupled with immunohistochemistry (Hochberg, 2009). Some rotifers secrete materials from their body wall that congeals into a gelatinous mass in which various items become embedded, including inorganic silt, organic debris, bacteria, algae, fecal pellets, and pseudofecal pellets. These materials may form a tube that obstructs one’s view of the inside, but in other species the jelly remains transparent. In a few species, the secretions harden into a firm tube, as in the sessile monogonont genus Limnias, but in others the gelatinous material hardens into a series of thin, loosely fitted tiles or plates, as in SECTION | III 232 the bdelloid Mniobia incrassate (Murray, 1905). In some colonial species that produce tubes, the jelly mass coalesces into a continuous matrix that surrounds all members of the colony. On the other hand, the secretions of some bdelloids form small amorphous tubes, termed nests (Habrotrocha and Rotaria) and a few monogononts secrete only a thin jelly coat (Notommata copeus Ehrenberg, 1834). Many workers suggest that tubedwelling rotifers do so to reduce predation, but the tube may serve other functions. Ascomorpha eucaudis (Perty, 1850) and Cephalodella forficula (Ehrenberg, 1832), for example, feed on bacteria and/or algae that colonize the walls of their secretions, and the nests of bdelloids may serve as a way to retain water during periods of dryness. A few species do not produce tubes, but occupy cavities produced by other organisms: e.g., shells (tests) of ameba (Arcella and Difflugia), retort cells of Sphagnum moss, and the lobules (cup-shaped, ventral leaf) of the liverwort Frullania eboracensis. Organ System Structure and Function Internally, rotifers possess a perivisceral cavity commonly termed the pseudocoelom, which bathes muscles and nerves, and digestive, reproductive, and protonephridial organs (Figures 13.1–13.3). (Note: We use the term pseudocoelom for convenience, recognizing that some researchers recommend its elimination, as it is a polyphyletic condition.) Respiratory and circulatory systems are absent in rotifers. Although the characteristics noted above are unremarkable, rotifers do possess two curious features. First, the cells of all post-embryonic tissues are syncytial, i.e., multinucleate. Second, all individuals of a species have a consistent number of nuclei in each organ throughout life. For example, citing an older work, Libbie Hyman noted that there are about 900 nuclei per female in Epiphanes senta (Müller, 1773) and that the total number of cells in all species that had been studied to date was between 900 and 1000 (Hyman, 1951). Consistency in cell number, called eutely, is seen in a few other invertebrates (e.g., nematodes, Chapter 14). Nuclei may be seen using a standard light microscope, but special optics such as differential interference contrast (Nomarski) enhances their visibility. Research on rotifer anatomy has proceeded rapidly during the past 30 years and promises to continue to increase (e.g., Clément and Wurdak, 1991; Hochberg, 2006; Hochberg and Ablak Gurbuz, 2008; Hochberg, 2009; Smith et al., 2010). However, no single work provides a comprehensive examination of rotifer structure; here we provide an overview of the literature. Corona The classic description of the corona in rotifers is in the form of two ciliated rings called the trochus and cingulum (L., trocho, wheel and L., cingulum, belt). Nevertheless, Protozoa to Tardigrada there is considerable variation in the shape of the anterior end, and a dozen or so different coronal forms have been described based on overall shape, placement of the mouth, and distribution of cilia. The cilia often are responsible for production of water currents that are used in locomotion and feeding. In some forms, a prominent ciliated food groove lies between the trochus and cingulum; this is best seen in species of the order Flosculariacea. Yet, it is inadvisable to rely solely on coronal morphology in making identifications. For one thing, the head often retracts when the animal is disturbed, and in fixed specimens the shape of the corona can become distorted unless special methods of preservation are used. However, not all rotifers possess a ciliated corona. Adults of the families Atrochidae and Collothecidae exhibit the most extreme variation from the typical plan. In collothecids, ciliation may be sparse, and long setae surround the rim of a funnel-shaped structure known as the infundibulum (L., a funnel) (Figure 13.8). These setae prevent the escape of prey as the edges of the infundibulum fold over the victim, capturing it in a fashion similar to the Venus flytrap. Adult atrochids possess no setae on their corona (Figure 13.9). In this small group, prey are captured by a large infundibulum the edges of which draw inward, capturing the prey like a purse net. Collothecids also possess a vestibulum, a small chamber located beneath the infundibulum; here the prey is held before being processed by the mastax (Figure 13.8). In other rotifers, ciliation may be limited to a ventral field or to a few lobes, as is seen in some creeping monogononts and in some bdelloids (Melone and Ricci, 1995). Other structures that may be present on the corona include cirri, sensory antennae, and palps. Trophi and Gut Food is processed by the mastax (a modified pharynx), which is lined by a chitinous material developed as a set of translucent jaws or trophi. Trophi work the food in various ways (e.g., grasping, piercing, grinding, scraping) before passing the food through the esophagus to the stomach. In atrochid and collothecid rotifers, a portion of the mastax is enlarged into a food-storage organ termed the proventriculus. The mastax leads into the esophagus and in turn into the stomach. Most species possess an intestine and anus, but the gut ends in a blind stomach in some genera (e.g., Asplanchna, Asplanchnopus). The posterior portion of the intestine receives eggs from the oviduct and fluid from either a bladder or directly from paired protonephridia (Figures 13.1–13.2). This part of the gut is termed a cloaca. Often the gut is pigmented, depending on the nature of recently ingested material. Different species found in the same sample may possess guts that vary in color due to dietary preferences. Chapter | 13 233 Phylum Rotifera Rotifer trophi are composed of several hard parts and associated musculature that articulate in a specific spatial arrangement. In their basic form, trophi consist of three functional units: an incus (L., anvil) and paired mallei (L., hammers) (Figure 13.10). The incus is composed of three pieces: a fulcrum and a pair of rami (L., branches). The latter move like forceps and articulate with the fulcrum at their bases. Each malleus consists of two parts: manubrium and uncus. The manubrium (L., handle) resembles a clubshaped structure extended at one end into a cauda and flared at the other end (head). The manubrium articulates with a toothed structure called the uncus (L., hook). The plane of movement of the pieces comprising the malleus is at a right angle to that of the rami. In some species, the trophi may be modified by reduction of the basic parts, addition of accessory structures, or asymmetrical development of one or more of the pieces. Trophi are important features in rotifer taxonomy, as some classes, orders, families, and even species can be determined based on details of the trophi alone. Nine (a) (c) FIGURE 13.9 Cupelopagis vorax attached to the undersurface of an aquatic plant. Members of Family Atrochidae possess no setae on their corona. (b) (d) FIGURE 13.8 Representatives of the order Collothecacea. (a) Collotheca trilobata, a sessile species; (b) Collotheca libera with one embryo, a planktonic species; (c) Collotheca ferox, a sessile species; (d) Collotheca sp., with one diapausing embryo. SECTION | III 234 FIGURE 13.10 Malleate trophi of Cyrtonia tuba. In this scanning electron microscopy photomicrograph, all elements have been color-coded to the labels. Photomicrographs courtesy of Diego Fontaneto and Giulio Melone, University of Milan, Italy. (a) (b) (d) different types of trophi are recognized based on the size and shape of the seven pieces and presence of any accessory parts (Figure 13.11); transitional and aberrant types also are known. The “Rotifer trophi Web page” (http://users.unimi. it/melone/trophi/) provides an excellent overview of the different types of rotifer trophi. In Malleate trophi (Figure 13.11(a)–(c),(e)–(f),(p)), all parts of the incus and mallei are well developed and functional, but the rami are characteristically massive and may possess teeth along the inner margin. Furthermore, the unci have four to seven large teeth. This form works by grasping food and grinding it before pumping the crushed material into the esophagus. Malleate trophi are present in such common rotifers as Brachionus, Keratella, and Lecane. Malleoramate trophi (Figure 13.11(i)) are found only in the order Flosculariacea and resemble the malleate form, (k) (c) (m) (l) (f) (e) (o) (g) Protozoa to Tardigrada (n) (p) (h) (r) (q) (s) (i) (j) (t) (u) FIGURE 13.11 Rotifer trophi types. (a) Malleate trophi of Epiphanes, ventral; (b) malleate trophi (Epiphanes), ventral elevated; (c) malleate trophi (Epiphanes), lateral; (d) virgate trophi of Notommata, dorsal; (e) malleate trophi of Proales; (f) hypopharynx muscle in lateral aspect (Proales); (g) incudate trophi of Asplanchna; (h) uncinate trophi of Collotheca; (i) malleoramate of Ptygura; (j) forcipate trophi of Dicranophorus; (k) asymmetrical virgate trophi of Trichocera rattus; (l) virgate trophi of Eothinia; (m) virgate trophi of Itura, oral plates enlarged; (n) virgate trophi of Synchaeta with the powerful hypopharynx muscle; (o) virgate trophi of Ascomorpha; (p) malleate trophi of Proales gigantea, (somewhat intermediate between malleate and virgate types); (q) virgate trophi of Cephalodella, dorsal; (r) incudate trophi of Asplanchna; (s) virgate trophi of Cephalodella, lateral; (t) ramate trophi of the bdelloid Pleureta lineata; (u) Cardate trophi of the family Lindiidae. Bars = 20 μm. (a)–(s) with permission from the authors Koste and Shiel (1987) and CSIRO (http://www.publish.csiro.au/paper/IT9870949.htm); (u) with permission from the publisher Harring and Myers (1922). Chapter | 13 Phylum Rotifera except that, in the malleoramate form, the rami are strongly toothed, and the unci possess many thin teeth. Similar to the malleoramate form, ramate trophi (Figure 13.11(t)) have large, semicircular shaped rami and unci with many teeth. Ramate trophi lack a fulcrum and are limited to the subclass Bdelloidea. Only members of the order Collothecacea possess uncinate trophi (Figure 13.11(h)). Although they resemble the malleoramate, these trophi are characterized by unci possessing few teeth, usually with one large one and a few small ones. Virgate trophi (Figure 13.11(d), (k)–(o),(q),(s)) are modified for piercing and pumping, and they generally can be recognized by their long fulcrum and manubria and by the presence of powerful hypopharyngeal muscles. Some trophi of this form are asymmetrical. Virgate trophi are found in the common genera Notommata, Polyarthra, and Synchaeta. Forcipate trophi (Figure 13.11(j)), as the name implies, have an action like forceps whereby the trophi are projected from the mouth to grasp prey that are then swallowed. Forcipate trophi are limited to the family Dicranophoridae. Incudate trophi (Figure 13.11(g),(r)) are restricted to the family Asplanchnidae. They also function by grasping prey with a forceps-like action, but this form has a morphology different from that of the forcipate type; the rami are quite large and the mallei very small. The mastax actually initiates prey capture by creating suction, drawing the prey into the mouth. Once captured, the prey is stuffed into the stomach with the aid of the trophi. Lacking an intestine and anus, species of the genera Asplanchna and Asplanchnopus also use their trophi to extract indigestible materials from the stomach. Cardate trophi (Figure 13.11(u)) are found only in the family Lindiidae and function by producing a pumping action, without the hypopharyngeal muscle. The fulcrate type of trophi has been described as an aberrant form and is incompletely understood. This form is found only in the class Pararotatoria (see above), a very small group of marine rotifers comprising only four species within two genera: Paraseison (1 species) and Seison (3 species). A relatively unexplored facet of rotifer nutritional physiology is digestive enzyme function. Some work has been done with homogenates of whole animals (Kleinow and Röhrig, 1995), but histochemical and enzyme-labeled fluorescence techniques have permitted researchers to locate specific enzymes within organs (e.g., Strojsová and Vrba, 2007). 235 known as body-wall outgrowths (Asplanchna, Figure 13.12). In others, such as Brachionus calyciflorus Pallas, 1766 (Figure 13.13), increasing the pressure results in a flexing of the articulation between spines and the body in the posteriolateral region of the animal. This causes the spines to swing outward and stiffen. In this extended position, the spines interfere with predators (e.g., Asplanchna) that would otherwise consume undefended prey. Striated, longitudinal muscles are responsible for retracting the corona and foot and for moving certain articulating projections that are not moved by changes in the hydrostatic pressure of the body cavity. Hexarthra, for example (Figure 13.14), possesses powerful muscles that control movement of arm-like locomotory appendages. Contractions of these muscles cause a swift sweep of the arms that results in a rapid displacement or jump of the rotifer. Jumps in members of the genus Polyarthra are achieved by the sweeping movement of paddle-shaped appendages. However, in the genus Filinia, long setae (spines or bristles) are moved by the action of an increase in the hydrostatic pressure. FIGURE 13.12 Body form variability in the genus Asplanchna sieboldii. BWO = body wall outgrowths. Drawn from original photomicrographs courtesy of John J. Gilbert, Dartmouth College. Muscular System The muscular system consists of small groups of longitudinal and circular muscles inserted at various points on the integument or between the integument and viscera (Figure 13.2). In loricate species, the integument provides a firm structure against which the muscles work. In some species, muscles retract the corona, which increases pressure within the body cavity, thus expanding flaccid portions of the integument, FIGURE 13.13 Induction of spines in Brachionus calyciflorus. Left to right: spineless individual, three individuals with increasing spine development, individual with fully developed spines. Figure modified from Gilbert (1967), with the kind permission of the author and www.schweizerbart.de. 236 SECTION | III Protozoa to Tardigrada FIGURE 13.14 Hexarthra showing positioning of muscles that initiate jumps. Bar = 100 μm. Muscles also are present in the viscera, particularly in the mastax and stomach. Staining rotifer musculature with a fluorescent dye linked to phalloidan and examining specimens with a confocal laser-scanning microscope has revealed much finer details of these tissues (e.g., Hochberg and Ablak Gurbuz, 2008). FIGURE 13.15 Schematic representation of the nervous system of Plationus patulus revealed with immunohistochemistry (anti-serotonin, anti-FMRFamide, anti-neurofilament) and confocal microscopy. Note the presence of individual neurons in the coronal ciliated field. Abbreviations: br = brain; cnr = coronal nerve ring; ma = mastax; mg = mastax ganglion; pg = pedal ganglion; pn = pedal neuron; st = stomach and associated gastric neurons; tc = transverse commissure; vlc = ventrolateral nerve cord. Figure courtesy of Rick Hochberg, University of Massachusetts at Lowell. Neural System The nervous system is simple, comprising only a cerebral ganglion or brain (Figure 13.1) located dorsally on the mastax, a few other ganglia present in the mastax and foot, and three types of sensory organs: mechano-, chemo-, and photoreceptors (Clément and Wurdak, 1991). Mechanoreceptive bristles are situated on the corona, whereas several antennae are located elsewhere on the body, usually laterally and caudally (Figures 13.1–13.2). Chemoreceptive pores also are present on the corona. Many species possess one or more photoreceptive eyespots, sometimes containing a red pigment. When present, eyespots are located at the anterior end, usually near the brain. Although some rotifers retain their eyespots throughout life, sessile rotifers often lose them after the juveniles attach to a substrate. Paired, ventral nerve cords proceed from the brain along the length of the body into the foot. Several other ganglia are usually found in the nerve cords at the exit points for lateral nerves (Figure 13.15). One interesting structure found in the apical region of many bdelloid and monogonont rotifers is the retrocerebral organ (RCO). This structure consists of paired subcerebral glands and an unpaired retrocerebral sac, both with ducts that lead to the surface of the corona (Figure 13.1). Although the RCO function is not completely understood, fewer protonephridia occur in rotifers with well-developed RCOs (Edmondson, 1959). Some researchers have speculated that the RCO functions as an exocrine gland perhaps lubricating the rotifer’s apical end (Clément and Wurdak, 1991). Information on neurobiochemistry remains limited, but immunocytochemical studies have shown that acetylcholine functions as a neurotransmitter (a cholinergic system) in species from at least six families. In addition, norepinephrine neuroreceptor sites (an adrenergic system) have been reported in B. plicatilis, and widespread catecholaminergic neuronal systems have been observed in species of Asplanchna and Brachionus. Research also has identified serotonin and FMRFamide immunoreactive neurons in monogononts. Nicotinic receptors also have been identified in monogononts and at least one bdelloid. Neuron size and innervation varied among species, but neuron number was constant and species-specific. Excretory System: Protonephridium A paired protonephridial system composed of tubules and flame cells functions in excretion of nitrogenous wastes and osmoregulation in all rotifers (Figures 13.1–13.2). Usually there are only a small number of flame cells (fewer than Chapter | 13 237 Phylum Rotifera six), but large rotifers may possess many more. For example, Asplanchna sieboldii (Leydig, 1854) may have up to 100 flame cells (Ruttner-Kolisko, 1974). Normally, these tubules drain into a urinary bladder (Figure 13.2) that leads to a cloaca, but the bladder is absent in some species and a contractile cloaca assumes its function. Reproductive System Gonads are paired in both marine seisonids (class Pararotatoria) and the bdelloids (class Eurotatoria, subclass Bdelloidea); but in the latter, males are completely unknown and reproduction is by parthenogenesis. Monogonont rotifers (class Eurotatoria, subclass Monogononta) have only one gonad. Although males are present in this group, they have not been described for a large number of species. Even so, it is generally assumed that all monogononts can produce males given the proper conditions, or at least that the ancestral forms were capable of male production. However, when males are made the primary episode of sexual reproduction typically lasts for a few days to a week. Thus, unless collections are made frequently, male rotifers may never be observed. Nevertheless, even when a portion of a population is producing males, parthenogenetic reproduction usually continues in the remainder. Sexual reproduction (mixis) is initiated in many monogononts when females respond to a crowding signal via a labile, quorum-sensing molecule (mixis signal) released into the environment by the females themselves; this results in the production of males. Males may fertilize females, which in turn produce embryos that do not immediately hatch from their egg case; these are called diapausing embryos (see the later section on “Reproduction and Life History”). A few strains of the genus Brachionus have been reported to be obligate parthenogens, apparently having lost the capacity for sexual reproduction after long-term culture in a chemostat. The reproductive organs of females are composed of three units: ovary, vitellarium, and follicular layer. The ovary of a rotifer is a small, syncytial mass that is closely associated with the yolk-producing vitellarium. At birth, the adult complement of oocytes already has been formed in the ovary. The vitellarium also is syncytial with a specific number of nuclei—a characteristic useful in the taxonomy of some species. The follicular layer surrounds both the ovary and vitellarium. In some species, this layer forms the oviduct, which connects to the posterior portion of the gut at the cloaca (Figure 13.1). In general, monogonont males are smaller, swim faster, and have shorter life spans than females of the species, but this generalization does not hold for all species (e.g., Rhinoglena; Melone, 2001). In those species with small males (e.g., Asplanchna, Brachionus, Conochilus), the gut of the male is reduced or is absent entirely (Figure 13.16). In these (a) (b) (c) (d) (e) (f) FIGURE 13.16 Examples of male rotifers. (a) Anuraeopsis fissa (bar = 10 μm); (b) Asplanchna girodi (bar = 100 μm); (c) Brachionus calyciflorus (bar = 20 μm); (d) Brachious plicatilis (bar = 50 μm); (e) Lecane quadridentata (bar = 15 μm); (f) Platyias quadricornis (bar = 10 μm). (Photomicrographs (a)–(c) & (e), (f) courtesy of Roberto Rico-Martínez, Universidad Autónoma de Aguascalientes, Aguascalientes, México; d, Modified, with permission, from Pourriot and Francez (1986).) so-called dwarf males, the rudimentary gut serves as an energy source (Ricci and Melone, 1998), but there are other structural reductions as well (e.g., corona) (Gilbert and Williamson, 1983). The single testis is large and saccate, usually containing fewer than 50 freely floating mature sperm. A ciliated vas deferens leads from the testis to the penis, often with one or rarely two pairs of accessory (prostate) glands that discharge into it. In sessile species, the male is free swimming, whereas the adult female remains affixed to the substrate. Most rotifers are oviparous, releasing their eggs outside the body where the embryos develop. Many planktonic species carry their eggs attached to the female by a thin thread (Brachionus), fix them to a substrate (Euchlanis), or release them into the plankton (Notholca). The sessile species Sinantherina socialis (Linnaeus, 1758) carries its eggs on a specialized structure called the oviferon, which is