Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
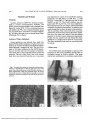
Investigative Ophthalmology & Visual Science, Vol. 33, No. 2, February 1992 Copyright © Association for Research in Vision and Ophthalmology The Influence of Cyclic AMP Upon Na,K-ATPase Activity in Rabbit Ciliary Epithelium N. A. Delamere and K. L. King ATPase activity was measured in samples of freshly dissected rabbit ciliary epithelium. The epithelium was ruptured in distilled water, frozen briefly, and incubated at 37°C in a buffer containing 100 mM NaCl and 32P-labeled adenosine triphosphate (ATP). The rate of ATP hydrolysis by the epithelium was linear for as long as 45 min. Ouabain (1 mM) reduced the ATP hydrolysis rate by approximately 50%. When the epithelium was preincubated for 10 min. in the presence of 1 mM dibutyryl cyclic adenosine monophosphate (cAMP), the ouabain-sensitive (Na,K-ATPase) activity was diminished; ouabain-insensitive ATPase activity was not reduced. Preincubation of the epithelium with forskolin with isobutylmethylxanthine also reduced ouabain-sensitive ATPase activity. These observations suggest that the ciliary epithelium may have a mechanism for short-term modulation of Na,K-ATPase activity by cAMP. Such a mechanism could be linked to the ability of cAMP-dependent protein kinase to reduce Na,K-ATPase activity in the tissue. Invest Ophthalmol Vis Sci 33:430-435,1992 Mittag et al6 have obtained direct evidence that the rabbit ciliary epithelium has adenylate cyclase that is vasoactive intestinal peptide and beta-adrenergically responsive. These findings suggest that cAMP might have a role in the transduction of signals within the epithelium. In a recent study, we isolated membrane material from rabbit ciliary processes and observed that Na,K-ATPase activity in the preparation was diminished in the presence of cAMP-dependent protein kinase, protein kinase A.7 This finding suggested that when cytoplasmic cAMP activates protein kinase A, Na,K-ATPase activity could be lowered, perhaps signifying that cAMP is involved in short-term regulation of Na,K-ATPase activity in rabbit ciliary processes. A role for cAMP in short-term regulation of Na,K-ATPase activity in kidney proximal tubule segments has been proposed by Bertorello and Aperia.8 In the present study, we have examined changes of Na,K-ATPase activity that occur when the ciliary epithelium is pretreated with dibutryl cAMP or forskolin. To do this, we isolated pure samples of rabbit ciliary epithelium and established a protocol for measuring Na,K-ATPase activity in permeabilized cells. This approach differs from the conventional methodology of determining Na,K-ATPase activity in a purified membrane preparation made by a lengthy differential centrifugation procedure, during which loosely bound membrane-associated components may be lost from the epithelial membranes. Using the permeabilized cell technique, ATPase activity could be measured immediately after the epithelium was removed from the incubation solution. The ciliary processes are covered by a doublelayered epithelium, the ciliary epithelium, which is recognized as the site of aqueous humor formation.1 The ciliary epithelium has a high Na,K-ATPase activity, and morphological studies have shown that the Na,K-ATPase is most densely localized at the basolateral membranes of the nonpigmented epithelium, the epithelial layer that faces the aqueous humor.2 While the precise contribution of active sodium transport to aqueous secretion has yet to be elucidated, it is widely believed that active sodium transport is linked to the mechanism of fluid formation by the epithelium.1 In fact, experiments by Cole,3 Becker,4 and Bonting and Becker5 have shown that aqueous humor formation in rabbits can be inhibited by ouabain, the specific inhibitor of Na,K-ATPase. It has been known for some time that intraocular pressure can be lowered by a number of agents that alter