Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
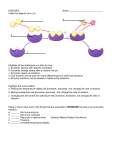
Bringing Together Biomolecular Simulation and Experimental Studies Calculating enzyme kinetic parameters from protein structures Matthias Stein*1 , Razif R. Gabdoulline*† and Rebecca C. Wade* *Molecular and Cellular Modeling Group, EML Research gGmbH, Schloss-Wolfsbrunnenweg 33, 69118 Heidelberg, Germany, and †BIOMS, University of Heidelberg, Im Neuenheimer Feld 368, 69120 Heidelberg, Germany Abstract Enzyme kinetic parameters can differ between different species and isoenzymes for the same catalysed reaction. Computational approaches to calculate enzymatic kinetic parameters from the three-dimensional structures of proteins will be reviewed briefly here. Enzyme kinetic parameters may be derived by modelling and simulating the rate-determining process. An alternative, approximate, but more computationally efficient approach is the comparison of molecular interaction fields for experimentally characterized enzymes and those for which parameters should be determined. A correlation between differences in interaction fields and experimentally determined kinetic parameters can be used to determine parameters for orthologous enzymes from other species. The estimation of enzymatic kinetic parameters is an important step in setting up mathematical models of biochemical pathways in systems biology. Introduction In systems biology, the fluxes and kinetics of substrates, metabolites and products in a biomolecular network are quantified. The network can involve metabolic and/or signalling pathways. The network may be described by a set of coupled deterministic or stochastic differential equations that may be solved by numerical integration. Setting up this set of coupled differential equations requires the input of molecular concentrations or particle numbers and a detailed set of enzymatic kinetic parameters that depend on environmental conditions such as temperature, pH and ionic strength. In enzyme catalysis, enzyme–substrate association and turnover are often described by Michaelis–Menten-like kinetics: k1 k2 E + S ES −→ E + P k−1 (1) where E is the enzyme, S is the substrate and P is the product. Here, kcat or the catalytic activity is given by k2 and K m ≈ k−1 /k1 (when k2 k−1 ) and can be interpreted as a substrate binding affinity. The ratio kcat /K m is a measure of the catalytic efficiency of the enzyme (see Figure 1). When constructing models of biochemical networks, it is often found that a necessary enzymatic kinetic parameter is missing or available only under different experimental conditions or for a different organism from that required. Here, we will review computational approaches to calculate these missing enzymatic kinetic parameters from threedimensional protein structures. Mathematical fitting of Key words: computational approach, enzymatic kinetic parameters, enzyme–substrate association, molecular interaction field, systems biology, three-dimensional structure of protein. Abbreviations used: KIE, kinetic isotope effect; MM, molecular mechanical; QM, quantum mechanical; PIPSA, protein interaction property similarity analysis; qPIPSA, quantitative PIPSA. 1 To whom correspondence should be addressed (email [email protected]). Biochem. Soc. Trans. (2008) 36, 51–54; doi:10.1042/BST0360051 experimental data or parameter estimation based on network models will not be considered here. Different steps of enzymatic catalysis need different modelling approaches Enzymes lower the transition state barrier and thus accelerate the reaction compared with the uncatalysed reaction in solution. The formation of an initial encounter complex is the first step in enzyme catalysis (affecting k1 and thus K m ) and can be investigated by modelling the enzyme–substrate association process. Enzymatic turnover may be modelled for the substrate bound in the active site of the enzyme. Recent progress has delivered insights into the degree of transition state stabilization by enzymes, e.g. the preferential binding of the transition state by a preformed enzyme pocket. The appropriate choice of computational tool to calculate enzyme kinetic parameters depends critically on the time scale of the slowest and thus rate-limiting step. Depending on the underlying chemistry, very fast processes, such as proton transfer (10−12 s) or side-chain reorientation (10−11 –10−10 s), may be rate-limiting. On the other hand, slower processes such as hinge bending at domain interfaces (10−11 –10−7 s) or other conformational changes may be critical [1] (for a review of simulation techniques, see [2]). Modelling the transition state When chemical bond breaking and formation processes are rate-determining, accurate modelling of the free energy differences between the enzyme–substrate complex and the transition state [E–S]# can yield the barrier to catalysis and thus kcat (see Figure 1). Modern transition state theory C The C 2008 Biochemical Society Authors Journal compilation 51 52 Biochemical Society Transactions (2008) Volume 36, part 1 Figure 1 Schematic diagram of the factors determining the rate