located on 238 its elongated foot below the anus. However, a few species are ovoviviparous (e.g., Asplanchna and Cupelopagis), and thus retain their embryos in the body until the offspring hatches. Environmental Physiology Locomotion Most rotifers swim during at least a portion of their life cycle, but those lacking a foot (e.g., Asplanchna, Keratella, and Notholca) never attach to surfaces, even temporarily. Swimming can influence acquisition of food and mates, increase number of encounters with predators, and promote dispersal of the juveniles of sessile forms. Some rotifers are unusual in that they can swim, but routinely remain in mucus tubes attached to a substrate (e.g., certain Cephalodella species), while others are free-floating in mucus sheaths (Ascomorpha). In addition, some sessile species have relatives that have evolutionarily abandoned the sessile existence becoming mobile; these include Collotheca libera (Zacharis, 1894) (Figure 13.8(b)), Ptygura libera Myers, 1934, and Sinantherina semibullata (Thorpe, 1893). Rotifers often swim in a helical pattern, so that the actual distance traveled is greater than the linear displacement (Starkweather, 1987). However, for practical reasons, most researchers do not attempt to calculate absolute distance when considering the distance traveled, although automatic tracking systems have made this tedious work easier. While the theoretical power requirements of swimming (i.e., the energy required to overcome water resistance) is <1% of total metabolism, actual energetic cost appears to be much greater. In B. plicatilis, this cost has been calculated to be ∼38% of total metabolism (Epp and Lewis, 1984). Several researchers have measured the swimming speeds of male and female B. plicatilis. Swimming speeds also have been measured for other species such as Anuraeopsis fissa Gosse, 1851, Asplanchna brightwellii Gosse, 1850, B. calyciflorus, Euchlanis dilatata Ehrenberg, 1832, and Keratella americana Carlin, 1943. Collectively, these studies show that swimming speed is temperature dependent and varies with species and strain, age, number of eggs carried, food levels, and body mass. Values recorded for B. plicatilis at 25 °C range from 0.5 to 1.5 mm/s for females and 1.3–1.5 mm/s for males, with young and old females swimming about 30% slower than mature females. Asplanchna brightwellii and Keratella spp. females normally swim at 0.9 and 0.5 mm/s, respectively. Swimming speeds ranging from 0.17 to 0.54 mm/s were reported for 11 species of freshwater rotifers by Santos-Medrano et al. (2001). In this study Reynolds numbers varied between 0.023 and 0.301, with drag coefficients ranging from 9.7 × 10−7 to 1.6 × 10−5 N. We can infer that at these low Reynolds numbers the watery world is governed by laminar flow: i.e., at their size and swimming speed there is no turbulence. SECTION | III Protozoa to Tardigrada Little is known about the swimming speeds of the juveniles of most sessile rotifers, but swimming speeds for juveniles of Ptygura beauchampi Edmondson, 1940 are known to vary with age. For example, newborns up to mid-aged juveniles (∼3 h old) swam at about 2–2.5 mm/s. This speed is faster than the rates reported for B. plicatilis males. However, older juveniles (>4.5 h) swim at ∼1.0 mm/s. Concomitant with these changes in swimming speed is an increase in turning frequency. Correlations of rotifer ultrastructure with swimming speed, turning frequency, and other behaviors have been well studied and have demonstrated that rotifers are excellent models for comparative neurobehavioral studies. For example, swimming speed, mean turning angle, and angular speed varies in B. calyciflorus as a function of the condition of the medium and animal nutritive state. Nevertheless, much more work remains to be done before a complete synthesis of structure, function, and behavior will be possible. Spines and other appendages influence both swimming speed and sinking rates in rotifers. For example, unspined Keratella testudo (Ehrenberg, 1832) generally swim faster and sink more slowly than spined forms. On the other hand, Polyarthra normally swims at a much slower velocity (0.24 mm/s) than any species previously discussed, but it is capable of very short bursts (about 0.065 s long) of rapid movements called jumps (see above). During a jump, Polyarthra may attain velocities greater than 50 mm/s, with a mean velocity of 35 mm/s, or ∼100 times its normal swimming speed (Starkweather, 1987). Jumps are produced by movements of 12 appendages, called paddles, that articulate with the body near the head. When Polyarthra detects a disturbance in the water up to a few body lengths away, powerful striated muscles rapidly flex some of the paddles upward, initiating the jump. Then, during the jump, the paddles return to their original positions, and the rotifer is displaced an average of 1.25 mm (ca. 12 body lengths). Jumps help this rotifer escape invertebrate predators, such as the rotifer Asplanchna and first instars of the phantom midge Chaoborus, as well as the filtering currents of microcrustaceans such as Daphnia. They likewise are effective against naive workers attempting to remove Polyarthra from plankton samples using pipettes! Hexarthra also can jump rapidly, but movement in these forms has not been as well studied. In contrast, Keratella cannot jump to avoid the filtering currents of large daphnids; thus, the animals are severely damaged or killed when swept into the branchial chambers of cladocerans. However, Keratella is not without some escape abilities; it can increase its swimming velocity by a factor of about 3.5 when it encounters inhalant currents of Daphnia or comes into contact with the predatory rotifer Asplanchna. Physiological Ecology Physiological tolerances of organisms prescribe environments where survival and reproduction are possible. Thus, Chapter | 13 Phylum Rotifera an environmental tolerance curve for a species summarizes the range of environments where reproduction occurs and is bounded by the upper and lower lethal limits for the species. Within a tolerance curve the environmental optimum for a species is that environment where survival and reproduction are maximal. Therefore, a set of tolerance curves (including temperature, pH, salinity, oxygen concentration, etc.) indicates the breadth of adaptation of a species and its niche dimensions. Environmental tolerances and niche widths have been characterized for few rotifer species. For example, Epp and Lewis (1980) described the response of B. plicatilis to temperature. Their study determined respiration rate over temperatures ranging from 15 °C to 32 °C, and recorded Q10s of 1.9–2.4 for a broad temperature interval. Respiration levels off in the range of 20–28 °C, indicating that B. plicatilis can maintain a constant metabolic rate over this temperature range. At higher and lower temperatures, respiration rate increased, presumably because of thermal stress beyond the homeostatic capability of this species. Snell (1986) showed that amictic and mictic females have different temperature tolerance curves. Amictic B. plicatilis females reproduced at 20 °C and 40 °C, whereas mictic females did not. In general, amictic females reproduced over a broader environmental range of temperature, salinity, and food levels than did mictic females. Effects of pH on distribution and abundance of rotifers is a topic that has received a good deal of attention. However, because hydrogen ion concentration is related to other important chemical parameters in freshwaters, the effect of pH on rotifer occurrence may be indirect. Nevertheless, some extensive early work by F. Myers in the 1930s demonstrated that rotifers could be classified into a few broad groups based on pH alone: alkaline species, acid species, and those with a broad range. In the late 1980s, B. Bērziņš and B. Pejler, working from a large number of habitats in Sweden, concluded that species found in oligotrophic waters had pH optima at or below neutrality, whereas those species common to eutrophic waters had optima at or above neutrality. Moreover, they noted that acid-water species were often nonplanktonic or semiplanktonic. Such generalizations are, of course, never exact, and other factors are important. For example, Weithoff (2004) documented occurrence of two planktonic species—Cephalodella hoodii (Gosse, 1886) and Elosa worallii Lord, 1891—in an acid lake (pH 2.7) in Germany, where temperature and resource availability contributed to niche separation. Much less is known of the specific metabolic responses of rotifers to pH, but in B. plicatilis swimming activity and respiration rate does not significantly vary at pH values of 6.5–8.5 (Epp and Winston, 1978). However, swimming was reduced below pH 5.6 and above pH 9, with alkaline waters depressing swimming activity more than acidic conditions. 239 Regardless of the difficulties of interpreting the effects of pH on rotifers, we know from the work of Frost et al. (2006) that a shift in pH can radically alter zooplankton community structure. For example, increasing acidification of lakes as a result of acid precipitation has been widespread in North America, and this has led to changes in the composition of the zooplankton, e.g., increasing dominance of Keratella taurocephala Myers, 1938. This observation has been repeated in lakes that have been experimentally acidified. In general, it appears that rotifers become more important as members of the zooplankton with increasing acidity. Individual rotifers rarely experience large fluctuations in osmotic conditions over their relatively short lifetime. However, a population living in a small water body subject to periodic inundation followed by evaporation may need to cope with increasingly greater solute concentrations. These habitats include tide pools, marine coastal ponds, and desert rock pools. Brachionus plicatilis, a common euryhaline species, survives osmolarities over the range of two to >95 psu (i.e., ∼50 to >2750 mOsmol/l). Transferring of B. plicatilis directly from ∼1.5 to ∼33 psu causes considerable mortality, but acclimation to elevated osmolarities can be achieved by gradually increasing ionic concentrations. On the other hand, B. plicatilis does not tolerate low osmolarities well; this probably accounts for its restriction to alkaline and brackish waters. This well-studied species can probably tolerate these extreme salinities because it can regulate its internal bodily fluid via a membrane-bound, Na+/K+ ATPase pump (Lowe et al., 2005). Although most rotifers require oxygen concentrations significantly above 1.0 mg/l, some tolerate anaerobic or near anaerobic conditions for short periods. Other species routinely live in oxygen-poor regions, such as the hypolimnion of eutrophic lakes or in sewage ponds. Rotifer resistance to starvation has been investigated in a number of species. For example, in the bdelloid Macrotrachela quadricornifera Milne, 1886 survivors of a long starvation period (20–60 days) resumed reproduction after being fed (Ricci and Perletti, 2006). Life history parameters like mean lifespan and lifetime fecundity were not different in starved bdelloids after the resumption of feeding, suggesting that starved rotifers simply shift their schedule of survival and fecundity. Thus, in bdelloids, metabolic reduction during starvation is similar to their physiological response to drought. Using life table experiments, Weithoff (2007) investigated the response of two monogononts, Cephalodella sp. and E. worallii, to starvation. Using food concentrations below the threshold for population growth, he found a tradeoff only in E. worallii, which increased lifespan at the expense of reproduction. Differential resistance to starvation is important, as it can determine the outcome of competition and thus influence community structure. For example, research has shown that starvation resistance was higher for Keratella cochlearis SECTION | III 240 (Gosse, 1851) grown on N-limited Cryptomonas rather than nutrient-sufficient algae. Food quality also has been shown to explain the vertical distribution of Cephalodella sp. and E. worallii in a meromictic lake in Germany (Weithoff and Wacker, 2007). Laboratory research confirmed that fecundity, growth, and resistance to starvation by E. worallii were negatively affected when the rotifer was fed the alga Chlamydomonas acidophilia that had been grown under heterotrophic vs. autotrophic culture conditions. However, Cephalodella sp. grew on a diet of C. acidophilia regardless of conditions under which the algae had been grown. In the field, a population of E. worallii was abundant in the epilimnion (above 6 m), where individuals of C. acidophilia would have been growing autotrophically. On the other hand, individuals of Cephalodella sp. were more abundant in the hypolimnion (below 6 m) where C. acidophilia would have been growing heterotrophically. Environmental Toxicology Because rotifers fill an important ecological role and are relatively easy to culture, they have been important in assessing the toxicity of chemicals in the laboratory, as well as examining the effects of pollutants in natural habitats. The response of rotifers to a variety of toxicants has been characterized in both natural and laboratory populations. Most methods have used brachionid rotifers, but other species have also been used. These involve both short-term (acute) and long-term (chronic) toxicity tests. The test materials include crude oil and other petrochemicals, oil dispersants, detergents, free ammonia, wastewater, endocrine disruptors, anti-infective agents, cyanotoxins, insecticides, herbicides, and heavy metals. Research has shown how exposure to these factors modifies rotifer life table parameters such as age-specific survivorship and fecundity, and the production of diapausing embryos. Standardization of toxicity tests provides a way to compare the effects of toxicants, which may otherwise differ depending on the protocol. A standardized acute toxicity test has been described that employs freshwater (B. calyciflorus) and marine (B. plicatilis) test animals (ASTM, 2012). This procedure is less expensive than many others because it uses test animals hatched from diapausing embryos, so it is not necessary to maintain stock cultures. Moreover, the test is simple, rapid, and sensitive. In addition, a standardized chronic toxicity test based on asexual reproduction has been published in Standard Methods for the Examination of Water and Wastewater (Snell, 1998). Because toxic agents affect swimming and feeding behaviors, these phenomena also have been used to assess toxicity. Besides such direct effects, toxicants can modify rotifer community structure through indirect effects, for example by altering food composition and thereby changing the outcome of resource competition. Protozoa to Tardigrada In general, rotifers seem to serve as good indicators of water quality in natural environments. In Europe, for example, composition of the rotifer community has been advocated as a component in the Saprobic Index. (The Saprobic Index is a measure of saprobity, an estimate of the level of organic pollution as measured by a combination of the biological oxidation demand of a water sample and the presence of specific indicator organisms in the habitat.) Studies of natural and experimental contamination of water bodies have shown that rotifer species composition and abundance change depending on the toxic agent and its concentration. In a study of the effects of artificial acidification on a lake in north central Wisconsin (USA), Frost et al. (2006) showed that whereas the zooplankton community (including rotifers) improved after release from the acid stress, the trajectory of the recovery followed a different path from the decline, thus indicating a substantial hysteresis. Even a temporary contaminant can have a strong effect on rotifers. After a spill of the so-called Red Sludge in the Danube, the rotifer community disappeared. Although it recovered a few weeks later, the levels were lower than before the spill (Schöll and Szövényi, 2011). Sustained, heavy contamination can have a profound effect on zooplankton composition. In a reservoir in the Chelyabinsk region of Russia, which appears to be the most radioactively contaminated inland waters in the world, the plankton community is composed of a near monoculture of cyanobacteria and rotifers (Pryakhin et al., 2012). Anhydrobiosis The ability of rotifers to tolerate desiccation and then be revived later has been known since the early 1700s, when Leeuwenhoek described rehydration of rotifers present in sediments collected from dry rain gutters. This phenomenon, called anhydrobiosis, is known to occur in both adults and embryos of many bdelloids. However, in monogononts, dormancy occurs only in the diapausing embryos, which also are capable of anhydrobiosis. These embryos are the product of sexual reproduction (see the section “Reproduction and Life History: Diapausing Embryos in Sediments”). While in the desiccated state, bdelloids and diapausing embryos are capable of passive dispersal by air currents (anemochory) or by mobile animals (zoochory), either attached on the outside (ectozoochory) or in their digestive tracks (endozoochory). An anhydrobiotic adult bdelloid rotifer resembles a wrinkled barrel and has been called a tun (Middle English, meaning barrel); unfortunately, this descriptive term is also routinely applied to anhydrobiotic tardigrades (Chapter 17). To avoid confusion, the term xerosome (G., xero, dry and G., soma, body) has been proposed (Wallace and Smith, 2009) for anhydrobiotic adult bdelloids. In xerosomes, the head and foot retracts into the animal’s trunk; at this Chapter | 13 Phylum Rotifera time, the female may deposit a mature egg (Figure 13.17). An egg, really an embryo, deposited during the desiccation process also can withstand desiccation and is termed a xerooum (G., dry egg). While the desiccation process is complex, the adaptive potential is substantial, as many bdelloids inhabit environments that dry completely at irregular intervals (Ricci et al., 2007). A few strictly aquatic bdelloids cannot withstand desiccation. Bdelloid anhydrobiosis involves more than simple drying; unless loss of metabolic water proceeds slowly, the rotifer usually dies. During anhydrobiosis, the fine structure of cells is retained, but in a greatly modified state. Changes that occur internally include at least a 50% reduction in the volume of the body cavity, a condensation of cells and organs, and a decrease in cytoplasmic volume, so that the entire animal is only about 25–30% of its original size. Nuclei, mitochondria, endoplasmic reticula, and other organelles form a compact mass within cells of the xerosomes (Ricci et al., 2007). Xerosomes have been reported to be viable even after more than two decades in the anhydrobiotic state. Recovery from anhydrobiosis may require as little as 10 minutes, or it make take several hours, according to prevailing environmental conditions. Survival is influenced by both biotic and abiotic factors. For example, young and starved animals are less likely to survive anhydrobiosis. Moist environments and high temperatures during desiccation also reduce recovery. A noteworthy feature of anhydrobiosis in bdelloids is that the time spent as a xerosome does not affect the total life span; this phenomenon has been termed the Sleeping Beauty Model (Ricci and Caprioli, 2005). For example, Ricci and colleagues have shown that 8-day-old Adineta ricciae Segers and Shiel, 2005 desiccated for 7 days lived as long as a group from the same cohort of animals that was not FIGURE 13.17 Adult bdelloid xerosome (left) and its desiccated embryo (xerooum) (right). Scanning electron photomicrograph courtesy of Giulio Melone, University of Milan, Italy. 241 desiccated (mean (±SD) = 41.2 (1.2) and 37.7 (1.2), respectively). However, the desiccated group did produce significantly more offspring per animal over their lifetime (mean (±SD) = 37.6 (0.4) and 31.9 (0.4)), respectively. Given that rotifers are eutelic, this increase in fecundity offers a mystery, especially as Ricci and colleagues noted that this species possesses only about 30 nuclei in its germarium. Ricci and coworkers suggested that anhydrobiosis may stimulate oocyte production by the germarium but then remind us that this idea runs counter to the current understanding of eutely. Further complicating our understanding of anhydrobiosis in bdelloids is the fact that, in at least two bdelloid species (A. ricciae and M. quadricornifera), repeated desiccations and rehydrations appear to enhance fitness over those in continuous culture. Anhydrobiosis in bdelloids occurs without production of the nonreducing disaccharide trehalose or an analogous molecule, and no trehalose synthetase genes have been identified (Mark Welch et al., 2009). Nevertheless, hydrophilic proteins found in desiccation-tolerant plants become abundant late in the embryo of the common bdelloid Philodina roseola Ehrenberg, 1832. Although trehalose is absent from bdelloids, it is present in desiccated diapausing embryos of the monogonont B. plicatilis. The molecular mechanisms of anhydrobiosis need further investigation, and bdelloids seem to be an excellent research model for dormancy studies. One intriguing proposition is that extra copies of the gene for alpha tubulin may be related to their ability to withstand desiccation. This is suggested because 11 to 13 copies have been found in bdelloids (Eyres et al., 2012) when only four to eight would be expected in these degenerate tetraploids. Bdelloids also possess extreme resistance to 137Cs gamma ionization radiation (IR) (Mark Welch et al., 2009). For example the bdelloids Adineta vaga (Davis, 1873) and P. roseola reproduce (albeit at a reduced rate over un-irradiated controls) even after exposure to doses of more than 800 Gy (by comparison, a chest X-ray will deliver ∼10−4 Gy). In contrast, the monogonont E. dilatata experienced greater reduction of fecundity, but at a radiation dose of only 20% that of the bdelloids. Moreover, although the bdelloids experienced hundreds of doublestranded breaks (DSB) to their DNA during exposure to the radiation, the damaged was repaired. Thus, bdelloids are not resistant to IR but possess an extraordinary ability to repair DSB. Their ability to tolerate desiccation and ionization radiation may stem from the fact that bdelloids are degenerate tetraploids (see above), with collinear pairs of chromosomes that may serve as templates for DSB repair via a form of homologous recombination (Mark Welch et al., 2009). Moreover, processes associated with DSB repair may foster removal of deleterious elements and have contributed to the persistence of these ancient asexual animals. 