the level of cyclic adenosine monophosphate (cAMP) in tissues at the anterior of the eye (Davson1). From the Department of Ophthalmology and Visual Sciences, Kentucky Lions Eye Research Institute, University of Louisville School of Medicine, Louisville, Kentucky. Supported by USPHS Research Grant No. EY06915, the Kentucky Lions Eye Foundation, and an unrestricted grant from Research to Prevent Blindness, Inc., New York, New York. Submitted for publication: March 12, 1991; accepted September 19, 1991. Reprint requests: Nicholas A. Delamere, Department of Ophthalmology and Visual Sciences, University of Louisville, Louisville, KY 40292. 430 Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933390/ on 08/03/2017 No. 2 NA,K-ATPA5E ACTIVITY IN CILIARY EPITHELIUM / Delomere ond King Materials and Methods Chemicals Ouabain, dibutyryl cAMP (N6,2'-O-dibutyryl adenosine 3', 5'-cyclic monophosphate), forskolin, and IBMX (isobutylmethylxanthine) were obtained from Sigma (St. Louis, MO). ATP as a triethylammonium salt labeled with 32P at the terminal phosphate group was purchased from Amersham (Arlington Heights, IL). All other chemicals were purchased from Fisher (Pittsburgh, PA) or Sigma. Isolation of Ciliary Epithelium Ciliary epithelium was obtained from adult New Zealand White rabbits weighing 2-3 kg that were sacrificed by an overdose of sodium pentobarbital administered through a marginal ear vein. The use of animals in this study conformed to the ARVO Resolution on the Care and Use of Animals in Research. After death, the eye was proptosed and the cornea was removed. The iris, ciliary body, lens, and vitreous body then were removed from the eye in one piece 431 and transferred to a dish of ice-cold Kreb's solution containing 119 raM NaCl, 4.7 mM KC1, 1.1 mM KH2PO4> 1 mM MgCl2; 5.5 mM glucose, and 25 mM NaHCO3 at pH 7.4. The Kreb's solution was pregassed with 95% 0^5% CO2. The ciliary epithelium was dissected from the eye using a method described in detail by Jumblatt and coworkers.9 A radial cut was made through the iris from the pupil to the periphery. The iris and base of the ciliary body then was gently peeled away from the lens, leaving the ciliary epithelium from the ridges and valleys of the pars plicata attached to the lens zonules (suspensory ligaments). Finally, the lens zonules were cut, freeing the ciliary epithelium as a single strip of tissue 25-35 mm long. The ciliary epithelium from each eye was divided into five pieces. ATPase Assay The ATPase assay was developed to measure ATP hydrolysis by the ciliary epithelium shortly after an incubation period in Kreb's solution. The assay method is a modification of a technique used by Katz Fig. 1. The rabbit ciliary epithelium immersed in Krebs solution. Viewed at a magnification of X100 (A), ridges can be seen at the points where the epithelium had covered the ciliary processes. At higher magnification (X400) using Nomarski optics, the nonpigmented cells are shown in freshly dissected tissue (B). A low-power transmission electron micrograph of the tissue (C) shows the two cell layers. Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933390/ on 08/03/2017 432 INVESTIGATIVE OPHTHALMOLOGY b VISUAL SCIENCE / February 1992 and coworkers.10 It differs from conventional ATPase assays because ATPase activity is measured in permeabilized cells rather than in membrane material that is first isolated from the tissue and purified by several stages of washing and centrifugation. After a specified period of incubation in Kreb's solution, the ciliary epithelium was removed and permeabilized in a two-stage process. First, each piece of epithelium was placed in a tube containing 184 n\ distilled water for 5 min so the cells could swell and rupture. Permeabilization was ensured by a brief freeze-thaw in liquid nitrogen before 166 /x\ of a buffer solution was added to give a final composition of 40 mM histidine HC1, 10 mM KC1, 3 mM mg MgCl2, 100 