of an enzymatic reaction The enzyme (E) and the substrate (S) form an enzyme–substrate complex (E–S). Upon converting the substrate, the enzyme– substrate complex passes over a transition state ([E–S]# ) to yield the product (P) plus the recovered enzyme. Sufficient conformational sampling around the transition state is required to achieve a statistically significant determination of the transition-state free energy changes. In transition-state theory, the changes in free energy [G# (T)] can be used to determine the kinetics [k(T)] of over-the-barrier motion. For light nuclei, QM corrections to transition state theory [among others, the insertion of a transmission coefficient γ (T) to account for through-the-barrier tunnelling motion of protons, hydrogen radicals and hydrides] may explain the KIEs of some reaction rates. T is temperature, β = k B T, and k B is Boltzmann’s constant. of enzymatic reactions can be found in [3]. Once the transition state has been localized, sufficient sampling of the configurational space around the transition state is critical for obtaining kinetic parameters. Usually, a VB (valence bond) description for educts, products and intermediates along a simplified reaction co-ordinate is used. Along this reaction co-ordinate, VTST (variational transition state theory) can be employed to calculate the free energies of activation and thus to calculate rate constants. For a review of possible potential energy functions to model the transition state, see [4]. QM (quantum mechanical) tunnelling effects become important when light nuclei (protons/hydrides) are involved. Then the reaction displays a KIE (kinetic isotope effect) and the reaction rate is lower when deuterated substrates or solvents are used. Through-barrier tunnelling rather than over-the-barrier motion (see Figure 1) can be dominating. Modelling of the rate-determining proton transfer steps requires a QM treatment of the bond breaking/forming process plus consideration of the effect of the surrounding protein atoms on the barrier height by hybrid QM/MM (molecular mechanical) models. An alternative to the static QM treatment of the active site is the consideration of Newtonian atomic motion plus explicit integration of the time-dependent Schrödinger equation in a C The C 2008 Biochemical Society Authors Journal compilation QCMD (quantum-classical Molecular Dynamics) approach and, recently, the extension to electronic motion in a hybrid Car–Parrinello QM/MM approach (see [5]). If sufficient sampling is carried out, accurate free energy barriers in enzymatic catalysis can be obtained. A comprehensive review on QM methods to calculate enzymatic rate constants can be found in [6]. Modelling diffusional enzyme–substrate association The enzyme–substrate association step can be responsible for a large modulation of enzymatic activity. When the enzyme–substrate interaction is strong and catalytic turnover is fast, the transition state barrier for the enzymatic substrate-to-product conversion is not rate-determining and kcat /K m is close to the diffusion limit [7]. In this case, the formation of an initial enzyme–substrate diffusional association complex becomes limiting. Indications for this are the dependence of the rate constants on ionic strength or viscosity of the solvent. Brownian dynamics simulation can be used to simulate the diffusional association of substrate and enzyme [8]. In these simulations, the solvent is considered implicitly by solving the Poisson–Boltzmann equation to model electrostatic interactions and its dynamic effects are modelled by stochastic terms. Examples of Brownian dynamic simulation that yield quantitative results in agreement with experimental findings include enzyme– substrate interactions (e.g. acetylcholinesterase, superoxide dismutase and triose phosphate isomerase), enzyme– inhibitor binding (e.g. barnase–barstar), antibody–antigen binding and the association of electron transfer proteins [9]. Molecular interaction fields Electrostatic potential has been recognized as one of the major determinants of enzymatic catalysis both in enzyme– substrate association and in transition state stabilization. The comparative analysis of the electrostatic potentials of a large number of enzymes by qPIPSA [quantitative PIPSA (protein interaction property analysis)] [10] allows differences in the molecular interaction field to be related directly to differences in enzymatic kinetic parameters between species or between mutants and wild-type (see Figure 2). For deriving such a correlation, a reference set of experimentally well-characterized enzymes is required. It has been shown that results from PIPSA are comparable with those from Brownian dynamics simulation but are obtained at a fraction of the computing time [11]. From qPIPSA, estimates of enzymatic K m and kcat /K m values can be made based on a comparison of the electrostatic potentials of enzymes around the active site or any region responsible for specific enzyme kinetics. The computational efficiency allows the functional and kinetic characterization of a large number of enzymes, the detection of