242 Generalized Stress Responses Studies have shown that rotifers respond to stresses such as heat, UV radiation, and low pH by producing an array of heat shock proteins (HSP) (e.g., Smith et al., 2012). In addition, Late Embryo Abundant genes (LEA) help in maintaining homeo-osmotic conditions (Denekamp et al., 2010). However, recent work indicates that rotifers also produce Transient Stress Granules (SG) in response to stress, but the extent to which they are produced varies by the nature and duration of the stress (Jones et al., 2013). Additional research will, no doubt, elucidate the relationship among environmental stressors and these stress response mechanisms. ECOLOGY AND EVOLUTION Diversity and Distribution Phenotypic Variation Phenotypic variation is an important adaptive mechanism in rotifers, but has posed difficult problems for systematists. Intraspecific variation arises by several mechanisms including cyclomorphosis, dietary- and predator-induced polymorphisms, dwarfisms, and polymorphisms in hatchlings from diapausing embryos. Cyclomorphosis is the seasonal phenotypic change in body size, spine length, pigmentation, and/or ornamentation found in successive generations of zooplankton. These changes are morphological alterations in the individuals of a single population that are related to physical, chemical, or biological features of the environment. Each different morphological form is called a morphotype (or morph). Specifically excluded from cyclomorphic changes are seasonal succession of sibling species and clonal replacements of genotypes (both of which are genetic changes in populations or communities), as well as induction of spines or changes in shape due to the presence of predators or specific biochemicals in the diet. Gilbert (1980) described a striking phenotypic change in morphology associated with a dietary polymorphism for three Asplanchna species (A. brightwellii, A. intermedia Hudson, 1886, A. sieboldii). Diets that include the plant product α-tocopherol (vitamin E) induce saccate females, the smallest morphotype, to produce cruciform daughters. Cruciforms have lateral outgrowths of the body wall (Figure 13.12) that protect them from cannibalism by conspecifics by making them larger and thus more difficult to ingest if captured. In the presence of vitamin E and certain prey types, cruciforms can produce a third morphotype called campanulates (more prevalent in A. sieboldii and A. intermedia). Campanulates are very large females (>2000 μm) that heavily cannibalize saccate females. Female polymorphism is much less pronounced in A. brightwellii; their increase in body size is only 50–60% larger than the norm. Dietary SECTION | III Protozoa to Tardigrada polymorphism in Asplanchna (gigantism) may have evolved originally as a generalized growth response to larger prey typical of eutrophic waters. The tocopherol response probably is adaptive, because it may signal availability of nutritious rotifer and microcrustacean prey. Another important source of phenotypic variation is predator-induced polymorphism. This topic was briefly considered above (“Environmental Physiology: Locomotion”). Spined and unspined forms had been recognized in several rotifer species for many years, but the causes and consequences of these variations remained an enigma (Figure 13.13). Induction of spines was first demonstrated in B. calyciflorus: the offspring of adults exposed to culture medium that had previously held predatory Asplanchna developed spines that dramatically reduced their vulnerability to predation by Asplanchna. Additional research has shown that several other prey species are capable of spine induction by Asplanchna, as well as by other predators. Two additional kinds of variation are aptera generations in Polyarthra (i.e., polymorphisms in the hatchlings of diapausing embryos) and a body size that is smaller than is commonly seem. The latter has been called dwarfism, but this term should not be confused with the smaller size often seen in monogonont males. Aptera morphotypes were initially thought to be different species of Polyarthra, but later were shown to be forms lacking the paddles that are characteristic of this genus. Only the generation hatching from the egg bank of diapausing embryos lacks paddles; their parthenogenetic offspring develop into typical morphotypes with these appendages. Similar polymorphisms between diapausing embryos hatchlings and parthenogenetic generations were described for Keratella quadrata (Müller, 1786) and are suspected for Notholca acuminata (Ehrenberg, 1832). Dwarfism in Brachionus caudatus Barrois and Daday, 1894, as reported for crater lakes in the Cameroon, is characterized by reduced body size and spination compared to normal morphotypes; high temperature, combined with reduced food supply and a different suite of predators, may account for this condition. However, we cannot discount the possibility that these populations actually represent cryptic species. Another source of phenotypic variation in body size is polyploidy. For example, Walsh and Zhang (1992) reported substantial variation in body size for different populations of E. dilatata. During a 12-month period of intensive sampling in Devils Lake (Oregon), these workers observed two distinct body sizes: a smaller morphotype ∼225 μm in length and a larger one ∼275 μm. The larger morphotype had a chromosome number of 21 and the smaller morphotype had 14 chromosomes. The haploid chromosome number found in males of this species is n = 7. This was the first case of polyploidy reported for rotifers, but new techniques for staining and counting rotifer chromosomes may make more systematic explorations possible. Chapter | 13 243 Phylum Rotifera Beyond morphological variation, plasticity also occurs in sexual reproduction. Transgenerational phenotypic plasticity in the responsiveness to the mixis signal has been described in B. calyciflorus. In some strains of this species, females hatching from diapausing embryos do not react to this biochemical, but, as the population develops, females become increasingly more responsive to it. That is, sexual reproduction is initiated, and a portion of a population becomes sexual. Low mixis propensity continues for up to 12 generations after hatching. This suppressed responsiveness to mixis signals also has been observed in Brachionus angularis Gosse, 1851, Rhinoglena frontalis Ehrenberg, 1853, and E. senta. Delayed mixis is regarded as an adaptive response to promote rapid asexual population growth soon after hatching, followed by sexual reproduction at high population densities (Serra et al., 2005). However, populations of Hexarthra sp. from temporary pools in the Chihuahuan Desert have a different mixis response (Schröder et al., 2007). Some of the diapausing embryos that hatch in these populations immediately go sexual, resulting in production of a new batch of diapausing embryos. Meanwhile, the remaining newly hatched embryos reproduce asexually. Because these small habitats may dry in a matter of days or persist for weeks, this variation is seen as a bet-hedging strategy: some diapausing embryos are produced immediately, while a portion of the population continues to expand asexually. Distribution and Population Movements Water bodies are not uniform habitats with respect to biotic factors such as food and predators and abiotic factors such as dissolved oxygen concentration, light intensity, temperature, and water movements. Therefore, it is not surprising to find that rotifers are not evenly distributed in lakes and ponds; often there is considerable variability with respect to their horizontal and vertical distribution. For example, two Filinia species displayed very different vertical distribution patterns over the course of a year in a small lake in Germany (Figure 13.18). In some meromictic lakes with strong clinograde oxygen gradients, population maxima may be restricted to near the oxycline. Similar patterns are seen in the horizontal distribution of rotifers. In fact, some rotifers are strictly littoral, being found in open waters only as occasional migrants. Others are pelagic, but the depth at which they are found varies with season. Although these major distribution patterns result from differential population growth and water currents and other large-scale water movements, within-lake distribution patterns can be influenced to a lesser degree by the locomotory behaviors of the animals. One commonly recognized behavior of marine and freshwater zooplankton is a daily (diel) vertical migration (DVM) in the water column in which the animals usually come to the surface only during the night. During the day zooplankton avoid visual predators (fish) FIGURE 13.18 Temporal and spatial distribution of two species of Filinia in Lake Pluβee, Germany. Solid line indicates limits of the population at one individual per liter; dotted line indicated boundaries of higher population levels (numbers of individuals per liter). Modified with permission from Hofmann (1982). 244 that occupy near-surface waters, but when they return to the surface at night they can exploit the rich algal resources present there. In rotifers, diel migrations typically are not as dramatic as those of microcrustaceans. Thus, the population center of rotifers usually changes only about 1–3 m over a daily cycle. For example, a study on the zooplankton of Ross Creek Reservoir (New Zealand) showed that Conochilopsis causeyae (Vidrine, McLaughlin, and Willis, 1985) exhibited a nocturnal ascent with its population center ascending from 2.1 m in the daytime to 0.9 m at night. However, two other species did not show a nocturnal rise: Hexarthra mira (no change) and K. cochlearis (reverse migration). In addition, ovigerous (egg-bearing) and nonovigerous females may have different DVM patterns. Ignoring this fact can cause serious errors in the calculations of birth rates for field populations. Errors of nearly an order of magnitude may occur if the population is sampled at only one depth or at different times during the day. Rotifers also move horizontally in aquatic systems, with some pelagic rotifers avoiding water close to the shore. This phenomenon, called avoidance of the shore, was demonstrated by using a circular plexiglass arena that permitted the researchers to assess the swimming direction of several zooplankton species over a set time interval. When the shadow produced by the natural elevation of the shoreline was artificially altered by adding a black collar around the arena, two pelagic species, Asplanchna priodonta Gosse, 1850 and Synchaeta pectinata Ehrenberg, 1832, swam away from the shadow, whereas the littoral rotifer, E. dilatata, showed no preference in its movements. Besides the effect of the shadow cast by the shoreline, rotifer position may be explained by differences in composition of the hydrophyte community in the littoral zone. For example, both lily pads and extensive populations of duckweed (Lemna) provide shade under which large numbers of Conochilus hippocrepis (Schrank, 1803) may congregate (Edmondson, 1959). However, plant physiological activities also alter the chemical properties of the water in the immediate vicinity of the plant. For instance, polar plants change the concentration of Ca+2 within the Prandtl boundary (see below), and Chara produces a musky smell that permeates the water. Rotifers also are present in the psammon, the sandy habitat along the wet reaches of shorelines. While population levels are never great in this interstitial (L., between) realm, their diversity can be significant. For example, EsjmontKarabin (2003) found a total of 110 species in the psammon of beaches of 18 lakes in Poland, a value representing 26% of all species recorded from Polish waters. This community included 22 species that occurred exclusively in the psammon, with monogononts apparently playing a much more important role than bdelloids. Her study also reported a tendency for higher rotifer diversity in the psammon communities of mesotrophic and eutrophic lakes as compared to hypertrophic lakes. SECTION | III Protozoa to Tardigrada Research on the abundance and distribution of rotifers in lotic systems has lagged behind that of lentic ones. However, we do know that both rotifer diversity and abundance are usually lower in rivers (∼5–400 individuals/l) than in lakes, although in some instances the population density may be quite high (ca. >6000/l). Some of the animals in flowing waters seem to be derived from upstream lakes or secondary channels. However, rotifers can reproduce in both the main and secondary channels and are, for all practical purposes, the dominant zooplankton (not counting protists) present in certain lotic systems, such as rivers of the Great Plains (Thorp and Mantovani, 2005). Sampling strategies for rivers can be as complicated as those carried out in lakes. These techniques include bank-to-bank net tows, repeated collections at a single site, or Lagrangian sampling. In Lagrangian sampling, the same parcel of water is repeatedly sampled as it flows downstream. Colonial Rotifers Most rotifers are solitary and interact only as potential prey or mates, but a number of species in 10 genera form intraspecific colonies or join in the formation of interspecific colonies (Figure 13.19). Nearly all of the colonial species are members of two monogonont families (Conochilidae and Flosculariidae), although colony formation has been noted in at least one bdelloid, but those colonies appear to be temporary (Wallace, 1987). All colonial species are microphagous and many produce secretions of various sorts that link colony mates. For example, some produce tubes from secretions that harden (Limnias), whereas in others the secretions are gelatinous (Conochilus and Lacinularia). A few species construct a tube out of pseudofecal pellets (e.g., Floscularia conifera (Hudson, 1886)) or jelly and fecal pellets (Ptygura pilula (Linnaeus, 1758)), with the resulting structures resembling the turret of a castle. These secretions are important in the overall structure of the colony, as they provide a substrate for attachment of larvae or a matrix in which new members are added to the colony (see below). Because colonial rotifers do not reproduce by budding or the formation of specialized zooids, colony members are not intimately connected, as are colonial bryozoans (Chapter 16); therefore, energy resources are not shared among colony members. The number of individuals comprising a colony varies greatly among genera. Floscularia ringens (Linnaeus, 1758) usually builds colonies of fewer than five individuals, as do some members of the family Conochilidae (e.g., Conochilus dossuarius Hudson, 1885). On the other hand, some species construct colonies of intermediate size (up to ∼35 individuals; Conochilus unicornis Rousselet, 1892), whereas large colonies (50–200 individuals) are produced by F. conifera, S. socialis (Linnaeus, 1758), and C. hippocrepis. A few taxa have been reported to produce colonies of enormous size: e.g., >500; Lacinularia elliptica Shephard, 1897. Chapter | 13 Phylum Rotifera FIGURE 13.19 Examples of colonial rotifers. (a) A portion of a small sessile colony of Limnias melicerta. These animals live within a clear, firm tube that they secrete. This species normally is solitary or forms colonies of only a few individuals. (b) A sessile colony of Lacinularia flosculosa. This species secretes a gelatinous matrix. (c) A tiny colony of Conochilus (=Conochiloides) dossuarius comprising a female and her most recent offspring. As in all of the Conochilidae, this species secretes a gelatinous matrix that is often populated by algae, bacteria, and sometimes Vorticella. Bars = 250 μm. So far, the mechanisms that control colony size have not yet been explored. For example, C. unicornis usually forms small colonies of fewer than 35 individuals; but in four lakes with the cladoceran predator Leptodora, we 245 have collected much larger colonies from a single sample. In those cases, the ranges of mean colony size varied from about 82 to >200 individuals. However, our record to date is from a population in Green Lake (Wisconsin, USA), where the largest colonies comprised more than 400 individuals. Similar observations of >50 individuals per colony have been made of C. unicornis populations in Canadian Shield lakes that have the cladoceran predator Bythotrephes longimanus. The mechanism that initiates this exuberant colony form has not yet been elucidated, but it seems likely that a predatory induction is responsible, as in predatorinduced spine formation in B. calyciflorus. In this case, induction may involve an alteration in the cohesiveness of the gelatinous matrix of the small vs. large colony forms. Colonies form by one of three methods (Figure 13.20), each of which produces colonies that presumably differ in the genetic relatedness among colony mates (Table 13.1). In allorecruitive colony formation, free-swimming juveniles produce colonies by settling on tubes of sessile adults. Thus, colonies of allorecruitive species grow in size (i.e., numbers of individuals per colony) by intercolonial recruitment of juveniles. The number of colonies within the habitat (colony density) increases by juveniles settling as solitary individuals, which are then joined by new recruits. Because juveniles joining these colonies may come from females belonging to another colony, genotypic relatedness within the colony is probably relatively low. These colonies are transitory, beginning when a recruit attaches to a previously settled adult and ending when recruitment to an old colony ceases. Late recruits to allorecruitive colonies may suffer because death of the founding individual can lead to dislodgment from the substrate and subsequent sinking to the benthos. Some species of the genera Floscularia and Limnias reproduce this way. A few species produce interspecific colonies of two or more species, but fusion of established colonies does not seem possible (Wallace, 1987). In autorecruitive colony formation, the young remain with their mother in the colony. Thus, colonies of autorecruitive species grow in size (numbers of individuals per colony) by intracolonial recruitment of the young into the parental colony, whereas the number of colonies within the habitat (colony density) increases by colony fission. Because the young joining autorecruitive colonies come from the parent colony, genotypic relatedness within the colony is probably high. Autorecruitive colonies are long-lived and develop continuously throughout the season, increasing in size as new individuals are added and decreasing only when the colony divides. Late recruits to these colonies probably do not suffer because, although older individuals die, the colony continues. Autorecruitive species have not been reported to produce interspecific colonies, nor has colony fusion been described. Species in the family Conochilidae, as well as S. semibullata, reproduce by autorecruitive colony formation. SECTION | III 246 Protozoa to Tardigrada of two different sizes (ages), which is likely the result of colony fusion (i.e., a juvenile colony joins with a previously settled colony). Lacinularia flosculosa (Müller, 1773) and S. socialis reproduce by geminative colony formation. The consequences of high genetic relatedness have not been explored, but may include increased vulnerability to parasites, uniformity of behaviors, and decreased genetic diversity of diapausing embryos. Two hypotheses have been offered to explain the adaptive significance of coloniality in rotifers. One hypothesis suggests that colonial animals possess an energetic advantage over solitary individuals of the same species. For example, colonial F. conifera apparently live longer and mature faster than solitary individuals. It is argued that juxtaposition of filtering currents produced by two or more individuals permits an increased filtering rate and/ or an enhanced filtering efficiency. Although experiments have not supported the idea that coloniality affects filtration rates, the dynamics of colony feeding currents suggests that coloniality increases filtering efficiency (Wallace, 1987). A second hypothesis argues that colonial existence can protect individuals from certain predators. For example, large C. hippocrepis colonies embedded in their gelatinous sheath are less vulnerable to attack by a calanoid copepod predator because they are too large to be engulfed whole. Also, in C. unicornis individual rotifers retract into the refuge of the gelatinous matrix of the colony and cannot be captured by the predatory rotifer Asplanchna. Sessile Rotifers FIGURE 13.20 Three types of colony formation in rotifers: Allorecruitive (Floscularia conifera), Autorecruitive (Conochilus), and Geminative (Sinantherina socialis). In geminative colony formation, all of the young born within a span of a few hours leave the parent colony as a free-swimming, juvenile colony. Members of this young colony subsequently explore and attach to a new substrate together. Thus, genotypic relatedness within a colony is probably moderate to high, depending on its particular history. Because size of a juvenile colony is dependent on fecundity of its parental colony, the number of individuals comprising a colony does not generally increase over its lifetime. Geminative species have not been reported to produce interspecific colonies, but it is possible that they do. We also have seen colonies that are composed of individuals Sessile species are found in three families: Atrochidae (two genera), Collothecidae (two genera), and Flosculariidae (nine genera). Although they are often overlooked because their microhabitats are generally not examined thoroughly, these forms are actually quite common in lakes and ponds. Occasionally they reach very high densities on plant surfaces (>6 individuals/mm2), especially in bogs and small eutrophic ponds. The juvenile motile stages of sessile rotifers (Figure 13.7) are not true larvae, as all of the adult organs are present in the young animal. However, because of the conceptual parallel with sessile marine invertebrates, and because the corona does not appear to be completely developed in the juvenile, the term larva is often used. Not surprisingly, the behavior of juveniles changes dramatically once they come into contact with a potential attachment site. These new behaviors have been described using terms such as selection, choice, and preference. However, use of such words is not meant to imply cognition by the rotifer (that would be a form of teleological thinking). These are merely convenient terms to describe this phenomenon. Several workers have