mM NaCl and 0.8 mM Tris-EGTA and 1 mM 32 P-labelled ATP at pH 7.4. The buffer solution also contained 11 /ig/ml alamethicin, which was added to ensure a high cation permeability of any resealed membrane compartments and to counter the possibility that Na,K-ATPase activity would be influenced by local cation gradients.11 In specified experiments, the solutions contained 1 mM ouabain. This buffer composition is the same as that used earlier for experiments with partially purified ciliary process membrane preparations.7 The assay mixture was incubated at 37°C for a specified period, and the reaction was terminated by the addition of 100 ^1 ice cold 0.5 NaOH using an established methodology.12 ATP hydrolysis in each tube was quantified by determining the amount of radiolabelled inorganic phosphate that had been released. The amount of tissue protein in each tube was determined by a Bio Rad assay. ATPase activities were calculated as nanomoles phosphate released//tg protein/ h. A paired Student's t test was used to determine the statistical significance of differences between ATPase activities measured in treated and control tissues. Vol. 33 layer. The epithelial layers remain attached to the lens zonules when the iris and ciliary body are stripped away. The epithelium is removed as a narrow sheet that is wrinkled at the points where it used to fold over the surface of the ciliary processes (Fig. 1). ATPase Activity ATPase activity was measured in freshly dissected ciliary epithelium following a brief permeabilization procedure. The rate of ATP hydrolysis was linear over a period of 45 min. (Fig. 2). The control rate of ATP hydrolysis was 4.1 ±0.6 nmol phosphate released//ig protein/h (mean ± SEM). In the presence of 1 mM ouabain, the rate was 2.0 ± 0.2 nmol phosphate released//^ protein/h, which is significantly different (P = .001) from the control value. Influence of Dibutyryl cAMP Upon ATPase Activity Dibutyryl cAMP was used because it is more permeable than cAMP. Freshly dissected ciliary epithelium was maintained for 10 min at 37°C in Kreb's solution containing 1 mM dbcAMP. Control pieces of ciliary epithelium were incubated in the absence of dbcAMP. After the incubation period, the pieces of epithelium were removed from the Kreb's solution immediately prior to permeabilization and measurement of ATP hydrolysis. Thus, dbcAMP was not present during the ATP hydrolysis portion of the experiment. In epithelium pretreated with dbcAMP, the ouabain-sensitive portion of ATP hydrolysis was reduced. CONTROL Results Isolation of Ciliary Epithelium In the past, most ATPase measurements and related transport studies were conducted with segments of iris-ciliary body or with ciliary processes as a source of tissue.712 These tissues contain a number of different cell types. In the present study, we have used a technique in which pure ciliary epithelium is isolated from the ridges and valleys of the pars plicata. The dissection procedure, described in detail by Jumblatt and coworkers9, relies on the close attachment of the lens zonules with the basement membrane of the nonpigmented (aqueous humor facing) ciliary epithelium OUABAIN 60 TIME (min) Fig. 2. Hydrolysis of ATP by permeabilized ciliary epithelium in the presence (closed circles) or absence (open circles) of ouabain. Each point represents the mean of data from 12 tissue pieces with the standard error indicated by a vertical line. The lines were fitted to the data by linear regression. Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933390/ on 08/03/2017 NA,K-ATPASE ACTIVITY IN CILIARY EPITHELIUM / Delomere and King No. 2 A typical experiment is shown in Figure 3a. In eight experiments, pretreatment of the epithelium with 1 mM dbcAMP reduced the total ATP hydrolysis rate by slightly more than 50% (P < 0.01), while the ouabain-insensitive ATPase component was not significantly altered (Fig. 3b). No significant alteration of ATPase activity was observed when the epithelium was pretreated with 10~4 M or 10"5 M dbcAMP. We also examined the influence of forskolin on ATPase activity because concentrations of 10"5 and 10~4 M forskolin activate adenylate cyclase in rabbit ciliary processes.13 ATPase activity was measured following a 10 min period of exposure of the ciliary epithelium to forskolin (5 X 10~5 M) in Kreb's solution that also contained the phosphodiesterase inhibitor IBMX (10"4 M). Forskolin/IBMX caused a reduction of total ATPase activity, but ouabain-insensitive ATPase activity was not affected (Fig. 4). Control epithelium dbcAMP-treated epithelium 433 C Z l - OUABAIN ^ + OUABAIN 120 100 ( .b 80 < 8 60 O c r-i 40 20 • CONTROL 10 min PRETREATMENT FORSKOLIN/IBMX (5X 105M/10"\i) Fig. 4. The influence of forskolin + isobutylmethylxanthine (IBMX) upon ATPase activity measured in ciliary epithelium. Tissues were preincubated for 10 min in control Krebs solution or Kreb's containing forskolin (5 X 10"5 M) together with IBMX (10"" M). ATPase activity was determined in the absence (open bars) or presence (shaded bars) of ouabain. The data are the mean of eight experiments with the standard error indicated by the vertical bar. Total ATPase activity ( - ouabain) was significantly lower (P < 0.01) in forskolin/IBMX-treated ciliary epithelium. Ouabain-insensitive ATPase activity (+ ouabain) was not significantly altered by forskolin/IBMX pretreatment. Discussion 0 10 20 30 40 SO 60 70 TIME (min) 0 10 20 30 40 SO 60 70 TIME (min) C H - OUABAIN + OUABAIN CONTROL 10 min PRETREATMENT WITH 1 mM dbcAMP Fig. 3. The influence of dibutryl cAMP upon ATP hydrolysis. (A) A typical experiment showing the ATP hydrolysis by pieces of ciliary epithelium preincubated for 10 min in control Krebs solution (left panel) or Krebs solution containing 1 mM dbcAMP (right panel). Each point shows the ATP hydrolysis measured from a single tissue piece. (B) The mean rates of ATP hydrolysis measured in control and dbcAMP-treated ciliary epithelium in the presence (shaded bar) or absence (open bar) of ouabain. The data of the mean of eight experiments with the standard error indicated by the vertical bar. The present experiments demonstrate that when rabbit ciliary epithelium from the pars plicata is pretreated with dibutyryl cAMP, ouabain-sensitive ATPase activity is reduced. Ouabain-insensitive ATPase activity is not diminished by dbcAMP pretreatment. Forskolin, an activator of adenylate cyclase, elicits similar results. While cAMP might alter many aspects of cellular function in the ciliary epithelium, our finding that dbcAMP or forskolin pretreatment reduces Na,K-ATPase activity adds support to Mittag's proposal6 that the adenylate cyclase system in ciliary epithelial cells might participate in the regulation of the membrane transport machinery involved in aqueous humor secretion. Because cAMP also may cause parallel increases or decreases in the activity of other membrane transport mechanisms in the ciliary epithelium, we are unable to specify what net changes of aqueous humor formation would result following cAMP-dependent reduction of Na,K-ATPase activity. However, there have been reports of reduced aqueous humor flow in forskolin-treated rabbits and humans.13 The design of these studies resulted in ATPase activity being measured shortly after removal of the epithelium from Kreb's solution that contained dbcAMP or forskolin. Thus, the modification of the ATP hydrolysis pattern, which was triggered by dbcAMP or forskolin, did not require the continued presence of these Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933390/ on 08/03/2017 434 INVESTIGATIVE OPHTHALMOLOGY & VISUAL SCIENCE / February 1992 agents. One possible explanation of these finding is that cAMP alters epithelial membrane permeability, and this change somehow alters the access of sodium or potassium to the Na,K-ATPase. However, this interpretation seems unlikely because the