experimental outliers and the characterization of the enzymes of an entire metabolic pathway [10,12,13]. Bringing Together Biomolecular Simulation and Experimental Studies Figure 2 Scheme showing a comparative approach for estimating enzymatic kinetic parameters based on an analysis of molecular interaction fields The molecular interaction fields of enzymes that catalyse the same reaction and possess similar three-dimensional structures are calculated and compared. These molecular interaction fields can be, for example, the electrostatic potential or a hydrophobic field. Differences in molecular interaction fields may be correlated with differences in enzymatic kinetic parameters, such as K m and k cat /K m , for a given set of experimentally well-characterized enzymes. Such a correlation can be used to determine enzyme kinetic constants for enzymes from other species or enzyme mutants [10]. Conclusions References Molecular simulations play an important role in elucidating enzymatic reaction mechanisms. Progress in computing algorithms and hardware has enabled the calculation of enzymatic kinetic parameters at various levels of theory. The direct applicability of simulated kinetic parameters in systems biology is still hampered by the required high accuracy of computed kinetic parameters. One example can be found in [14]. In systems biology, the focus shifts from investigating single enzymes to protein families, enzyme classes or entire pathways. Thus computationally efficient simulations are necessary to deal with a large number of biomolecules. Coarse-graining and comparative approaches are general strategies to accomplish this task [15]. The availability of more and more accurate experimental data and computational approaches will bring molecular and biochemical network simulations closer together, thus enabling insights into more complex biological phenomena. 1 Benkovic, S.J. and Hammes-Schiffer, S. (2003) A perspective on enzyme catalysis. Science 301, 1196–1202 2 Karplus, M. and McCammon, J.A. (2002) Molecular dynamics simulations of biomolecules. Nat. Struct. Biol. 9, 646–652 3 Garcia-Viloca, M., Gao, J., Karplus, M. and Truhlar, D.G. (2004) How enzymes work: analysis by modern reaction rate theory and computer simulations. Science 303, 186–195 4 Gao, J., Ma, S., Major, D.T., Nam, K., Pu, J. and Truhlar, D.G. (2006) Mechanisms and free energies of enzymatic reactions. Chem. Rev. 106, 3188–3209 5 dal Peraro, M., Ruggerone, P., Raugei, S., Gervasio, F.L. and Carloni, P. (2007) Investigating biological systems using first principles Car–Parrinello molecular dynamics simulations. Curr. Opin. Struct. Biol. 17, 149–156 6 Gao, J.L. and Truhlar, D.G. (2002) Quantum mechanical methods for enzyme kinetics. Annu. Rev. Phys. Chem. 53, 467–505 7 Stroppolo, M.E., Falconi, M., Caccuri, A.M. and Desideri, A. (2001) Superefficient enzymes. Cell. Mol. Life Sci. 58, 1451–1460 8 Gabdoulline, R.R. and Wade, R.C. (2002) Biomolecular diffusional association. Curr. Opin. Struct. Biol. 12, 204–213 9 Gabdoulline, R.R. and Wade, R.C. (2001) Protein–protein association: investigation of factors influencing association rates by Brownian dynamics simulations. J. Mol. Biol. 306, 1139–1155 10 Gabdoulline, R.R., Stein, M. and Wade, R.C. (2007) qPIPSA: relating enzymatic kinetic parameters and interaction fields. BMC Bioinformatics 8, 373 11 de Rienzo, F., Gabdoulline, R.R., Menziani, M.C., de Benedetti, P.G. and Wade, R.C. (2001) Electrostatic and Brownian dynamics simulation analysis of plastocyanin and cytochrome f. Biophys. J. 81, 3090–3104 12 Stein, M., Gabdoulline, R.R. and Wade, R.C. (2006) Integrating structural and kinetic enzymatic information in systems biology. in NIC Workshop 2006: From Computational Biophysics to Systems Biology, NIC Series, Vol. 34 (Hansmann, U.H.E., Meinke, J., Mohanty, S. and Zimmermann, O., eds), pp. 129–131, John von Neumann Institute for Computing, Jülich Financial support from the German Federal Ministry for Research (BMBF) ‘HepatoSys’ project (grant numbers 0313076 and 0313078C), the Klaus Tschira Foundation and the Center for Modelling and Simulation in the Biosciences (BIOMS; Heidelberg, Germany) is gratefully acknowledged. C The C 2008 Biochemical Society Authors Journal compilation 53 54 Biochemical Society Transactions (2008) Volume 36, part 1 13 Stein, M., Gabdoulline, R.R. and Wade, R.C. (2007) The estimation of kinetic parameters in systems biology by comparing molecular interaction fields. in Experimental Standard Conditions of Enzyme Characterizations; Proceedings of the 2nd International Beilstein Workshop on ESCEC (Hicks, M.G. and Kettner, C., eds), pp. 237–253, Logos Verlag, Berlin 14 Gabdoulline, R.R., Kummer, U., Olsen, L.F. and Wade, R.C. (2003) Concerted simulations reveal how peroxidase compound III formation results in cellular oscillations. Biophys. J. 85, 1421–1428 C The C 2008 Biochemical Society Authors Journal compilation 15 Stein, M., Gabdoulline, R.R. and Wade, R.C. (2007) Bridging from molecular simulation to biochemical networks. Curr. Opin. Struct. Biol. 17, 166–172 Received 14 September 2007 doi:10.1042/BST0360051