demonstrated that juveniles can select a particular substrate among all surfaces available Chapter | 13 247 Phylum Rotifera TABLE 13.1 Colony Formation in Rotifers Colony Formation Category Increase in Colony Size (Individuals per Colony) Mode of Increase of Colony Density in the Habitat Predicted Genotypic Relatedness Within Colonies Frequency of Interspecific Colony Formation Allorecruitive (intercolonial) Young are recruited to established colonies or remain solitary Young leave the parental colony and may establish a new colony Low Common Floscularia (Figure 13.20); Limnias (Figure 13.19) Autorecruitive (intracolonial) Young stay within the parental colony Fission: adult colonies High fragment, producing smaller colonies Absent Conochilus (Figure 13.20); Sinantherina semibullata Geminative Rare to absent Cohorts of larvae collectively establish new colonies Rare to absent Lacinularia (Figure 13.19); Sinantherina socialis (Figure 13.20) Moderate to high for settlement. For example, juveniles of F. conifera settle with a greater frequency on the tubes of conspecifics than on aquatic plants, although there is substantially more plant surface available. This propensity leads to the formation of intraspecific colonies, each with 50 or more individuals. During the growing season, ≥75% of the entire population may be colonial. Some species attach to a surface based on water chemistry. In a population of Collotheca (gracilipes) campanulata (Dobie, 1849) in a small pond filled with several hydrophyte species, including Elodea, Lemna, Myriophyllum, and Nymphaea, most individuals were present on Elodea canadensis (Wallace and Edmondson, 1986). In experiments in which juvenile C. campanulata were presented with submerged Elodea, >90% of the individuals preferred the under surface of the leaves to the upper surface (Figure 13.21). (In this case, the under surface is anatomical; it does not merely refer to the way the plant was growing in the water, which may be sideways or upside down due to crowding by other plants.) Attachment to the under surface is apparently in response to the way in which Elodea is able to alter the concentration of calcium ions in the water immediately around the leaf, i.e., in the Prandtl boundary layer. In water having a pH in the neutral to alkaline range, Elodea acts as a polar plant, removing Ca+2 from beneath the leaf and releasing it above. Choice of the under surface of the leaves of Elodea provides a superior habitat in comparison to the upper surface. In short-term laboratory experiments, young C. campanulata attached to under surfaces of Elodea leaves grew significantly taller and produced more eggs per female than those induced to attach to the upper surfaces of the same leaves. Several other species of sessile rotifers exhibit strong preferences for particular substrates, but the significance of these associations has not been fully elucidated. Examples FIGURE 13.21 Members of the sessile species Collotheca campanulata (=gracilipes) colonizing the undersurface of a leaf of the aquatic macrophyte Elodea canadensis. Many (>25) individuals may be seen attached to the under surfaces of the leaf. A biofilm may be seen on the upper surface of the distal one third of the leaf (leaf length ≈1 cm). Substrate selection behaviors have been described for three species: C. campanulata, P. beauchampi, and S. socialis. In general, juveniles appear to react to potential surfaces in similar ways. Newborns avoid settling for periods up to several hours after hatching. Once this refractory period is past, all surfaces are explored; however, some receive much more attention and generate different behaviors, including some reminiscent of male mating behavior. A juvenile will traverse these surfaces with both its corona and foot in contact with the surface, occasionally stopping in this slightly bent position. The juvenile may continue exploration of the surface for several minutes, but eventually it attaches to the surface using a type of cement from glands in the foot and then undergoes metamorphosis. 248 Immediately after attachment and metamorphosis, the young of most sessile rotifers begin to secrete a protective tube. Often this secretion is in the form of a clear, gelatinous material (e.g., Collotheca and Stephanoceros; Figure 13.8), but in some species the tube becomes obscured by colonizing microorganisms and debris (e.g., Ptygura and Beauchampia; Figure 13.22 a). At least two sessile species augment their jelly tube with fecal pellets (Floscularia janus (Hudson, 1881) and P. pilula; Figure 13.22(b)). Limnias melicerta Weisse, 1849 forms a cement tube that looks like a series of transparent rings placed on top of one another (Figure 13.22(c)). Regardless of how the tube is formed, it probably deters some predators, but fragments of the tubes of certain sessile rotifers may be found in snail fecal material (e.g., Limnias). Perhaps the most fascinating example of tube construction is found in the genus Floscularia (Figure 13.20). Species in this genus possess a small ciliated cup (modulus) on the ventral side of the head. In some species of this genus, the animals pass tiny particles and other small debris collected by the corona. The cilia in the modulus appear to be in constant motion, mixing gelatinous secretions with the particles to form small pellets (pseudofeces), either in the shape of bullets or balls. Once a pellet is fully formed, the rotifer places it on the top of the tube in an action resembling the movements of a bricklayer. In this way, the tube is constantly elongated as the animal grows. Because pellets are manufactured from particles collected from the water, they are colored, usually light to dark brown. However, when a heavy rainstorm temporarily suspends soil in the water, the pellets produced by Floscularia usually turn out to be very dark (Figure 13.23). Thus, the event of the storm is marked as a dark ring in the tubes of all the animals alive at that time. In his classic study, Edmondson (1945) noted this fact, and used suspensions of powdered carmine and SECTION | III Protozoa to Tardigrada carbon black to mark the tubes of F. conifera so that he could study the dynamics of population growth in a field population. Another unexplored aspect of sessile rotifers is the presence of anisotropic (birefringent) crystalline structures (ACS) that have been found in the guts of several species (Wallace et al., 2006). Anisotropic structures possess two to three refractive indices. Such structures may be visualized by examining a specimen set between cross polarizing filters. Under this illumination, the background is black and the ACS bright. They also are visible under standard bright field illumination as dark structures. When present, ACS (ca. 10–30 μm) may take the form of small balls of compact crystals, as in the free-swimming juvenile stages of S. socialis, as well as in the embryos of other sessile taxa. FIGURE 13.23 Dark pellets in the tube of Floscularia conifera. In this instance, the animal alternately used darkly colored and lightly colored suspended material to create its pellets. Seen here are five (left to right) successive occurrences of dark bands. (See also Figure 13.20, top panel, where the banding pattern is also visible.) FIGURE 13.22 Some variation in some tube construction by rotifers. (a) Beauchampia crucigera possesses a gelatinous tube obscured by debris. (Closed arrow = tube; open arrow = single antenna.) (b) Ptygura pilula possesses a clear gelatinous tube supplemented with fecal pellets (fp). (c) Limnias melicerta produces a clear tube resembling a stack of rings. s = subitaneous egg. Bars = 100 μm. Chapter | 13 Phylum Rotifera As juvenile S. socialis age, these structures break apart into small individual crystals that slowly disappear. However, in the adults of the raptorial predators, Acyclus inquietus Leidy, 1882 and Cupelopagis vorax ACS resemble compact spheres (Figure 13.24). Although present in the free- swimming juveniles of A. inquietus and C. vorax, these bodies are still present in adults and enlarge as the animal ages. In neonates, ACS may represent a source of energy that helps the juvenile through settling and metamorphosis; their persistence in adults may represent energy reserves that are used during periods of starvation. Reproduction and Life History Reproduction The type of reproduction varies considerably within Rotifera. Species in class Pararotatoria (seisonids) reproduce exclusively through bisexual means, gametogenesis occurring via classical meiosis with the production of two polar bodies. At the other extreme, members of subclass Bdelloidea reproduce entirely by asexual parthenogenesis (i.e., via apomictic thelytoky). Thus, no males have been (a) (c) (b) 249 observed in bdelloids, a fact that has been referred to as an “evolutionary scandal” (Mark Welch et al., 2009). On the other hand, species in subclass Monogononta exhibit cyclical parthenogenesis where asexual reproduction predominates, but sexual reproduction occurs occasionally. However, loss of capacity for sex has been documented within laboratory populations, and males have not been documented for all species. Cyclical Parthenogenesis and Diapause Cyclical parthenogenesis in monogononts (Figure 13.25) involves an asexual (amictic) phase and a sexual (mictic) phase. Although at certain times both asexual and sexual reproduction occurs concurrently, in most populations only amictic females are present. Amictic females are diploid and produce diploid eggs termed amictic or subitaneous eggs. These embryos develop mitotically via a single equational division and usually hatch as females within 24 hours. Although the term summer egg has been applied to these embryos, the term is misleading because, depending on the species, amictic rotifers can found at any time of the year. The switch in the reproductive mode of amictic to mictic is initiated when an amictic female responds to a specific stimulus (e.g., a conspecific chemical signal or an environmental cue) and begins to produce mictic daughters. These (d) FIGURE 13.24 Examples of anisotropic crystalline structures (ACS) in sessile rotifers. (a) juvenile Sinantherina socialis, with ACS dispersing from its compact mass (cross polarized filters slightly offset); (b) adult Acyclus inquietus, ACS ∼30 μm (bright field); (c) recently settled young Cupelopagis vorax, ACS group ∼18 μm (bright field); (d) amictic embryo of Floscularia conifer, ∼15 μm (cross polarized filters). Symbols: arrows = ACS; int = intestine; pro = proventriculus; sto = stomach. FIGURE 13.25 Generalized life cycle for monogonont rotifers. The life cycle of bdelloids consists of only the parthenogenetic portion. Mixis stimulus: absent (−); present (+). Timing of mating for newborn mictic females is critical for the outcome: early = diapausing embryos (2n), late = no fertilization, male embryos (n). SECTION | III 250 mictic females then produce haploid eggs via meiosis that, if unfertilized, develop into haploid males (n) and, if fertilized, develop into diapausing embryos (2n). So far, the genetic controls of the parthenogenetic switch have not been well examined; however, recently Hanson et al. (2013) have shown that a cell cycle regulatory gene is important to maintaining the typical cyclical parthenogenetic cycle. If a male fertilizes a mictic haploid egg still early in its development, the diploid condition is restored. However, development of the fertilized mictic egg (zygote) is arrested well before maturation; this results in a diapausing embryo. These embryos possess thick walls that often are sculptured (Figure 13.26). (In some literature, the terms resting egg, winter egg, or cyst are used to describe the diapausing embryo, but we suggest that these terms be abandoned. Mixis is not confined to the colder months, and the term cyst has other meanings in biology. In addition, once cell cleavage begins, a fertilized egg is properly termed an embryo.) Diapausing embryos are very resistant to harsh environmental conditions, including desiccation, and may be dispersed by wind (anemochory), migrating animals (zoochory), or flowing water (hydrochory). After a period of dormancy (a) (b) (c) (d) (e) (f) FIGURE 13.26 Diapausing eggs of several monogonont rotifers. (a) Asplanchna girodi; (b) Hexarthra fennica; (c) Brachionus calyciflorus; (d) Conochiloides natans; (e) Sinantherina socialis, (f) Kellicottia bostoniensis. (Bars = 10 μm; the wrinkled backgrounds in panels (b) and (f) are membrane filters that held the embryos during preparation.) Original scanning electron photomicrographs courtesy of Hendrik Segers, Royal Belgium Institute of Natural Sciences, Belgium. Protozoa to Tardigrada that varies among species, diapausing embryos respond to environmental cues and hatch as diploid amictic females (Gilbert and Schröder, 2004). The stimuli that induce hatching may include changes in light, temperature, salinity, and oxygen concentration. As noted above, variations exist to this typical sexual cycle. Diapausing embryos of Hexarthra sp. from rock pools in the Chihuahuan Desert can hatch into mictic females, bypassing canonical mixis induction pathways (Schröder et al., 2007). This may be adaptive for a population that lives in ephemeral habitats that may dry up within a few days or weeks after a rainfall, facilitating rapid production of sexually produced diapausing embryos able to withstand desiccation. With a few notable exceptions, the stimulus for initiating sexual reproduction is poorly understood. Vitamin E controls the shift from amictic to mictic reproduction in some Asplanchna species, and photoperiod plays a similar regulatory role in Notommata. A chemical signal produced by the rotifers themselves triggers mixis in at least four species of Brachionus, two species of Epiphanes, and R. frontalis. This process is analogous to quorum sensing (QS) in bacteria (Snell, 2011). In B. plicatilis, the mictic signal has been proposed to be a protein that induces sex upon accumulating to a threshold concentration as population density increases. As a result, rotifers are capable of auto-conditioning their medium via secretion of the mixis induction protein(s) (MIP). In some instances, auto-conditioned medium is effective in inducing mixis at population densities below about one individual per 15 ml. In fact, females can be so sensitive to their MIP that individuals are sometimes cultured separately with bi-daily transfer to avoid accumulation of the signal to threshold concentrations. Due to nonlinear effects, culture volume can affect the observed density for mixis induction in laboratory experiments. In addition to environmental factors, genetic factors play a major role in determining the sensitivity of particular strains to mictic stimuli. Strains that have lost the ability to undergo mixis may still release the MIP but have lost the ability to respond to it. At this point, the exact nature of the MIP has not been determined. Amictic females produce oocytes one at a time during their reproductive period, but the embryo’s fate—i.e., to become an amictic female or to develop into a mictic female—depends on the concentration of the MIP to which the mother is exposed (Gilbert, 2007). Oocytes exposed to high levels of MIP prior to, or soon after, oviposition have a greater chance of developing into a mictic female than those exposed to lower concentrations. However, the probability of amictic vs mictic progeny is not uniform over the life of the maternal female. Offspring produced early or late in a female’s reproductive sequence have a lower probability of developing as mictic females. Chapter | 13 Phylum Rotifera Not all reproduction in monogonont rotifers follows the pattern described above. In some populations of Asplanchna, Conochilus, and Sinantherina, amphoteric females (individuals that produce both female and male offspring) have been recorded (Wallace et al., 2006). Amphoteric rotifers can produce both diploid (female) and haploid (male) eggs, with some producing both females and diapausing embryos, or males and diapausing embryos. The presence of amphoteric reproduction in other rotifers has not been fully investigated, and its significance in the life history of those genera that exhibit it remains to be determined. A second variation is production of eggs that resemble diapausing embryos (e.g., present of a multilayered shell) but that were produced via parthenogenesis in the absence of males. These eggs are termed pseudosexual eggs. In S. pectinata, two different types of parthenogenetic amictic eggs are produced. One type resembles normal subitaneous eggs in that they are thin shelled and develop and hatch without diapause within 1 day. Production of the second egg type is induced by starvation. These eggs are thick-shelled and enter an obligatory diapause after one to three cleavage divisions. Duration of diapause in these eggs ranges from 4 to 13 days. The adaptive significance of diapausing amictic eggs seems to be to increase the ability of populations to survive short-term food limitation. Synchaeta pectinata also produce diapausing embryos following sexual reproduction, and these have the capacity for extended dormancy. Diapausing Embryos in Sediments Because of their capacity for extended dormancy, diapausing embryos theoretically can accumulate to high levels in sediments, but the densities reported vary considerably. On an aerial basis, densities of diapausing embryos extracted from sediments range from 100 to 4000/cm2. Apparently, embryos can be abundant and still viable deep into the sediments. In one study, diapausing embryos collected from at least 21 cm deep (>40 years old) were still capable of hatching. However, reports of viability in much older sediments have been published (Wallace et al., 2006). Of course, the age of diapausing embryos in sediments cannot directly be determined, but their age can be estimated from information about the sediment itself (e.g., using radiometric dating techniques that use 210Pb and 137Cs). Sediments from dry vernal pools in both temperate and desert waters also will yield rotifers when rehydrated under the proper conditions (Langley et al., 2001). Schröder (2005) described conditions required for diapausing egg formation, survival, and hatching, as well as possible strategies involved in these processes. No doubt the number of diapausing embryos in basin sediments is a function of both abiotic and biotic processes. Abiotic factors include sedimentation rate, sediment focusing (concentration of sediments in certain regions of the basin), and sediment mixing and burial. Biotic factors include previous production levels, 251 bioturbation, hatching rates, and mortality in the sediments, including from predation and disease. Embryogenesis The study of embryogenesis has not kept pace with other aspects of rotifer reproduction. In their examination of the bdelloid M. quadricornifera, Boschetti et al. (2005) reported embryological studies on seven species prior to 1925 and only five more between 1956 and 1988. Most of those studies used light microscopy on live embryos. However, confocal laser scanning microscopy has shown that early development of M. quadricornifera is similar to that of other rotifers: holoblastic cleavage leads to gastrulation that occurs by epiboly. Using phallodin conjugated with rodamin, these workers visualized the mastax and trophi, noting that these structures were formed within a mold of actin filaments. In another study using confocal laser scanning microscopy, Boschetti et al. (2011) identified the development stage at which the diapausing embryos of nine monogonont species became arrested. In one group, embryos stopped cell division after about five cell divisions (mean ∼30 nuclei, with low variation in nucleus number); in the second group cell division stopped after another division, but the variation in nucleus number was greater (∼40–60). Reproductive Behavior Descriptions of mating behaviors are available for many species, including A. brightwellii, B. calyciflorus, B. plicatilis, E. senta, Lecane quadridentata (Ehrenberg, 1830), Platyias quadricornis (Ehrenberg, 1832), and Trichocerca pusilla (Jennings, 1903). The males and females in these as in other species show pronounced sexual dimorphism, with males being smaller and faster swimmers (Figure 13.2). Lacking a functional foot, males swim constantly without attaching. Because males and females swim randomly, the probability of male–female encounters in planktonic species can be modeled mathematically (Snell and Garman, 1986). In the colonial, sessile rotifer S. socialis, males can copulate with several females of one colony in succession. Females take no active role in locating a mate, but B. plicatilis females often exhibit specific reactions such as foot flipping or accelerated swimming once a male encounter occurs. Males, at least for some species, display a distinct mating behavior upon encountering conspecific females. In Brachionus, mating behavior begins when the corona of the male squarely contacts the female. However, not all head-on encounters result in mating; indeed, probability of copulation in laboratory cultures generally varies from 10% to 75% in B. plicatilis, depending on the strain. The requirement for head-on contact by the male is thought to be due to the presence of chemoreceptors in his coronal region that apparently respond to a species-specific glycoprotein on the surface of the female. Mating begins with the male swimming circles around the female, skimming over the surface 252 FIGURE 13.27 Male mating behavior in brachionid rotifers. Arrows indicate general swimming movements by male and female rotifers. (Male not drawn to scale.) of her lorica (Figure 13.27). During this phase, the male maintains contact with the female with both his corona and penis; this requires the male to remain in a slightly bent position. After several seconds of circling, the male attaches his penis to the female, usually in the region of her corona, and loses coronal contact. After about 1.2 minutes of copulation in B. plicatilis, sperm transfer is completed and copulation is terminated when the male and female break apart and swim away. Newborn B. plicatilis males only have about 30 sperm, and they transfer two to three at each insemination. The littoral rotifer E. senta displays an unusual mating behavior that is unique for monogonont rotifers (Schröder, 2003). Females are mostly stationary on the substrate, whereas males are active swimmers. If a male encounters a conspecific female embryo, he remains near it, apparently waiting to mate with the newly hatched female. Males discriminate between male and female eggs, and diapausing embryos, and they exhibit a