ATP hydrolysis measurements were carried out in the presence of alamethicin, which establishes high membrane cation permeability.11 Therefore, it seems possible that the diminished ouabain-sensitive ATPase activity in dbcAMP-treated tissue may signify a cAMPdependent mechanism for short-term reduction of Na,K-ATPase activity in the rabbit ciliary epithelium. The existence of a mechanism for short-term regulation of Na,K-ATPase activity in rat kidney proximal convoluted tubules (PCT) has been proposed by Bertorello and Aperia.8 Inhibition of PCT Na,K-ATPase by dopamine14 and the demonstration that GTP binding proteins and protein kinase C may be involved1516 have led Bertorello and Aperia to propose a complex model for the Na,K-ATPase regulatory system8. Because PCT Na,K-ATPase activity is reduced by specific DA 1 (dopaminergic) agonists17 and the effect can be potentiated by dbcAMP or forskolin,18 activation of adenylate cyclase and an increase in cellular cAMP were suggested to be an integral part of the Na,K-ATPase regulatory process. A similar pattern of altered Na,K-ATPase activity was found recently in guinea pig striatal neurons that had been exposed to dopamine, specific D, and D2 dopamine receptor agonists, and forskolin.19 Interestingly, cAMP has been reported to stimulate the active transport of sodium and potassium in frog retinal pigment epithelium2021 and dogfish rectal gland.22 In many cases, cellular responses to cAMP are mediated by cAMP-dependent protein kinase23. This kinase, but not cAMP, reduces the Na,K-ATPase activity measured in a membrane preparation purified from rabbit ciliary processes.7 In the present experiments with intact rabbit ciliary epithelium, cAMP-dependent protein kinase activation may be involved in the mechanism that translates dbcAMP or forskolin exposure into a change of Na,K-ATPase activity. Experiments by Lingham and Sen24 and by Meister et al17 support the notion that cAMP-dependent protein kinase might be a part of a mechanism that is able to modulate Na,K-ATPase activity. However, how activation of cAMP-dependent protein kinase could result in a change of Na,K-ATPase activity has yet to be determined. While Yeh and coworkers25 have suggested that Na,K-ATPase can be phosphorylated by a membrane-bound protein kinase, and Lowndes et al26 have shown that protein kinase C can induce phosphorylation of Na,K-ATPase, there is no convincing evidence that phosphorylation of the Vol. 33 Na,K-ATPase subunits by cAMP-dependent protein kinase takes place or that such phosphorylation would alter the activity of the enzyme as an ion transporter. Phosphorylation of an intermediate protein may be required to bring about a change in Na,K-ATPase activity. It has been suggested recently that DARPP-32, a cAMP-regulated phosphoprotein found in kidney27 and ciliary epithelium.28 may be involved, possibly as a third messenger, in Na,K-ATPase modulation.19 Key words: ciliary epithelium, cAMP, Na,K-ATPase References 1. Davson H: Aqueous humor and the intraocular pressure. In Physiology of the Eye, Davson H, editor. New York, Pergamon Press, 1990, pp. 3-95. 2. Usukura J, Fain GL, and Bok D: 3H ouabain localization of Na,K-ATPase in the epithelium of the rabbit ciliary body pars plicata. Invest Ophthalmol Vis Sci 29:606, 1988. 3. Cole D: Secretion of the aqueous humour. Exp Eye Res 25:161, 1977. 4. Becker B: Ouabain and aqueous humor dynamics in the rabbit eye. Invest Ophthalmol 2:325, 1963. 5. Bonting SL and Becker B: Inhibition of enzyme activity and aqueous humor flow in the rabbit eye after intravitreal injection of ouabain. Invest Ophthalmol 3:523, 1964. 6. Mittag TW, Tormay A, and Podos SM: Vasoactive intestinal peptide and intraocular pressure: Adenylate cyclase activation and binding sites for vasoactive intestinal peptide in membranes of ocular ciliary processes. J Pharmacol Exp Ther 241:230, 1986. 