strong preference for female embryos that are only a few hours from hatching, probably by sensing a chemical diffusing through the eggshell. However, males have shown no difference in attending eggs that would develop as mictic vs amictic females. Only in the former case would a mating result in a fertilization and production of diapausing embryos. This mating behavior is similar to the precopulatory mate guarding of copepods, but it lacks male monopolization of the female. Aging Rotifers have been used as models of aging and senescence for a long time. Early studies on the monogonont Proales sordida Gosse, 1886 suggested the potential for maternal effects, in which increased age of the maternal female was associated with a reduced overall lifespan. Concomitant with decreased lifespan was greater variability in fecundity SECTION | III Protozoa to Tardigrada and rate of development, in her progeny. This theme was explored in a series of papers in the 1940s, which showed that parental age also influenced longevity in the bdelloid Philodina citrina. In these studies, orthoclones were established by isolating the first offspring of parental females, then the first offspring of F1 females, and so forth. This procedure yields an isogenic clone derived exclusively of firstborn females. Orthoclones comprising the offspring of older females also were generated (e.g., a series of the sixth offspring). This protocol is a powerful tool for investigating effects on aging, because the only difference among orthoclones is the age of their mothers. In P. citrina, the mean lifespan decreased in older orthoclones, suggesting that the phenomenon was controlled by a maternal effect that was transmissible and cumulative. However, this effect was reversible, because first-born females from an old orthoclone outlived their parents. Based on these and other studies, it was proposed that accumulation of calcium was the maternal factor responsible. This phenomenon became known as the Lansing Effect after the researcher who first reported it. The rate of calcium accumulation and its importance in rotifer senescence became the reigning hypothesis of the day. Since this early work, researchers have reported similar results in some species but not in others. However, a re-examination of the original data indicates that there may not have been a direct effect on aging but, rather, that short-lived offspring from older mothers had increased levels of reproduction and began to reproduce at a younger age (King, 1983). Conversely, the first reproduction of long-lived females born to young parents did not occur until a later age. King argued that the Lansing Effect actually could be attributed to alterations in fecundity, whereby reproduction becomes concentrated within a few age classes in lines of short-lived rotifers. Concentrating reproduction into such an abbreviated time scale may itself be the cause of the shortened lifespan. Until more is known about the mechanism for these changes, King suggested that interpretations of the Lansing Effect be made with caution. Other experiments on aging indicate that vitamin E and thiazolidine-4-carboxylic acid (TCA), or a combination of chemicals can extend lifespan in both bdelloids and monogononts. Presumably, these chemicals work by quenching free radical reactions. Two other factors that influence free radical production are light and diet. Lifespan is influenced by photoperiod (light:dark cycle (L:D)), with longer photoperiods decreasing lifespan (e.g., short 0:24 vs long 12:12). Additionally, laboratory experiments have shown that ultraviolet radiation (UVR) shortens lifespan significantly, with lifespan declining logarithmically as a function of dose. UVR has the potential for damaging zooplankton in natural systems, but for rotifers susceptibility is species-specific and also depends on water temperature and concentration of dissolved organic carbon Chapter | 13 Phylum Rotifera (DOC) in the lake. For example, some species are protected from UVR by the presence of biochemicals called mycosporine-like amino acids (MAAs) that absorb critical wavelengths. However, MAAs are not present in all rotifers; those that possess them do so by accumulating these biochemicals from their diet. A separate physiological response to UVR damage is via photoenzymatic repair (PER). Water conditions may influence light penetration into lakes and thus the impact of UVR on zooplankton. Lakes with significant levels of DOC, and presumably other lightabsorbing materials, attenuate UVR at shallow depths when compared to clear water. Therefore, very low levels of DOC in the water column have the potential to structure zooplankton species composition, favoring those species with MAAs or the ability to undergo repair processes such as PER, or that undergo DVM. For example, Obertegger et al. (2008) linked the vertical distribution of rotifers in the water column to dry weight levels of MAAs. In their study of a clear-water oligotrophic lake Italy, they found that P. dolichoptera and Synchaeta grandis Zacharias, 1893, which remained in the upper layer during the daytime, had ∼6 and 2 μg MAAs mg-1 dry weight, respectively. Dietary restriction has life-extending effects in a wide variety of animals, including rotifers. Two different modes of dietary restriction can yield longer lifespans: reduction of food intake and intermittent feeding. One study of dietary restriction in rotifers indicated that restriction caused an increase in mean and maximum lifespan, reproductive lifespan, and doubling time in most species. It seems plausible that chronic dietary restriction is associated with changes in reproductive allocation during starvation. Most species that reduced their reproduction during starvation experienced increased lifespan. In contrast, species that continued reproduction through starvation experienced decreased lifespan. Yet, exceptions exist with some species not showing a clear tradeoff in lifespan and fecundity. Male lifespan has been examined far less thoroughly than that of females, but in those species in which the male does not feed (e.g., Brachionus manjavacas Fontaneto, Giordani, Melone and Serra, 2007) males live only about half as long as females (Figure 13.28). To our knowledge, no information is available on the lifespans of males in species that do feed (e.g., R. frontalis). Population Dynamics Studies of rotifer population dynamics attempt to explain the causes of changes in population size and community structure. By separating and quantifying the relative contributions of reproduction, mortality, and dispersal, ecologists are able to recognize factors regulating population size and determinants of average abundance. Because some species are easily cultured and experimentally manipulated, rotifers are useful models for investigating the dynamics of animal 253 FIGURE 13.28 Female and male survival of Brachionus manjavacas. populations. Techniques also exist for investigating the dynamics of natural rotifer populations, further broadening their usefulness in ecological studies. Life Tables The most rigorous approach to studying population dynamics is life table analysis, i.e., age-specific records of all reproduction and mortality occurring in a cohort. The general protocol for gathering life table data on rotifers is to collect a group of newborn females, isolate them in small volumes of culture medium (∼1–2 ml), and make daily observations of their survival and reproduction. Maternal females usually are transferred daily to fresh medium with a specific food level, and offspring are counted and removed. A table is then constructed of age, number surviving in each age class, and number of offspring produced in each age class. From these observations, vital population statistics can be calculated such as mean and total lifespan, age at first reproduction, age-specific survival (lx) and fecundity (mx), net reproductive rate (Ro), intrinsic rate of increase (r), generation time (T), and stable age distribution. The detailed observations required for life table analysis usually are possible only in laboratory populations. A number of genera have been examined in this way, including species of Asplanchna, Brachionus, and several bdelloids. In certain cases, life table analyses can be performed on field populations, as in Edmondson’s classic study of the colonial sessile rotifer F. conifera (Edmondson, 1945). The major environmental factors affecting age-specific survival and reproduction are temperature, food quantity and quality, genetic strain, and reproductive type (either amictic or fertilized or unfertilized mictic females). Increased temperature shortens lifespan. For example, in a study of B. calyciflorus by Halbach (1970), 50% of a cohort survived to age 16 days at 15 °C, 11 days at 20 °C, and 5 days at 25 °C (Figure 13.29). Fecundity is compressed into fewer age classes at higher temperatures. At 15 °C, fecundity occurs at a low rate over 20 days. In 254 SECTION | III Protozoa to Tardigrada FIGURE 13.29 Effect of temperature on age-specific survival and fecundity of Brachionus calyciflorus. The left Y-axis is the percentage of the cohort surviving (open circles), and the right Y-axis is the offspring per female per hour (closed circles). Figure redrawn from Halbach (1973). FIGURE 13.30 Rotifer population size is determined by a variety of factors, including food quantity and quality and levels of predation and parasitism. contrast, at 25 °C reproduction occurs at a high rate over 10 days. Females produce a total of 13 offspring at 15 °C, 16.6 at 20 °C, and 12.9 at 25 °C. The intrinsic rates of increase (r) were 0.34, 0.48, and 0.82 offspring per female per day at the respective temperatures. Food supply obviously is an important determinant of rotifer abundance and many studies have shown that both the quantity and quality of the diet have an impact on population growth. However, interspecific competition (with rotifers, cladocerans, and protists) and predation (e.g., from cladoceran and copepods) are also important. For example, the large predatory cladoceran Bythotrephes longimanus, a recent invader into North American lakes, appears to have had a significant negative impact on herbivorous and predatory cladocerans and predatory copepods in Harp Lake (Canada). As a result, the population of the colonial rotifer C. unicornis increased significantly when released from the pressures imposed by these crustaceans (Hovius et al., 2007). Thus, to obtain a realistic view of rotifer growth potential, both bottom-up and top-down processes need to be considered (Figure 13.30). Lake area (Ao) and food supply, as measured by annual primary productivity (gC/m2/yr), also are important factors in determining rotifer species richness (S). Dodson et al. (2000) reported that there was a significant positive linear relationship between log S and log Ao for 33 well-studied lakes located in the northern hemisphere. They also present a regression analysis of their dataset fitted to a quadratic model, arguing that there is a unimodal relationship between S and productivity, with the highest S occurring at ca. 100 gC/m2/yr. Rotifers also can detect and rapidly exploit microscale patches (thin layers, ≤0.5 m) of food resources. In a laboratory study, individuals of B. plicatilis quickly aggregated within thin layers of food, either Nannochloropsis oculata or Skeletonema costatum or both, until the resource was depleted (Ignoffo et al., 2005). From this work, we may infer that free-swimming rotifers utilize their habitat according to Ideal Free Distribution theory, but this is a topic that needs greater study. The effect of food quantity on lx and mx schedules can be substantial. In one intensive study, Halbach and Halbach-Keup (1974) fed B. calyciflorus the green alga Chlorella at densities from 0.05 to 5.0 × 106 cells/ml at 20 °C. In that study the best survival occurred at algal concentrations 0.5 × 106 cells/ml and 1.0 × 106 cells/ml, where mean lifespan was 9 days. Lifespan decreased at lower food levels reaching 2.5 days at 0.05 × 106 cells/ml. Short lifespans also were recorded at food concentrations above 1.0 × 106 cells/ml, probably the result of accumulation of algal metabolic products that are toxic to rotifers. Fecundity also peaked at 1.0 × 106 cells/ml, with a mean of 17 offspring per female. In contrast, at 0.05 × 106 cells/ml, lifetime fecundity was only 0.5 offspring per female, whereas three offspring per female were produced at 5.0 × 106 cells/ml. Similar results have been recorded for other brachionids fed Chlorella at densities of 106 cells/ml. Intraspecific differences in survival among strains vary considerably. For example, in B. plicatilis, mean lifespan for three strains all cultured at 25 °C ranged from about 6 to 14 days. Because these strains were collected from different habitats but were acclimated to, and tested in, a common Chapter | 13 255 Phylum Rotifera FIGURE 13.31 Life histories of Brachionus calyciflorus females grown at high (H) and low (L) food concentrations. Upper panel: amictic females producing diploid female eggs; middle panel: unfertilized mictic females producing haploid male eggs; bottom panel: fertilized females producing diploid diapausing eggs. Numbers in parentheses are typical ranges of egg production for each female type. Percentage of life for all female types calculated based on setting the life span of unfertilized mictic females at the high food level to 100%. Data reanalyzed from Xi et al. 2001. Internat. Rev. Hydrobiol. 86, 211–217. environment, the observed differences must be genetic. (Growing or culturing organisms with different genotypes or from different habitats is typical in ecological research and is known as the common garden technique.) The three different female types (amictic, and unfertilized and fertilized mictic) differ markedly in their agespecific survival and fecundity. In studies on species in the genus Brachionus, unfertilized mictic females usually lived longer and produced more offspring than amictic females, depending on food levels. Fertilized mictic females produced by far the fewest embryos (Figure 13.31). The greatly reduced fecundity of fertilized females is typical of other rotifer species and is likely due to higher energy requirements for the formation of amictic or diapausing embryos over unfertilized mictic embryos (male eggs). Species with different phenotypic growth forms (e.g., spined and unspined) can possess very different intrinsic rates of increase under identical culture conditions. For example, r for spined and unspined forms of K. testudo can differ significantly, but that difference depends on food concentration. At low food levels (0.1–1 μg/ml dry algal biomass), there is no significant difference in r for the two forms of this rotifer (∼0.05–1). At higher food concentrations (1.2–8 μg/ml), however, the unspined form had much greater values of r than the spined form (0.3–0.4 vs 0.15, respectively). Presumably, similar phenomena will be found in other species in which there are spined and unspined forms. The reader is referred to Gilbert (2013) for a recent review of this topic. When life-history patterns of zooplankton are compared, rotifers have higher r than either cladocerans or copepods; r ranges from 0.2 to 1.6, 0.2 to 0.6, and 0.1 to 0.4 offspring per female per day, respectively. Rotifers achieve their high population growth rates by short development times that more than compensate for their small clutch sizes. Rotifers also show the greatest response to increased temperatures. The high population growth rates of rotifers may be an adaptive response to predation, against which most rotifer species have relatively few strong defenses. As a result of these characteristics, rotifers are regarded as being able to quickly exploit new conditions. Dynamics of Field Populations Annual cycles of natural rotifer populations have been characterized for several species, but to achieve a good appreciation of the population dynamics of rotifers long-term studies are needed. Rotifer dynamics have been followed for several years in Neusiedlersee, a shallow, well-mixed Austrian lake (Figure 13.32). In this lake some species, like K. quadrata, have relatively stable population sizes and occupy the lake nearly continuously. In contrast, abundances of other species such as Filinia longiseta (Ehrenberg, 1834), Hexarthra fennica (Levander, 1892), and Rhinoglena fertoeensis (Varga, 1928) fluctuate much more, whereas others (e.g., A. fissa) are intermittent. Fluctuation of rotifer abundance over two or three orders of magnitude during a seasonal cycle is typical of many natural populations. Another important long-term study of zooplankton is that of Lake Washington (Washington, USA). This work began in 1933 and continued with intermittent sampling until 1963, when a more intensive sampling schedule was implemented. Over one 13-year period, the population level of K. cochlearis varied by a factor of 12-fold, while another species, C. hippocrepis, which was present only sporadically since 1933, became important (Figure 13.33). Techniques have been developed to characterize the dynamics of natural zooplankton populations. The intrinsic rate of increase of a population (r) is the difference between instantaneous birth (b) and death (d) rates, where: r=b−d Although more complex models can be constructed, the value of r for a given population may be estimated from the equation: r = (ln Nt − ln N0 ) /t This procedure requires two successive estimates of population size (N0 and Nt) separated by time interval t. The estimate of r is based on several assumptions about the population including that it is growing exponentially with a stable age distribution. One also can estimate the birth rate 256 SECTION | III Protozoa to Tardigrada FIGURE 13.32 Phenologies of several important rotifer species in Neusiedler See, Austria. Rotifer population densities are reported as individuals per liter; note variations in the population scales. Gaps indicate missing data. Panels, from top to bottom for both columns: Total rotifer density; Rhinoglena fertoeensis; Brachionus calyciflorus; Keratella quadrata; Keratella cochlearis; Notholca acuminata; Anuraeopsis fissa; Polyarthra vulgaris/dolichoptera; Asplanchna girodi; Synchaeta tremula/oblonga; Filinia terminalis; Filinia longiseta. The Polyarthra and Synchaeta species combinations are due to the difficulty in diagnosing these species in preserved samples. Data courtesy of Alois Herzig, Biologische Station Neusiedlersee, Austria. Chapter | 13 Phylum Rotifera FIGURE 13.33 Population dynamics of two planktonic rotifers (Keratella cochlearis and Conochilus hippocrepis) in Lake Washington over a 13-year period. Adapted from the Lake Washington database of W. ‘Tommy’ Edmondson, as funded by the Andrew Mellon Foundation; data courtesy of Daniel Schindler, University of Washington, WA, Seattle. in this population by counting the number of eggs carried per female. The finite hatching rate is: B = E/D, where E is the number of eggs per female and D is the developmental rate at a specific temperature. The value E is determined directly from samples of the population, and values of D by observing hatching of eggs from samples brought back to the laboratory and monitored for egg hatching. When E and D have been calculated, the intrinsic birth rate, b, can be estimated from the following equation: b = ln (E + 1) /D With these data, it is possible to estimate death rate, d, by subtraction: d=b−r The egg ratio technique makes it possible to predict growth of natural rotifer populations in which reproduction by amictic females is parthenogenetic. A few simple measurements allow ecologists to estimate important population parameters that summarize processes of birth and death occurring in the population. Thus, this technique has proved useful for investigating the population dynamics and secondary production of zooplankton. As noted above, the dynamics of natural rotifer populations are affected by a number of environmental factors, including temperature and food quantity and quality; however, exploitative and interference competition, predation, and parasitism also are important. Temperature is a major factor affecting fertility, mortality, and developmental rates. In general, higher temperatures do not increase the number of offspring produced per female; instead, they shorten birth intervals by decreasing the development time. Lifespan also is reduced, but the net effect of higher temperatures is an increased population growth rate. In laboratory studies, 257 FIGURE 13.34 Synergistic effects of temperature and food density on the intrinsic rate of population increase (rm) of Brachionus calyciflorus. From Starkweather, (1987), with kind permission of the author and Elsevier Science and Technology Books. higher temperatures and elevated food levels combine synergistically to increase mean rate of population increase of B. calyciflorus (Figure 13.34). Other abiotic environmental factors, such as oxygen concentration, light intensity, levels of pesticides, and pH, also influence rotifer population dynamics. Food availability is a major biotic factor regulating rotifer population growth, and species have markedly different food requirements for reproduction. Threshold food concentration, the food level where population growth is zero, was determined for eight species of planktonic rotifers by Stemberger and Gilbert (1985). Small species such as K. cochlearis had lower threshold food concentrations than did larger species such as A. priodonta or S. pectinata. The logarithm of the threshold concentration was positively related to the logarithm of rotifer body mass, so the smallest species had the lowest food thresholds. The food concentration required to support 50% of the maximum population growth rate (r/2) varied 35-fold among the eight rotifer species and also was positively related to body mass. Small rotifer species, therefore, appear to be better adapted to food-poor environments, because those species possess low food thresholds. Larger species, in contrast, are better adapted to food-rich environments, where they have higher reproductive potentials. Natural rotifer populations often are food limited. In one study, populations of four species in two mountain ponds, K. cochlearis, Polyarthra vulgaris Carlin, 1943, Synchaeta oblonga Ehrenberg, 1832, and Synchaeta sp. grew faster when provided supplemental food in enclosures. The intensity of food limitation changed rapidly, indicating strong temporal variation in resource availability. Although food limitation was common, other factors such as intraspecific resource competition also strongly affected population dynamics. Population Dynamics in Chemostats Studies of field populations are hampered by many confounding factors, including sampling errors and variability 258 SECTION | III Protozoa to Tardigrada in both abiotic and biotic factors that can affect rotifer growth. Thus, chemostats are useful tools for probing the details of population dynamics in environments in which abiotic factors (e.g., temperature, light, medium chemistry) and biotic factors (e.g., food quantity and quality) can be more rigorously controlled (Boraas, 1983). Such work has explored the conditions required for populations to establish an equilibrium level or to cycle. In some chemostat experiments, populations have rapidly decreased or lost their capacity for sexual reproduction. This suggests a strong selection against the sexual cycle (see Serra and Snell, 2009). expression) has been used to identify genes involved in mate recognition and thermotolerance. Comparison of sequence variation has been used to query genes for signatures of positive or purifying selection. Finally, next- generation sequencing technologies provide an unprecedented opportunity to sequence entire genomes or transcriptomes, and have been applied to characterize genes and gene families expressed under conditions such as dormancy. Genetic Variation Initially, indirect inferences of high levels of genetic variation were derived from mating assays. These showed reproductive boundaries or inbreeding depression. However, with the advent of electrophoretic techniques, researchers began to study levels of allozyme variability among localities. Some of these studies revealed high levels of genetic differentiation, whereas other studies suggested dominance of a few molecular types. Later tools for sequencing markers (e.g., microsatellites, mitochondrial cytochrome c oxidase genes, and rDNA) have refined the ability to characterize genetic variation within and between populations, to discriminate among morphologically similar or identical species (i.e., cryptic species), and to compare evolutionary relatedness (i.e., phylogenetic analysis). Sequencing and karyotype analyses also have increased our understanding of the genetic structure in rotifers, discovering, for example, the tetraploid genome of bdelloid rotifers, in contrast to the diploid condition of monogononts. A prominent theme in rotifer genetic analyses is the recognition of species boundaries and understanding the speciation process. Molecular tools have improved our ability to distinguish members of cryptic species complexes, and description of new species is ongoing. For example, the B. plicatilis species complex is composed of three main morphotypes (the larger B. plicatilis, intermediate-sized Brachionus ibericus Ciros-Peréz, Gómez and Serra, 2001, and smaller B. rotundiformis), as well as other sibling species. This group presents a classic example of morphological stasis, combined with niche partitioning thought to facilitate species coexistence (Gómez et al., 2002). Complexes of sibling species are probably common throughout the Rotifera, making molecular analyses critical for accurate characterization of diversity. Genetic variation among species of bdelloids presents a particular puzzle, as these asexual animals cannot be discriminated by the classic biological species definition of reproductive incompatibility. Novel techniques hold promise not only for advancing understanding of species relatedness and genetic variation, but also for understanding functional diversification. Application of interference RNA (targeted suppression of gene Research on food selection by rotifers began by simple microscopic observation of rotifers feeding and by correlating the density of natural populations of rotifers to the various types of foods available in the habitat. However, to obtain more precise information on food selectivity, workers examined the relative number of food particles ingested by rotifers to particle concentration in the water. This technique has been employed for both laboratory cultures and natural populations of rotifers, and can use both natural foods (i.e., algae, bacteria, yeast, protists) and artificial materials (e.g., latex microspheres). To determine feeding rates, known concentrations of food particles are provided to rotifers, which are allowed to feed for short periods of time. From that, the number of cells ingested per animal per unit time (feeding rate) or the volume of water cleared of all particles per unit time (clearance rate) can be calculated. Natural foods also have been labeled using radioisotopes, but loss of the radioactive label from the food can result in significant errors when determining feeding rates. Bacteria and algae also can be labeled by using stable isotope tracers or by using fluorescent dyes. However, even the simple procedure of adding a few drops of food suspension to a Petri dish with rotifers may lead to interesting observations concerning how rotifers process food. For example, using powdered carmine, Wallace (1987) demonstrated that adult S. socialis do not act independently of one another when in a colony. Instead, the corona of the animals in a large section of the colony all face in the same direction for several minutes. Animals exhibiting this behavior form a group called an array. The distribution of arrays within a colony determines where the dominant water currents flow to the colony, how the water currents flow around the colony, and where the currents leave the colony. Rotifers feed in ways that are directly related to their general life history. Although most planktonic rotifers, such as Asplanchna, Brachionus, Polyarthra, and Rhinoglena, swim at similar rates through comparable areas of the water column, the ways in which they encounter and capture food can be quite different. Brachionus uses its swimming currents to sweep small food particles into the buccal region for processing; this is often termed filter or suspension feeding. Ecological Interactions Foraging Behavior Chapter | 13 259 Phylum Rotifera Asplanchna, on the other hand, does not use its swimming currents to gather food. Once this rotifer contacts a potential food item with its corona, it may or may not attempt to ingest the item, based on such factors as hunger level and the size and type of prey. Some benthic rotifers creep along algal filaments and feed by piercing the filament and sucking the cytoplasm from the cells; examples include Notommata copeus Ehrenberg, 1834 and Trichocerca rattus (Müller, 1776). Sessile rotifers capture food in one of two ways, depending on the family. Members of the family Flosculariidae (e.g., Floscularia, Ptygura, Sinantherina) create feeding currents in a manner similar to the planktonic suspension feeders. In contrast, all collothecid rotifers are ambush raptors (Collotheca, Stephanoceros). Once a prey enters the infundibular region of the corona of these rotifers, long setae (Collotheca) or arms (Stephanoceros) fold over the prey, capturing it much like the action of a Venus flytrap (Figure 13.8). All members of the family Atrochidae lack setae. For example, in C. vorax, an enlarged, umbrella-like corona folds over the prey to capture it (Figure 13.9). In both cases, prey is pushed through the rotifer’s mouth and into the proventriculus where it is stored until the mastax transfers it into the stomach. When prey are particularly abundant, several live organisms may be seen in the proventriculus. Occasionally, predatory rotifers such as Cupelopagis may become so engorged that attempting to ingest another prey item results in the loss, from the proventriculus, of one previously captured. However, not all potential food items are consumed. By observing individual B. calyciflorus in various densities of suspended food particles, it is possible to see that this rotifer regulates the ingestion of food particles by three different mechanisms. First, this species can use pseudotrochal cirri to screen certain large particles away from the mouth. Second, particles collected by the corona may be rejected by cilia within the buccal tube: i.e., particles that gain entrance to the oral cavity may be removed by a reversal of the action of oral cilia. Finally, the mastax also may actively reject particles. Members of the genera Asplanchna and Asplanchnopus also use their trophi to remove empty carapaces of hard-bodied prey such as other rotifers and cladocerans from their stomachs. Although the diets of some rotifers are highly specialized, many species consume a wide variety of both plant and animal prey and may be described as generalist feeders. However, even generalist feeders may vary in their method of food acquisition. For example, the herbivorous genera Brachionus and Ptygura, which process many tiny particles in rapid succession (microphagous), have been seen to consume small ciliates. In contrast, the main food source of Asplanchna is often large algae; however, despite its herbivorous diet, this genus often is considered predatory. Perhaps a more informative term for this genus is raptorial. Thus, in rotifers, clear distinctions between what constitutes a predator or herbivore can be fuzzy. Therefore, it may be more constructive to consider the ways in which rotifers encounter and process food than what they eat. This form of analysis is more informative about a species’ functional role in the environment. Thus, with these distinctions in mind, it is possible to characterize the dominant trophic type by using an index called the Guild Ratio (GR). This is the ratio of raptorial (R) to microphagous (M) feeders. For example, compare the trophi illustrated in Fig. 13.10 g to 13.10 i, respectively. The GR, which can be expressed either in numbers of individuals or in biomass (e.g., R# and Rb), is calculated as follows: or in an alternative form, Thus, GR′ values will range between −1 and +1, with values <0 indicating microphagous dominance and values >0 indicating raptorial dominance (Obertegger et al., 2011). Given that the GR′ is determined by means of similar information used to calculate traditional diversity indices (e.g., S and H′), it can provide another way to follow trends in community structure. Molinero et al. (2005) used a similar approach in their study of changes in the rotifer community of Lake Geneva. This study, which was based on body size, identified a sudden swing toward smaller-bodied rotifers beginning in about 1987. This shift separated two regimes: an early one (1972–1986) characterized by low temperature and high phosphate concentrations, and a later one (1987–1998) with higher temperature and lower phosphate levels. Functional Role in the Ecosystem Three groups dominate the freshwater zooplankton (protists, rotifers, and microcrustaceans), yet it is the microcrustaceans that usually receive the most attention from researchers. Such a disparity is to be expected for two reasons, one trivial and one significant. The trivial reason is that microcrustaceans are easier to observe, identify, and manipulate in laboratory situations. The significant reason is that microcrustaceans commonly account for a greater proportion of the total zooplankton biomass than either protists or rotifers; however, this varies seasonally, and protist and rotifer biomass can be considerably greater. Nevertheless, although protists and rotifers usually have a smaller standing biomass than microcrustaceans, their greater numbers and high turnover rate makes them very important to the trophic dynamics of freshwater planktonic communities, including the microbial loop (see also “Predator–Prey Interactions”). Additionally, studies have shown that the 260 relative importance of all three taxa is often idiosyncratic to the habitat. For example, in one study that examined the biomass of zooplankton in two large shallow lakes, ciliates comprised >60% of total biomass in one lake but only 6% in the other. In the first lake, the remaining zooplankton biomass was composed of cladocerans (17%), rotifers (10%), and copepods (9%), whereas in the second lake the pattern was cladocerans (38%), rotifers (20%), and copepods (36%). The importance of each species to the community also can be visualized by ordering the species according to their relative contribution to either community abundance or biomass. The cladoceran Bosmina longirostris was ranked first in density and biomass of the metazoan zooplankton in a Brazilian reservoir. However, although the rotifer P. vulgaris was second in density, it was sixth in biomass (Figure 13.35). In fact, the mean change in position of the species based on a ranking of number of individuals to one based on biomass was −8.2, +8.2, and +9.3, for rotifers, cladocerans, and copepods, respectively. Thus, rotifers, which dominated the top 10 positions by abundance in this example, did not contribute proportionally to community biomass. Of course, knowledge of densities and biomass alone is insufficient for understanding the functional roles of each species in the community. Additional required information includes energy consumption and transfer through food webs. Feeding rates of zooplankton generally are referred to as filtration or clearance rates, and are measured as microliters of water cleared of a certain food type per FIGURE 13.35 Species rank–abundance and rank–biomass curves of the summer zooplankton community (microcrustaceans and rotifers) of Ponte Nova Reservoir, São Paulo, Brazil. Symbols: closed figures = abundance; open figures = biomass; diamonds = rotifers; circles = cladocerans; squares = copepods (including juveniles). Numbers indicate the first four rotifers with the highest numerical ranks. Data calculated from Table 2 of Sendacz et al. (2006). SECTION | III Protozoa to Tardigrada animal per unit of time (i.e., μl/animal/h). Usually, rotifer clearance rates are lower than those of cladocerans and copepods, although the rates depend heavily on food type, temperature, and animal size. For most rotifers, clearance rates are commonly between 1 and 10 μl/animal/h, whether determined in the laboratory or in the field. However, a few species can achieve levels exceeding 50 μL/animal/h. Using estimates of clearance rates, it becomes apparent that even moderate-sized rotifers with body volumes of about 10−3 μl process enormous amounts of water with respect to their size: >103 times their own body volume each hour! Ingestion rates (biomass consumed per animal per unit time) also are very high for rotifers. An adult rotifer may consume food resources equal to 10 times its own dry weight per day. If their assimilation efficiencies (i.e., assimilation divided by ingestion) are between 20% and 80%, rotifers can convert a good deal of their food to animal biomass that may be passed on to the next trophic level. Although microcrustaceans generally have higher clearance rates than rotifers (∼10–150 and 100–800 μl/animal/h for cladocerans and copepods, respectively), rotifers can exert greater grazing pressure on phytoplankton than some small cladocerans. In one study in a small eutrophic lake, K. cochlearis populations accounted for about 80% of the community grazing pressure on small algae during the year. This study also showed that K. cochlearis had clearance rates about 5–13 times higher per unit biomass than the cladoceran B. longirostris. Therefore, under certain conditions, rotifers may be important competitors to small, filter-feeding microcrustaceans and are important in nutrient recycling in aquatic systems. Furthermore, rotifers can alter the species composition of algae in certain systems. Studies have shown that intense feeding by Brachionus rubens Ehrenberg, 1838 can cause a shift in the dominant algal species from Scenedesmus to the spined algae, Micractinium. Apparently this shift is based on the inability of B. rubens to consume algae with protective spines. Although generally not ingested, cyanobacteria are increasingly recognized as playing an important role in determining zooplankton species composition. Large cladocerans are more sensitive to cyanobacteria (e.g., Anabaena spp.) than rotifers. The mechanism for this differential sensitivity to cyanobacteria toxicity is based on different tendencies to ingest filamentous cyanobacteria and different physiological tolerances to their toxins. Thus, having the ability to reproduce rapidly, rotifers may account for 50% or more of the zooplankton production, depending on the prevailing conditions. This production, in turn, can be an important food source for other rotifers, Asplanchna, cyclopoid and calanoid copepods, malacostracans (Mysis), zebra mussels, aquatic mites, insect larvae (Chaoborus) and adults (Buenoa), and small fishes. Abundance and species composition of rotifers often reflect the trophic status of lakes. For example, numerous Chapter | 13 261 Phylum Rotifera studies have reported changes in the maximal, totalpopulation density of several orders of magnitude when lakes were subject to intense eutrophication. Individual species sometimes undergo dramatic population changes during those periods. This was observed in the zooplankton of Lake Constance, where the density of Asplanchna increased its maximum population level 280-fold over a period of 28 year. However, in other lakes, dramatic population declines have been seen. In the years in which in Lake Washington had elevated concentrations of dissolved phosphorus, low water transparency, and high algal densities, K. cochlearis was abundant; however, as these water quality parameters improved, the population of this rotifer declined dramatically. Overall, there was at least a 20-fold increase and then a decline during a period of 15 years. Studies of the interactions of rotifers with other organisms will probably continue to receive attention in the future, especially predator–prey interactions, exploitative and interference competition among rotifers and other herbivorous zooplankton, life history strategies, and the toxic effects of cyanobacteria, dinoflagellates, and diatoms. Moreover, little detailed work has been done to examine the concept of functional complementarity in rotifer communities. This hypothesis argues that, within the constraints of niche requirements, a decline in the population levels of some species is offset by a rise in others, a concept termed compensatory dynamics (Fischer et al., 2001). The idea that the GR′ of communities can deviate widely over a season (see above) and that some rotifer communities appear to exhibit compensatory dynamics needs to be explored in greater detail. extinction after 2–3 weeks (Figure 13.36). Daphnia is unaffected by the presence of rotifers. Of course, protist–rotifer competition for bacterial food also can be important. Population growth of certain rotifers also is inhibited by Daphnia through interference competition. For example, in the presence of Daphnia, K. cochlearis suffers mechanical damage (i.e., is killed, wounded, or loses its eggs) when swept into the branchial chamber of the daphnids; the rate at which rotifers were killed is apparently proportional to daphnid body length. On occasion, Keratella may be found in the guts of Daphnia and Cypris (Figure 13.37), indicating a surprising pathway for trophic interactions (Gilbert, 2012). These microcrustaceans have the greatest impact on Keratella populations when larger than 2 mm; similar effects have been reported for other cladocerans (e.g., Scapholeberis kingi). Of course, interference and exploitative competition may occur simultaneously. Predator–Prey Interactions Predation is another important regulatory factor in rotifer population dynamics, as rotifers are prey for several aquatic predators including protists, other rotifers, insects, cladocerans, copepods, and planktivorous fish. From many studies, we know that predation affects rotifer population dynamics both directly, by contributing to mortality, and indirectly, as a selective force shaping rotifer morphology, physiology, and behavior. One area that deserves additional study is the trophic interactions of microbes, protists, and Competition with Other Zooplankton As noted before, rotifers, cladocerans, and copepods often compete for limited food resources and, in general, rotifers are relatively poor exploitative competitors because their clearance rates are usually many times lower than those of daphnids. In addition, rotifers also have a more limited size range of particles that they can ingest compared to cladocerans and are less resistant to starvation. Thus, cladocerans generally have broader food niches than rotifers in terms of food type and size, and through direct competition may suppress rotifer population growth. However, this outcome may be reversed when a sufficient quantity of suspended sediments is present in a lake. Exploitative competition between rotifers and daphnids is readily demonstrated when these zooplankton are grown in single and mixed cultures. Brachionus calyciflorus and Daphnia pulex both grow well on the alga Nannochloris oculata in single species cultures. When both species are present, however, Daphnia removes an increasingly larger proportion of algal cells until the rotifers gradually starve to FIGURE 13.36 Competition between Brachionus calyciflorus and Daphnia pulex. Brachionus and Daphnia were grown in single species (closed symbols) and mixed-species (open symbols) batch cultures at 20 °C, daily renewed with 5 × 106 Nannochloris cells per milliliter. Population size (Y axis) is the number of individuals in the 80-mL culture. Error bars (±1 SE) are visible when they exceed the size of the symbols. Figure redrawn from Figure 1: Gilbert (1985), with kind permission of the author and the Ecological Society of America. SECTION | III 262 rotifers within the microbial loop (Figure 13.38) (see also Parasitism on Rotifers below). Most rotifers are transparent and quite small; some are smaller than many ciliates (ca. 60–250 μm long). Although these features benefit planktonic rotifers by reducing their visibility to fish, a small body size renders rotifers more vulnerable to those invertebrate predators that are tactile feeders. Many rotifers produce a thickened integument (lorica) and/or spines and other projections, or carry their eggs, all of which have been shown to reduce the ability FIGURE 13.37 Fecal pellet from Cypris pubera cultured with Keratella tropica. Visible in this pellet are at least five loricas of K. tropica (numbers). Original photomicrograph courtesy of J.J. Gilbert, Dartmouth College; see also Gilbert (2012). Protozoa to Tardigrada of predatory zooplankton to prey upon them (Wallace and Smith, 2009). On the other hand, small size also appears to be a deterrent to predation. In laboratory cultures, Sarma and Nandini (2007) demonstrated