7. Delamere NA, Socci RR, and King KL: Alteration of sodium, potassium-adenosine triphosphatase activity in rabbit ciliary processes by cyclic adenosine monophosphate-dependent protein kinase. Invest Ophthalmol Vis Sci 31:2164, 1990. 8. Bertorello A and Aperia A: Short-term regulation of Na+,K+-ATPase activity by dopamine. American Journal of Hypertension 3:5IS, 1990. 9. Jumblatt MM, Raphael B, and Jumblatt JE: A simple method for the isolation of ciliary epithelium. Exp Eye Res 52:229, 1991. 10. Katz Al, Doucet A, and Morel F: Na-K-ATPase activity along the rabbit, rat, and mouse nephron. Am J Physiol 237:F114, 1979. 11. Xie Z, Wang Y, Ganjeizadeh M, McGee R, and Askari A: Determination of total (Na+-K+)-ATPase activity of isolated or cultured cells. Anal Biochem 183:215, 1989. 12. Socci RR and Delamere NA: The effect of vanadate upon calcium-stimulated ATPase of the rabbit iris-ciliary body. Invest Ophthalmol Vis Sci 29:1866, 1988. 13. Caprioli J, Sears M, Bausher L, Gregory D, and Mead A: Forskolin lowers intraocular pressure by reducing aqueous inflow. Invest Ophthalmol Vis Sci 25:268, 1984. 14. Aperia A, Bertorello A, and Seri I: Dopamine causes inhibition of Na+-K+-ATPase activity in rat proximal convoluted tubule segments. Am J Physiol 252:F39, 1987. 15. Bertorello A and Aperia A: Na+-K+-ATPase is an effector protein for protein kinase C in renal proximal tubule cells. Am J Physiol 256:F370, 1989. Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933390/ on 08/03/2017 No. 2 NA K-ATPASE ACTIVITY IN CILIARY EPITHELIUM / Delomere and King 16. Bertorello A and Aperia A: Regulation of Na+,K+-ATPase activity in kidney proximal tubules: Involvement of GTP binding proteins. Am J Physiol 256:F57, 1989. 17. Meister B, Fryckstedt J, Schalling M, Cortes R, Hokfelt T, Aperia A, Hemmings HC, Nairn AC, Ehrlich M, and Greengard P: Dopamine- and cAMP-regulated phosphoprotein (DARPP-32) and dopamine DA, agonist-sensitive Na+,K+-ATPase in renal tubule cells. Proc Natl Acad Sci USA 86:8068, 1989. 18. Bertorello A and Aperia A: Permissive role of cAMP for dopamine-2 inhibition of Na+,K+-ATPase activity in renal proximal tubule cells. Proceedings of the International Union of Physiological Sciences, Helsinki, Finland, July 1989, p. 368. 19. Bertorello A, Hopfield J, Aperia A, and Greengard P: Inhibition by dopamine of (Na+ + K+)ATPase activity in neostriatal neurons through D, and D2 dopamine receptor synergism. Nature 347:386, 1990. 20. Miller S and Farber D: Cyclic AMP modulation of ion transport across frog retinal pigment epithelium. J Gen Physiol 83:853, 1984. 21. Hughes BA, Miller SS, and Machen TE: Effects of cyclic AMP on fluid absorption and ion transport across frog retinal pigment epithelium. J Gen Physiol 83:875, 1984. 435 22. Silva P, Stoff J, and Epstein FH: Indirect evidence for enhancement of Na-K-ATPase activity with stimulation of rectal gland secretion. Am J Physiol 237:F468, 1979. 23. Berridge MJ: The molecular basis of communication within the cell. Sci Am 253:142, 1985. 24. Lingham RB and Sen AK: Regulation of rat brain (Na+,K+), ATPase activity by cyclic AMP. Biochim Biophys Acta 688:475, 1982. 25. Yeh L, Ling L, English L, and Cantley L: Phosphorylation of the (Na,K)-ATPase by a plasma membrane-bound protein kinase in friend erythroleukemia cells. J Biol Chem 258:6567, 1983. 26. Lowndes JM, Hokin-Neaverson M, and Bertics PJ: Kinetics of phosphorylation of Na+/K+-ATPase by protein kinase C. Biochim Biophys Acta 1052:143, 1990. 27. Aperia A, Hokfelt T, Meister B, Bertorello A, Fryckstedt J, Holtback U, and Seri I: The significance of L-amino acid decarboxylase and DARPP-32 in the kidney. American Journal of Hypertension 3:1 IS, 1990. 28. Stone RA, Laties M, Hemmings HC, Ouimet CC, and Greengard P: DARPP-32 in the ciliary epithelium of the eye: A neurotransmitter-regulated phosphoprotein of brain localizes to secretory cells. J Histochem Cytochem 34:1465, 1986. Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933390/ on 08/03/2017