that, despite its small body size (70 μm), Anuraeopsis fissa was not consumed at the same rate as two alternative brachionid prey by either A. brightwellii or A. sieboldii. In fact A. sieboldii ignored A. fissa, although it was consumed by A. brightwellii. Rotifers have other means of reducing predatory pressures. Spines are produced in some rotifers (e.g., B. calyciflorus, Figures 13.13 and 13.39) in response to a build-up of soluble substances released by invertebrate predators such as Asplanchna and several genera of copepods Epischura, Mesocyclops, and Tropocyclops. This phenomenon is another type of chemical signaling, except that the communication is between predators and prey rather than among conspecifics as in quorum sensing. Polymorphic spine production has been observed in B. calyciflorus, Brachionus urceolaris Müller, 1773, F. longiseta, K. cochlearis, Keratella slacki Bērziņš, 1963, and K. testudo. The importance of spined morphotypes is a significant reduction in capture and ingestion by invertebrate predators by making the rotifer more difficult to manipulate and swallow. The presence of spines on B. calyciflorus is a good example of the phenomenon (Figure 13.13) that works as follows. When disturbed by a potential predator, B. calyciflorus will retract its corona and, by doing so, increases the hydrostatic pressure within its pseudocoelom. (a) (c) (b) (d) FIGURE 13.38 Examples of rotifers in the microbial loop. (a) Heliozoan with ingested Lecane. (b) Ciliate (Frontonia) with ingested Lecane. Photomicrograph courtesy of John Maccagno. (c) Asplanchna with two Keratella within its gut. (d) Several individuals of an unidentified microbe within the lorica of a dead Euchlanis. This culture crashed within three days; all of the dead rotifers were infested like this one. Key: r = rotifers; bar = 100 μm. Chapter | 13 Phylum Rotifera The elevated pressure causes the posteriolateral spines to swing forward and outward (Figure 13.39), making it more difficult for a predator to manipulate. For Asplanchna, this change is sufficient to prevent ingestion after capture. After a period of time, during which Asplanchna attempts to swallow B. calyciflorus (usually >60 s), the predator will release (reject) the prey, which then swims away unharmed. In this example, Asplanchna releases a biochemical cue (an allelochemic, called a kairomone) that initiates a developmental change in subsequent generations of B. calyciflorus (spine production), reducing the effect of predation. Some forms of K. cochlearis also possess posterior spines that make them much more likely to be rejected after capture by Asplanchna girodi than unspined forms. Unfortunately, the cost of spine production in rotifers, either in terms of developmental costs of producing the spines or increased energetic demands on swimming by increased mass, has not been fully resolved (Gilbert, 2013). Sometimes when Asplanchna or a related species (Asplanchnopus) swallows a spined form, the spines become lodged in the gut of the predator and may even puncture the delicate tissues. Presumably, both predator and prey die when that happens. Similarly transparent mucus sheaths produced by some planktonic, colonial rotifers (e.g., Conochilus, F igure 13.19(c), and Lacinularia) deter predation; the sheaths make the effective size of the prey too large for the invertebrate predator (e.g., Asplanchna and predatory copepods) without making them more visible to fish. The tubes and sheaths of sessile rotifers also work as refugia (Figures 13.8(a),(d) and 13.19(a)). Some rotifers escape predators by making rapid jumps using a variety of appendages: setous arm-like appendages (Hexarthra, Figure 13.14), paddles (Polyarthra, Figure 13.40), and long setae (Filinia, Figure 13.41). However, apparently Filinia also can use its long setae as foils to ward off predators and not just as lever-arms to initiate a jump. FIGURE 13.39 Extension of the posterolateral spines in Brachionus calyciflorus. Here the spines are extended due to preservation in formalin. In life retraction of the corona causes an increase in the pressure within the pseudocoelom, which results in an outward flexing of the spines. 263 Some rotifers assume a passive posture, displaying what has become known as the dead-man response, rather than fleeing when predators attack (Asplanchna, Brachionus, Keratella, Sinantherina, and Synchaeta). This simple behavior is FIGURE 13.40 Polyarthra. This rotifer possesses long paddle-like appendages that are used in making rapid jumps to escape predators. Photomicrograph courtesy of Martin V. Sørensen, University of Copenhagen. FIGURE 13.41 Filinia. This rotifer possesses long spines that it uses as foils to rapidly fend off predators. Photomicrograph courtesy of Martin V. Sørensen, University of Copenhagen. 264 SECTION | III Protozoa to Tardigrada members of the Scenedesmaceae (Chlorophyta). For example, Scenedesmus obliquus exhibited a logistic dose– response to dilutions of test medium in which B. calyciflorus was incubated, thus indicating the presence of grazingreleased biochemicals, termed infochemicals (Verschoor et al., 2004). Algal defenses such as these can influence the long-term stability of both rotifer and algal populations and, in doing so, steady the population fluctuations often seen in bi- and tritrophic food chains. Rapid evolution also has been documented in long-term chemostats in which rotifers were fed algae from a stock composed of multiple clones. In the presence of rotifer grazing, there is strong selection for algal genotypes that are more digestion resistant; however, this defense comes at a cost of slower population growth (Yoshida et al., 2004). Parasitism on Rotifers FIGURE 13.42 A few individuals of a colony of Sinantherina socialis. Here all the animals have retracted their corona, a behavior that exposes the warts on their anteroventral surface. merely a retraction of the corona into the body and passive sinking. Contraction of the corona stops the animal from swimming, which eliminates the vibrations it produces that may be detected by the predator. In addition this behavior may make the rotifer more difficult to grasp in its turgid state. In Sinantherina spinosa (Thorpe, 1893), this passive posture exposes a group of small spines on its anteroventral body surface that may function in defense against planktivorous fish. To date, only one species of rotifer, S. socialis, has been shown to be unpalatable to small zooplanktivorous fishes (Felix et al., 1995) and certain invertebrates (Walsh et al., 2006). While neither the nature nor the location of the unpalatability factor(s) is known, this colonial species probably possesses a chemical that is held in gland-like structures, called warts, located at the anterior end of the animals (Figure 13.42). Defenses against predators such as spines, mucus sheaths, thickened loricas, and escape movements are energetically demanding. Some species, such as Synchaeta pectinata, are not well defended against predators, but have evolved very high maximal population growth rates that offset mortality from predation. Avoiding potential predators in space and/or in time is another simple yet effective defense mechanism against predators. Some rotifers occupy the habitat at a different time of year from that of an important predator, migrate vertically or horizontally in the habitat, or live in zones with low oxygen concentration, thereby evading predatory pressures altogether. Some algae initiate colony formation as an antipredator defense when rotifers feed on them. This occurs in The importance of parasites in controlling population density in rotifers has not been examined thoroughly, although a few studies have correlated parasitic infection with a decrease in population density of planktonic species. In certain cases, parasites apparently caused the demise of an entire population in a lake within a few days. At least one virus causes high mortality in B. plicatilis aquaculture systems. The s porozoan parasite Microsporidium (Plistophora) frequently infects planktonic rotifers possessing thin loricas, such as members of the genera Asplanchna, Brachionus, C onochilus, Epiphanes, Polyarthra, and Synchaeta. Water temperature seems to be an important mediating factor in the spread of the parasite, as infection rates drop off at water temperatures below 20 °C. In infected rotifers, the pseudocoelom of the animal becomes nearly filled with cysts (Figure 13.43). Several workers have described endoparasitic fungi that attack soil rotifers of the genera Adineta and P hilodina. Some fungi form peg-like adhesive appendages on both small conidia (spores, ca. 30 μm long) and long vegetative hyphae. Once the adhesive pegs attach to a rotifer, they germinate and rapidly colonize the pseudocoelom (Figure 13.44). Other fungi produce spores that initiate parasitic attack when ingested. Another avenue of infection occurs via hypodermic injection of a vegetative cell into the host. Once inside the host, these fungal cells grow into assimilative hyphae, producing more infective cells either inside or outside the rotifer. However, bdelloids apparently free themselves from fungal parasites through anhydrobiosis (Wilson and Sherman 2010, 2013). Rotifers also can ingest the oocysts of parasitic protists important to human health such as Cryptosporidium and Giardia. However, it is not known whether they can significantly reduce the numbers of these parasites in natural conditions. Rotifers also act as vectors of disease agents such as white spot syndrome virus, which have been reported to attack shrimp in aquaculture systems. Chapter | 13 265 Phylum Rotifera FIGURE 13.45 Phoretic association between Brachionus and Daphnia. Photomicrograph courtesy of Elizabeth J. Walsh, University of Texas at El Paso. FIGURE 13.43 Photomicrograph of Brachionus sp. infected with a sporozoan parasite within its body cavity. termed phoresis (G., to carry), to either the daphnid or to the rotifer have not been completely explored. COLLECTING, CULTURING, AND PREPARATION FOR IDENTIFICATION Collecting FIGURE 13.44 An example of a fungal parasite of rotifers. Four rotifers trapped by adhesive pegs on the vegetative hyphae of Cephaliophora. Bar = 100 μm. (Redrawn with permission of George Barron and the Canadian Journal of Botany 61(5):1345–1348.) Rotifers as Parasites A few rotifers have been described as being parasitic on algae, sponges, other rotifers, freshwater oligochaetes, snail eggs, crustaceans, and fishes. However, we really know too little about these associations to classify all of them as parasitic. Members of the genera Brachionus, Limnias, Pleurotrocha, Proales, and Ptygura have been known to make temporary attachments to either invertebrates or vertebrates. In fact, it is not uncommon to find the carapace of Daphnia colonized by numerous individuals of B. rubens (Figure 13.45). The consequences of this phenomenon, Collecting rotifers does not require complex or expensive equipment (Wallace et al., 2006; Wallace and Smith, 2009). One can almost always collect several species of planktonic rotifers by towing a fine-mesh (25–50 μm) net through any body of water. It is important to note that nets with larger mesh sizes (≥63 μm) tend to miss small-bodied forms (Chick et al., 2010). Productive lakes and ponds usually provide especially good sampling sites, but because fine-mesh nets can clog easily in these habitats, the water may need to be prefiltered through a net with a larger mesh size. More elaborate equipment such as closing nets and the Clarke-Bumpus sampler work well, but they are not necessary unless required by a specific sampling protocol. Water collected by discrete sampling devices (e.g., Van Dorn sampler, Kemmerer bottle, plankton trap, submersible pump) is then filtered. In weedy areas, a dip net or flexible collecting tube, called a water core, are very useful. Another simple method to collect rotifers is to submerge a 3- to 4-l (1-gallon) glass jar in a weedy region and to arrange loosely a few aquatic plants in it before retrieval. In the laboratory, place the jar near a subdued light source such as a north-facing window or a low-intensity lamp. Rotifers that swim to the surface on the lighted side may be removed using a transfer pipette. Certain aquatic plants such as Elodea, Myriophyllum, and filamentous algae are SECTION | III 266 good substrata to examine for the presence of sessile rotifers, but Utricularia usually provides the richest diversity (Edmondson, 1959; Wallace, 1980). Plants with highly dissected leaves may be examined in small dishes using a dissecting microscope. Broad-leaved plants must be cut into strips and examined on edge. The upper few centimeters of moist sand taken just above the water line along a lake or a marine shoreline or the hyporheic interstitial zone of a streambed usually provides several species of rotifers. Unfortunately, very few studies have been conducted on rotifers from the psammon, in part because of the difficulty in separating the organisms from the sand. Recent studies have revealed a remarkable diversity (35–85% of the fauna) and abundance (up to 105 individuals/l) of rotifers in this habitat. Under certain conditions, rotifers may be found at depths of up to 60 cm into the interstitial. Sediments collected from the bottom using a dredge, coring apparatus, or suction device usually provide several species. The upper few centimeters of sediments from a core normally contain diapausing embryos that can be induced to hatch within a few days when several milliliters of sediment are incubated at ambient spring or summer temperatures. Dry sediment from desert rock pools also provide material when rehydrated in culture fluid and incubated for several days. Do not overlook laboratory aquaria as potential sources of material. We have found some unusual species in aquaria that had remained almost unattended for months. One might try adding a small amount of sediments from several sites as a way of adding variety to the rotifer community within the aquarium. Sessile rotifers may be present if the aquarium contains aquatic plants recently collected from the field. However, if you attempt to keep sessile forms, be sure to remove snails from the aquarium. Aquatic plants from hobby shops may bring new rotifer taxa to your aquarium, but the plants themselves may be alien species. Exercise care so that they are not released into the environment. Populations of rotifers may be maintained or even increased by adding a small amount of food once or twice a week (see Culturing), but keep the aquarium aerated. Aquarium filters that use fibrous materials to remove suspended materials may reduce the rotifer population. Once the collection has been made, it should be examined alive as soon as possible. It is generally a good idea to place live samples in jars over ice for the return trip to the laboratory, although we have found that a few species suffer when cooled (i.e., collections made from warm waters). Anesthetizing rotifers in the field has proved to be difficult (see below). Three preservatives are commonly used to preserve rotifers: formalin and Lugol’s iodine (I2KI) at concentrations of 5% or less, and ethanol at about 30–50%. (Higher concentrations of ethanol are used in preservation for DNA Protozoa to Tardigrada analysis.) Lugol’s has two advantages over formalin. It is less toxic, and it stains the specimens slightly, which makes the animals more visible during sorting. Unfortunately, fixation deforms the specimens, making them difficult, if not impossible, to identify. On the other hand, in the genus Lecane, fixation with formalin is advisable so that one can study the morphology of the lorica. Dyes such as Rose Bengal are commonly used to stain preserved specimens. However, preservation sometimes causes rotifers to stick together and to other zooplankton. This preservation artifact has been misinterpreted as a behavior, as is seen in case of live B. rubens, which attaches to free-swimming Daphnia (Figure 12.45). Culturing Laboratory Culture Many species of rotifers have been cultured for research, most notably in the genus Brachionus, both freshwater (B. calyciflorus and B. rubens) and saline (B. plicatilis). Because of their extensive use, these species have been referred to as the white mice of the rotifer world. The culture systems can be quite simple, using only small vessels such as depression slides, watch glasses, plastic tissue culture plates (Figure 13.46(a)–(b)), and small beakers or flasks (Figure 13.46(c)). However, larger systems are often used (Figure 13.46(d)), and, in aquaculture, very large systems are employed (Figure 13.46(e)–(f)). Wallace et al. (2006) provides a summary of procedures for culturing rotifers for research or aquaculture settings. Carlson (2000) provides directions for construction of a simple system for the hobbyist, which can be scaled up for research or large-scale culture (see Lawrence et al., 2012). Culture vessels that have had contact with formalin should not be used, as a residue of this toxin is thought to remain attached to both glass and plastic. Some species easily adapt to artificial conditions, attaining densities of >105 individuals per liter in a few weeks, whereas others seem impossible to keep even for short periods. Most cultures need regular maintenance a few times per week (e.g., changing the medium, feeding, cleaning the vessels). However, some species require little care. For example, some bdelloid species (e.g., Habrotrocha rosa, which is found in pitcher plant traps) are easily cultured in dilute suspensions of powdered baby food or crushed dried pet food. In this case, decomposition of the food source provides bacteria for the rotifers. In a similar manner, the techniques used to culture protists (e.g., making extracts and infusions of various grains, hay, manure, soil) have been adopted for the culture of some rotifer species with great success. Such cultures may be ignored for days and perhaps weeks at a time without loss of the culture. Nevertheless, most researchers grow the food needed to culture their Chapter | 13 267 Phylum Rotifera (a) (b) (c) (d) (e) (f) FIGURE 13.46 Different scales in culturing rotifers. Small-scale laboratory cultures: (a) Plastic tissue culture plates are common for small studies. Here, a 12-well plate (effective volume ≤5 ml) is shown, but 48-well plates (∼1 ml) are used to culture individual animals. (b) Plastic tissue culture flasks are used to achieve slightly larger populations (ca. 250 ml). Medium-scale laboratory cultures. (c) In this two-stage culture system, algae are grown in plastic bags on one shelf (numbers indicate relative age; oldest = 1), which is slowly supplied to the small columns (∼575 ml) on the lower shelf by gravity feed or a pump. Larger laboratory systems: (d) A single-stage culture system in which algae are grown in 250-l plastic columns and then a starter population of rotifers is added. Aquaculture scales: (e) High-density mass culture (>1 m3) achieve up to 4000 individuals/l (Nagasaki Prefectural Institute of Fisheries). (f) Mass culture pools (50 m3) achieve up to 400 individuals/l (Japan Sea Farming Association). Photographs (e) and (f) courtesy of Atsushi Hagiwara, Nagasaki University. target species separately and feed it to the culture at regular intervals, usually 2–3 times per week. Although more costly, commercial products such as Roti-Rich® (Florida Aqua Farms, Inc.) and Sparkle® and related products (INVE Aquaculture®) give excellent results, especially where large numbers of rotifers are required. However, do not overlook the prospect of large populations suddenly arising in fish tanks. One of us (R.L.W.) followed the population dynamics of the sessile rotifer C. vorax (ca. 200–1100 μm) on the sides of a 115 L (50-gal.) aquarium for nearly 7 years; during that time, the density varied from <0.1 to ∼20 individuals per square centimeter. Most rotifer cultures are maintained as xenic systems without much of a problem. In fact, sometimes rotifers are found contaminating cultures of protists, microcrustaceans, etc., that have been provided by commercial biological supply companies. Rotifers also have been maintained under axenic or monoxenic culture conditions, but these require much more effort. Sophisticated culture techniques using single- and two-stage chemostats have been discussed elsewhere in this chapter. Aquaculture Many small fish are well adapted to locate, pursue, capture, and ingest microzooplankton, including rotifers. These organisms are easy prey because they swim slowly and frequently lack sufficient predator defenses. Aquaculturists have exploited this important relationship between planktivorous fish and rotifer prey in intensive aquaculture systems in both freshwater and marine systems (Figure 13.46(e)–(f)). This field has developed into a major 268 technical discipline in several countries including China, India, Israel, Japan, and several countries in Europe. Most of this work has the practical goal of determining the correct biotic and abiotic factors necessary to maintain mass cultures of rotifers. The rotifers are then provided as the first food for larval stages of crustaceans and fishes. In general, rotifers are highly nutritious, and their biochemical composition can be furthered improved by specialized diets. Most systems for mass culturing of rotifers are simple batch cultures capable of producing kilogram quantities of rotifer biomass each day. SECTION | III Protozoa to Tardigrada crushing or distorting the specimen with the cover glass. Both objectives may be accomplished with a compression microscope slide. If a compressor is unavailable, then tiny pieces of broken cover glass (not recommended) or little clay corner supports (recommended) work well to elevate slightly the cover glass. If the clay supports are too high, a slight pressure from a pencil on each corner will reduce the height of the cover glass to the desired level. With some practice, one can trap a planktonic rotifer sufficiently to prevent swimming without undue constriction. Sessile rotifers are handled more easily. Plant material with attached rotifers can be trimmed with iridectomy scissors to a size suitable for placement on a microscope slide. The animal will Preparation for Identification remain in place without the need of compression as long as Unless special precautions are taken before fixation, illoricate the plant material is large enough to act as an anchor. Clay rotifers (especially bdelloids) will contract into a completely supports can help level the cover glass as necessary. unidentifiable lump, making identification difficult, if not Methylcellulose or other viscous agents and fibrous impossible. Such specimens have lost all value for taxonomic material, such as glass wool or shredded filter paper, also purposes, although the trophi still may be useful. Sugar– may be used to impede swimming species. Unfortunately, formalin solutions, which are used to prevent osmotic shock methylcellulose interferes with ciliary function, and fibers in cladocerans (e.g., Daphnia), are not helpful in preventing reduce observation to a game of hide-and-seek. Any lightcontraction in rotifers. Whenever possible, live specimens ing conditions may be used, as long as you are careful not to should be examined first, then fixed to determine the effect overheat the specimens. Strobe lighting provides a marvelof a particular fixative on body shape. In some forms, identi- ous view of ciliary movements, and dark field illumination fied based on the shape of their lorica (e.g., Lecane species), is often spectacular! formalin fixation is required (see above). The final identification of many rotifer species requires Anesthetics ranging from simple carbonated water to examination of the trophi, which, in certain species, may be chemicals that are controlled substances have been used done by compressing an intact animal (e.g., Asplanchna). to anesthetize and sometimes to kill rotifers, but none of However, one may extract trophi from surrounding soft tisthese are universally effective (Edmondson, 1959; Wallace sues using a small volume of bleach (sodium hypochlorite). et al., 2006). Adding minute amounts of powdered MS-222 Some descriptions of this technique use a depression slide, (Tricaine), a fish anesthetic, over the course of 30–45 min but that requires that the trophi be moved to a regular slide to a small drop of water works well for some species. Some after the hydrolysis is complete; this is a very difficult task. researchers use this technique, but instead substitute minute We recommend using a regular microscope slide from the drops of formalin over longer periods of time. This takes start, but without the cover glass. Because the trophi are extreme patience and still may not work. Carbonated water liberated rather quickly when the bleach comes into contact (club soda) may be used to anesthetize rotifers, as well as with the rotifer, it is necessary to find the animal rapidly; microcrustaceans. Apparently the elevated CO2 causes otherwise, it becomes necessary to scan the entire slide asphyxiation, which may be only temporary. Unfortunately, for the small trophi. Be aware that bubbles may form and adding carbonated water to the edge of a cover glass results in obscure your view when bleach comes into contact with production of annoying bubbles that can hinder one’s view. some biological materials. In all work with trophi, it is An alternative approach that bypasses the need for anestheti- important to remember that these structures are very small zation is to use the hot-water fixation technique (Edmondson, (<50 μm) and that they are three-dimensional objects with a 1959). This technique generally gives good results with a particular spatial arrangement among all seven pieces. number of species, once it has been mastered. However, even DNA barcoding has been applied to rotifers with some when great care is taken to anesthetize a rotifer, the striking success, but it has not yet been widely adopted and has a beauty of the living animal is lost by any fixation and mount- long way to go before it will be practical for identification ing procedure. Techniques for anesthetization, preservation, of species. In early studies, the mitochondrial gene for suband mounting of rotifers for examination using light micros- unit 1 of cytochrome oxidase (cox1, also mtCOI) was used copy and transmission and scanning electron microscopy almost exclusively and worked well on a variety of bdelloids have been reviewed by Wallace et al. (2006), and Jersabek and monogononts. Of course, DNA extractions can be done et al. (2010) provides a detailed protocol. successfully only with animals that have been preserved in Observations of live rotifers are not always easily ethyl alcohol (95%). We believe that an experienced taxonaccomplished. The goal is to retard their movements without omist should be consulted to confirm species identifications Chapter | 13 Phylum Rotifera early in genetic work, and certainly well before sequences are deposited in GenBank®. REFERENCES ASTME 1440–91. ASTM, 2012. ASTM. Standard guide for acute toxicity test with the rotifer Brachionus. Am. Soc. Test. Mater. (West Conshohocken, Pennsylvania). Błędzki, L.A., Ellison, A.M., 2002. Nutrient regeneration by rotifers in New England (US) bogs. Verhandlungen Int. Ver. Limnol. 28, 1328–1331. Boraas, M.E., 1983. Population dynamics of food-limited rotifers in a twostage chemostat culture. Limnol. Oceanogr. 28, 546–563. Boschetti, C., Leasi, F., Ricci, C., 2011. Developmental stages in diapausing eggs: an investigation across monogononts rotifer species. Hydrobiologia 662, 149–155. Boschetti, C., Ricci, C., Sotgia, C., Fascio, U., 2005. The development of a bdelloid egg: a contribution after 100 years. Hydrobiologia 546, 323–331. Carlson, S., 2000. How to rear a plankton menagerie. Sci. Am. 283, 84–85. Chick, J.H., Levchuk, A.P., Medley, K.A., Havel, J.H., 2010. Underestimation of rotifer abundance a much greater problem than previously appreciated. Limnol. Oceanogr. Methods 8, 79–87. Clément, P., Wurdak, E., 1991. (Chapter 6) Rotifera. In: Harrison, F.W., Ruppert, E.E. (Eds.), Microscopic anatomy of invertebrates. Aschelminthes, vol. 4. Wiley-Liss, Inc., New York, NY, pp. 219–297. De Meester, L., Gómez, A., Okamura, B., Schwenk, K., 2002. The monopolization hypothesis and the dispersal–gene flow paradox in aquatic organisms. Acta Oecol 23, 121–135. Denekamp, N.Y., Reinhardt, R., Kube, M., Lubzens, E., 2010. Late embryogenesis abundant (LEA) proteins in nondesiccated, encysted, and diapausing embryos of rotifers. Biol. Reprod. 82, 714–724. Dodson, S.I., Arnott, S.E., Cottingham, K.L., 2000. The relationship in lake communities between primary productivity and species richness. Ecology 81, 2662–2679. Dolan, J.R., Gallegos, C.C., 1992. Trophic role of planktonic rotifers in the Rhode River Estuary, spring - summer 1991. Mar. Ecol. Progess Ser. 85, 187–199. Edmondson, W.T., 1945. Ecological studies of sessile Rotatoria, Part II. Dynamics of populations and social structure. Ecol. Monogr. 15, 141–172. Edmondson, W.T., 1959. (Chapter 18) Rotifera. In: Edmondson, W.T. (Ed.), Freshwater biology, second ed. John Wiley & Sons, Inc., New York, NY, pp. 420–494. Epp, R.W., Lewis Jr, W.M., 1980. Metabolic uniformity over the environmental temperature range in Brachionus plicatilis (Rotifera). Hydrobiologia 73, 145–147. Epp, R.W., Lewis Jr, W.M., 1984. Cost and speed of locomotion for rotifers. Oecologia 61, 289–292. Epp, R.W., Winston, P.W., 1978. The effects of salinity and pH on the activity and oxygen consumption of Brachionus plicatilis (Rotatoria). Comp. Biochem. Physiol. 59A, 9–12. Esjmont-Karabin, J., 2003. Rotifera of lake psammon: community structure versus trophic state of lake waters. Pol. J. Ecol. 51, 5–35. Eyres, I., Frangedakis, E., Fontaneto, D., Herniou, E.A., Boschetti, C., Carr, A., Micklem, G., Tunnacliffe, A., Barraclough, T.G., 2012. Multiple functionally divergent and conserved copies of alpha tubulin in bdelloid rotifers. BMC Evol. Biol. 12, 148. 269 Feike, M., Heerkloss, R., 2009. Does Eurytemora affinis (Copepoda) control the population growth of Keratella cochlearis (Rotifera) in the brackish water Darß–Zingst Lagoon (southern Baltic Sea)? J. Plankton Res. 31, 571–576. Felix, A., Stevens, M.E., Wallace, R.L., 1995. Unpalatability of a colonial rotifer, Sinantherina socialis, to small zooplanktivorous fishes. Invertebr. Biol. 114, 139–144. Fischer, J.M., Frost, T.M., Ives, A.R., 2001. Compensatory dynamics in zooplankton community responses to acidification: measurement and mechanisms. Ecol. Appl. 11, 1060–1072. Fontaneto, D., Barbosa, A.M., Segers, H., Pautasso, M., 2012. The ‘rotiferologist’ effect and other global correlates of species richness in monogonont rotifers. Ecography 35, 174–182. Frost, T.M., Fischer, J.M., Klug, J.L., Arnott, S.E., Montz, P.K., 2006. Trajectories of zooplankton recovery in the Little Rock Lake whole-lake acidification experiment. Ecol. Appl. 16, 353–367. Gilbert, J.J., 1967. Asplanchna and postero-lateral spine production in Brachionus calyciflorus. Arch. für Hydrobiol. 64, 1–62. Gilbert, J.J., 1980. Female polymorphisms and sexual reproduction in the rotifer Asplanchna: evolution of their relationship and control by dietary tocopherol. Am. Nat. 116, 409–431. Gilbert, J.J., 1985. Competition between rotifers and Daphnia. Ecology 66, 1943–1950. Gilbert, J.J., 2007. Induction of mictic females in the rotifer Brachionus: oocytes of amictic females respond individually to population-density signal only during oogenesis shortly before oviposition. Freshwater Biol. 52, 1417–1426. Gilbert, J.J., 2012. Effects of an ostracod (Cypris pubera) on the rotifer Keratella tropica: predation and reduced spine development. Int. Rev. Hydrobiol. 97, 445–453. Gilbert, J.J., 2013. The cost of predator-induced morphological defense in rotifers: experimental studies and synthesis. J. Plankton Res. 35, 461–472. Gilbert, J.J., Schröder, T., 2004. Rotifers from diapausing, fertilized eggs: unique features and emergence. Limnol. Oceanogr. 49, 1341–1354. Gilbert, J.J., Williamson, C.E., 1983. Sexual dimorphism in zooplankton (Copepoda, Cladocera, and Rotifera). Annu. Rev. Ecol. Syst. 14, 1–33. Gómez, A., Serra, M., Carvalho, G.R., Lunt, D.H., 2002. Speciation in ancient cryptic species complexes: evidence from the molecular phylogeny of Brachionus plicatilis (Rotifera). Evolution 56, 1431–1444. Halbach, U., 1970. Die Ursachen der Temporal Variation von Brachionus calyciflorus Pallas (Rotatoria). Oecologia 4, 262–318. Halbach, U., 1973. Life table data and population dynamics of the rotifer Brachionus calyciflorus pallas as influenced by periodically oscillating temperature. In: Wieser, W. (Ed.), Effects of Temperature on Ectothermic Organisms. Springer-Verlag, Berlin, pp. 217–228. Halbach, U., Halbach-Keup, G., 1974. Quantitative Beiehungen zwischen Phytoplankton und der Populationdynamik des Rotators Brachionus calyciflorus Pallas. Befunde aus Laboratoriums-experimenten und Freilanduntersuchungen. Arch. für Hydrobiol. 73, 273–309. Hanson, S.J., Schurko, A.M., Hecox-Lea, B., Mark Welch, D.B., Stelzer, C.-P., Logsdon Jr, J.M., 2013. Inventory and phylogenetic analysis of meiotic genes in monogonont rotifers. J. Hered. 104, 357–370. Harring, H.K., Myers, F.J., 1922. The rotifer fauna of Wisconsin. Trans. Wisc. Academy Sci., Arts Lett. 20, 553–662. Hochberg, R., 2006. On the serotonergic nervous system of two planktonic rotifers, Conochilus coenobasis and C. dossuarius (Monogononta, Flosculariacea, Conochilidae). Zool. Anz. 245, 53–62. 270 Hochberg, R., 2009. Three-dimensional reconstruction and neural map of the serotonergic brain of Asplanchna brightwellii (Rotifera, Monogononta). J. Morphol. 270, 430–441. Hochberg, R., Ablak Gurbuz, O., 2008. Comparative morphology of the somatic musculature in species of Hexarthra and Polyarthra (Rotifera, Monogononta): its function in appendage movement and escape behavior. Zool. Anz. 247, 233–248. Hofmann, W., 1982. On the coexistence of two pelagic Filinia species (rotatoria) in lake plußee i. Dynamics of abundance and dispersion. Arch. für Hydrobiol. 95, 125–137. Hovius, J.T., Beisner, B.E., McCann, K.S., Yan, N.D., 2007. Indirect food web effects of Bythotrephes invasion: responses by the rotifer Conochilus in Harp Lake, Canada. Biol. Invasions 9, 233–243. Hudson, C.T., Gosse, P.H., 1886. The Rotifera; or Wheel-animalcules, Both British and Foreign, vols I and II, Longmans, Green, and Co., London. Hyman, L.H., 1951. The Invertebrates: Acanthocephala, Aschelminthes, and Entoprocta. The Pseudocoelomate Bilateria, vol. III, McGrawHill, New York, NY. 572 pp. Ignoffo, T.R., Bollens, S.M., Bochdansky, A.B., 2005. The effect of thin layers on the vertical distribution of the rotifer Brachionus plicatilis. J. Exp. Mar. Biol. Ecol. 316, 167–181. Iltis, A., Riou-Duvat, S., 1971. Variations saisonniéres du peuplement en rotifères des eaux natronées du Kanem (Tchad). Cah. Orstom Hydrobiol. 5, 101–112. Jersabek, C.D., Bolortsetseg, E., Taylor, H.L., 2010. Mongolian rotifers on microscope slides: instructions to permanent specimen mounts from expedition material. Mong. J. Biol. Sci. 8, 51–57. Jersabek, C.D., Segers, H., Dingmann, B.J., 2003. The Frank J. Myers Rotifer collection at the Academy of Natural Sciences: the whole collection in digital images. Acad. Nat. Sci. Phila. Special Publication 20. Jones, B.L., VanLoozen, J., Kim, M.H., Miles, S.J., Dunham, C.M., Williams, L.D., Snell, T.W., 2013. Stress granules from in Brachionus majavacas (Rotifera) in response to a variety of stressors. Comp. Biochem. Physiology, Part A 116, 375–384. King, C.E., 1983. A re-examination of the Lansing effect. Hydrobiologia 104, 135–139. Kleinow, W., Röhrig, A., 1995. Enzyme activity measurement on isolated organs of Brachionus plicatilis (Rotifera). Hydrobiologia 313/314, 171–174. Koste, W., Shiel, R.J., 1987. Rotifera from Australian inland waters. II. Epiphanidae and Brachionidae (Rotifera: Monogononta). Invertebr. Taxon. 7, 949–1021. Langley, J.M., Shiel, R.J., Nielsen, D.L., Green, J.D., 2001. Hatching from the sediment egg-bank, or aerial dispersing? – the use of mesocosms in assessing rotifer biodiversity. Hydrobiologia 446/447, 203–211. Lawrence, C., Sanders, E., Henry, E., 2012. Methods for culturing saltwater rotifers (Brachionus plicatilis) for rearing larval zebrafish. Zebrafish 9, 140–146. Lowe, C.D., Kemp, S.J., Bates, A.D., Montagnes, D.J.S., 2005. Evidence that the rotifer Brachionus plicatilis is not an osmoconformer. Mar. Biol. 146, 923–929. Mark Welch, D.B., Ricci, C., Meselson, M., 2009. (Chapter 13) Bdelloid Rotifers: progress in understanding the success of an evolutionary scandal. In: Schön, I., Martens, K., van Dijk, P. (Eds.), Lost sex: The evolutionary biology of parthenogenesis. Springer Science + Business Media B.V, Dordrecht, pp. 259–279. SECTION | III Protozoa to Tardigrada Melone, G., 2001. Rhinoglena frontalis (Rotifera, Monogononta): a scanning electron microscopic study. Hydrobiologia 466/467, 291–296. Melone, G., Ricci, C., 1995. Rotatory apparatus in Bdelloids. Hydrobiologia 313/314, 91–98. Molinero, J.C., Anneville, O., Souissi, S., Balvay, G., Gerdeaux, D., 2005. Anthropogenic and climate forcing on the long-term changes of planktonic rotifers in Lake Geneva, Europe. J. Plankton Res. 28, 287–296. Obertegger, U., Flaim, G., Sommaruga, R., 2008. Multifactorial nature of rotifer water layer preferences in an oligotrophic lake. J. Plankton Res. 30, 633–643. Obertegger, U., Smith, H.A., Flaim, G., Wallace, R.L., 2011. Using the guild ratio to characterize pelagic rotifer communities. Hydrobiologia 662, 157–162. Park, H.G., Lee, K.W., Cho, S.H., Kim, H.S., Jung, M.-M., Kim, H.-S., 2001. High density culture of the freshwater rotifer, Brachionus calyciflorus. Hydrobiologia 446/447, 369–374. Pourriot, R., 1979. Rotiféres du sol. Rev. d’Ecologie Biol. Sol. 16, 279–312. Pourriot, R., Francez, A.-J., 1986. Rotifères. Bull. Mens. la Société linnéenne Lyon 55, 2–37. Pryakhin, A., Tryapitsina, G.A., Deryabina, L.V., Atamanyuk, N.I., Stukalov, P.M., Ivanov, I.A., Kostyuchenko, V.A., Akleyev, A.V., 2012. Status of ecosystems in radioactive waste reservoirs of the Mayak production association in 2009. Health Physcian 101, 61–63. Ramulu, H.G., Raoult, D., Pontarotti, P., 2012. The rhizome of life: what about metazoa? Front. Cell. Infect. Microbiol. 2, 50. Ricci, C., Caprioli, M., 2005. Anhydrobiosis in bdelloid species, populations and individuals. Intergrative Comp. Biol. 45, 759–763. Ricci, C., Caprioli, M., Fontaneto, D., Melone, G., 2007. Volume and morphology changes of a bdelloid rotifer species (Macrotrachela quadricornifera) during anhydrobiosis. J. Morphol. 269, 233–239. Ricci, C., Fontaneto, D., 2003. Mediterranean rotifers: a very inconspicuous taxon. Biogeographia 25, 161–167. Ricci, C., Melone, G., 1998. Dwarf males in monogonont rotifers. Aquat. Ecol. 32, 361–365. Ricci, C., Perletti, F., 2006. Starve and survive: stress tolerance and lifehistory traits of a bdelloid rotifer. Funct. Ecol. 20, 340–346. Ruttner-Kolisko, A., 1974. Planktonic rotifers: biology and taxonomy. Die Binnengewässer Suppl. 26, 1–146. Santos-Medrano, G.E., Rico-Martinez, R., Velásquez-Rojas, C.A., 2001. Swimming speed and Reynolds numbers of eleven freshwater rotifer species. Hydrobiologia 446/447, 35–38. Sarma, S.S.S., Nandini, S., 2007. Small prey size offers immunity to predation: a case study on two species of Asplanchna and three brachionid prey (Rotifera). Hydrobiologia 593, 67–76. Schöll, K., Szövényi, G., 2011. Planktonic rotifer assemblages of the Danube River at Budapest after the red sludge pollution in Hungary. Bull. Environ. Contam. Toxicol. 87, 124–128. Schröder, T., 2003. Precopulatory mate guarding and mating behaviour in the rotifer Epiphanes senta (Monogononta: Rotifera). Proc. Royal Soc. Lond. B 270, 1965–1970. Schröder, T., 2005. Diapause in monogonont rotifers. Hydrobiologia 546, 291–306. Schröder, T., Howard, S., Arroyo, L., Walsh, E.J., 2007. Sexual reproduction and diapause of Hexarthra sp. (Rotifera) in short-lived Chihuahuan Desert ponds. Freshwater Biol. 52, 1033–1042. Seaman, M.T., Gophen, M., Cavari, B.Z., Azoulay, B., 1986. Brachionus calyciflorus Pallas as agent for removal of E. coli in sewage ponds. Hydrobiologia 135, 55–60. Chapter | 13 Phylum Rotifera Segers, H., 2002. The nomenclature of the Rotifera: annotated checklist of valid family and genus-group names. J. Nat. Hist. 36, 631–640. Segers, H., 2007. Annotated checklist of the rotifers (Phylum Rotifera), with notes on nomenclature, taxonomy and distribution. Zootaxa 1564, 1–104. Segers, H., 2008. Global diversity of rotifers (Rotifera) in freshwater. Hydrobiologia 595, 49–59. Sendacz, S., Caleffi, S., Santos-Soares, J., 2006. Zooplankton biomass of reservoirs in different trophic conditions in the state of São Paulo, Brazil. Braz. J. Biol. 66, 337–350. Serra, M., Snell, T.W., 2009. (Chapter 14) Sex loss in monogonont rotifers. In: Schön, I., Martens, K., van Dijk, P. (Eds.), Lost sex: The evolutionary biology of parthenogenesis. Springer Science + Business Media, B.V, Dordrecht, pp. 281–294. Serra, M., Snell, T., Gilbert, J.J., 2005. Delayed mixis in rotifers: an adaptive response to the effects of density dependent sex on population growth. J. Plankton Res. 27, 37–45. Smith, H.A., Burns, A.R., Shearer, T.L., Snell, T.W., 2012. Three heat shock proteins are essential for rotifer thermotolerance. J. Exp. Mar. Biol. Ecol. 413, 1–6. Smith, J.M., Cridge, A.G., Dearden, P.K., 2010. Germ cell specification and ovary structure in the rotifer Brachionus plicatilis. EvoDevo 1, 5. Snell, T.W., 1986. Effects of temperature, salinity and food level on sexual and asexual reproduction in Brachionus plicatilis (Rotifera). Mar. Biol. 92, 157–162. Snell, T.W., 1998. Estimating chronic toxicity using rotifers. Stand. Methods Exam. Water Wastewater 20 (8420), 8–62. 68–65. Snell, T.W., 2011. A review of the molecular mechanisms of monogonont rotifer reproduction. Hydrobiologia 662, 89–97. Snell, T.W., Garman, B.L., 1986. Encounter probabilities between male and female rotifers. J. Exp. Mar. Biol. Ecol. 97, 221–230. Sørensen, M.V., Giribet, G., 2006. A modern approach to rotiferan phylogeny: combining morphological and molecular data. Mol. Phylogenet. Evol. 40, 585–608. Starkweather, P.L., 1987. Rotifera. In: Pandian, T.J., Vernberg, F.J. (Eds.), Animal energetics. Protozoa through insecta, Vol. 1. Academic Press, Orlando, FL, pp. 159–183. Stemberger, R.S., Gilbert, J.J., 1985. Body size, food concentration, and population growth in planktonic rotifers. Ecology 66, 1151–1159. Strojsová, M., Vrba, J., 2007. Rotifer digestive enzymes: direct detection using the ELF technique. Hydrobiologia 593, 159–165. Thorp, J.H., Mantovani, S., 2005. Zooplankton of turbid and hydrologically dynamic prairie rivers. Freshwater Biol. 50, 1474–1491. 271 Verschoor, A.M., Van der Stap, I., Helmsing, N.R., Lu¨rling, M., Van Donk, E., 2004. Inducible colony formation within the Scenedesmaceae: adaptive responses to infochemicals from two different herbivore taxa. J. Phycol. 40, 808–814. Wallace, R.L., 1980. Ecology of sessile rotifers. Hydrobiologia 73, 181–193. Wallace, R.L., Edmondson, W.T., 1986. Mechanism and adaptive significance of substrate selection by a sessile rotifer. Ecology 67, 314–323. Wallace, R.L., 1987. Coloniality in the phylum Rotifera. Hydrobiologia 147, 141–155. Wallace, R.L., Ricci, C., 2002. (Chapter 2) Rotifera. In: Rundle, S.D., Robertson, A.L., Schmid-Araya, J.M. (Eds.), Freshwater Meiofauna: Biology and ecology. Backhuys Publishers, Leiden, pp. 15–44. Wallace, R.L., Smith, H.A., 2009. Rotifera. In: Likens, G.E. (Ed.), Encyclopedia of inland waters. Elsevier, Oxford, pp. 689–703. Wallace, R.L., Snell, T.W., Ricci, C., Nogrady, T., 2006. Rotifera, second ed. Biology, ecology and systematics, vol. 1, Backhuys Publishers, Leiden. 299 pp. Walsh, E.J., Salazar, M., Remirez, J., Moldes, O., Wallace, R.L., 2006. Predation by invertebrate predators on the colonial rotifer Sinantherina socialis. Invertebr. Biol. 125, 325–335. Walsh, E.J., Schröder, T., Wallace, R.L., Ríos Arana, J.V., Rico-Martínez, R., 2008. Rotifers from selected inland saline waters in the Chihuahuan Desert of México. Saline Syst. 4, 7. Walsh, E.J., Zhang, L., 1992. Polyploidy and body size variation in a natural population of the rotifer Euchlanis dilatata. J. Evol. Biol. 5, 345–353. Weithoff, G., 2004. Vertical niche separation of two consumers (Rotatoria) in an extreme habitat. Oecologia 139, 594–603. Weithoff, G., 2007. Dietary restriction in two rotifer species: the effect of the length of food deprivation on life span and reproduction. Oecologia 153, 303–308. Weithoff, G., Wacker, A., 2007. The mode of nutrition of mixotrophic flagellates determines the food quality for their consumers. Funct. Ecol. 21, 1092–1098. Wilson, C.G., Sherman, P.W., 2010. Anciently asexual bdelloid rotifers escape lethal fungal parasites by drying up and blowing away. Science 327, 574–576. Wilson, C.G., Sherman, P.W., 2013. Spatial and temporal escape from fungal parasitism in natural communities of anciently asexual bdelloid rotifers. Proc. Royal Soc. B 280, 20131255. Yoshida, T., Hairston Jr, N.G., Ellner, S.P., 2004. Evolutionary trade-off between defence against grazing and competitive ability in a simple unicellular alga, Chlorella vulgarls. Proc. Royal Soc. B 271, 1947–1953.