Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
SNARE (protein) wikipedia , lookup
Multi-state modeling of biomolecules wikipedia , lookup
Endomembrane system wikipedia , lookup
Cell membrane wikipedia , lookup
Lipid bilayer wikipedia , lookup
G protein–coupled receptor wikipedia , lookup
Theories of general anaesthetic action wikipedia , lookup
Signal transduction wikipedia , lookup
Implicit solvation wikipedia , lookup
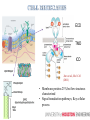
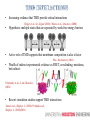
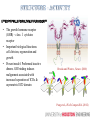
MULTISCALE MODELING OF THE ORGANIZATIONS OF RECEPTOR TRANSMEMBRANE DOMAINS IN LIPID MEMBRANES Wei Dai, Manolis Doxastakis Dept. of Chemical & Biomolecular Engineering University of Houston ECD TMD ICD Jura et al., Mol. Cell (2011) • Membrane proteins 25% but few structures characterized • Signal transduction pathways; Key cellular process • Increasing evidence that TMD provide critical interactions Finger et al., Sci. Signal (2010); Moore et al., Structure, (2008) • Hypothesis: multiple states that are separated by weak free energy barriers • Active role of TMD supports that membrane composition is also a factor Pike., Biochemistry (2002) • Wealth of indirect experimental evidence as FRET, crosslinking, mutations, but indirect. Polyansky et al., J.Am.Chem.Soc. (2012) • Recent simulation studies support TMD interactions Janosi et al., Biophys. J. (2010); Prakash et al., Biophys. J. (2010)(2011) • Amino acid sequence – membrane composition • Spatial and temporal limitations rotations at (O)(10-6) -- (O)(10-4) s Rigby et al., Biochemistry (1996); Sharpe et al., Biochemistry (2002) de Meyer et al., Biophys. J. (2010) Glycophorin A (GPA) • Mesoscopic protein models in complex environments • Implicit membrane models • Coarse-Grained models with a level of amino-acid specificity Marrink et al., J. Phys. Chem. (2007) MacKenzie et al., Science (1997) • Parallel Monte Carlo simulations with hundreds of unconstraint pairs algorithm Janosi et al., J. Chem. Phys. (2009); Janosi et al., Biophys. J. (2010) • GpA in single component bilayer Free energy of dimerization -7 to -9 kcal/mol Lipid induced interactions significant at short separations Prediction of TMD dimer in excellent agreement to experimental data • ErbB family TMDs • ErbB2 in DPPC + Cholesterol Cholesterol alters association pathway Prakash et al., Biophys. J. (2010) (2011) DPPC • Cholesterol promotes dimerization Anbazhagan and Schneider, Bioch. Biophys. Acta, (2010) • Decreased association affinity in mammalian membranes (~30% cholesterol) , -3.9 kcal/mol Single component +30% chol Chen et al., J. Am. Chem. Soc., (2010) • Dramatic destabilization in certain lipid mixtures Single component Hong and Bowie., J Am Chem Soc. (2012) Anbazhagan and Schneider, Bioch. Biophys. Acta, (2010) E70PEITLIIFGVMAGVIGTILLISYGIR96 • Single GpA helix atomistic molecular dynamic simulations (Gromacs, Forcefield Gromos 53a6, 100ns) • Extract secondary structure which is a required input in the MARTINI model No significant change in helicity • CG simulations of a single helix: Monotonic change of thickness Tilt angle decreases with cholesterol Significant differences with ErbB2 Minimal response to the change of thickness At low tilt angles protein precession entropy is significant Lee and Im, Phys. Rev. Lett. (2008) • DPPC molecules maintain a similar arrangement • Cholesterol preferentially locates in proximity • Less change of lipids distribution around protein, less tilt angle change Pure DPPC 10% CHOL • Cholesterol promotes dimerization of GpA, for 10% and 20% CHOL • Increase of repulsive barrier of lipids drives rearrangement so that direct protein-protein increase • 10% CHOL, contact at PHE78 and ALA82 • Preliminary results support that for 30%, unfavorably lipid-induced interactions destabilize significantly the dimer • Reconstruction algorithm: simulate annealing, CG and atomistic structures are coupled via restraints Rzepiela et al., J.Comput. Chem. (2010) • Added restraints for hydrogren bonds Without Restraints Restraints E260EDFYFPWLLIIIFGIFGLTVMLFVFLFSKQQR292 • The growth hormone receptor (GHR) – class Ⅰ cytokine receptor • Important biological functions: cell division, regeneration and growth • Present model: Preformed inactive dimers. GH binding induces realignment associated with increased separation of ICDs & asymmetric ECD domain Brooks and Waters, Nature. (2010) Pang et al., PLoS.Comput.Biol. (2012) • MC free energy calculations reveal two different states in proximity • Left-handed dimer: increased Cterminal separation asymmetric tilt angles • Long AA MD simulations (1 microsecond) in DPPC bilayer • Cysteine mutations per each amino acid to compare to experimental crosslinking data • Crosslinking parallel interface inactive • Single point-mutations LH active • Linking left-handed dimer to GH:ECD receptor (PDBid:3HHR) • Generate missing residues by Modeller • Supplement our predictions with homology modeling to build the complete receptor Brooks et al, submitted • Stable in explicit bilayers for >0.5μs • Cholesterol promotes dimerization of GpA up to 20% • New Mechanism for GHR activation : Binding of GH to ECDs induces a transition from inactive parallel dimer to left-handed crossover dimer. • Current work: – General framework for activation of receptors (EPOR) – Multiscale methodology combines the efficiency of CG model and the accuracy of AA model ACKNOWLEDGMENTS • Prof. M.Waters (Univ. of Queensland). • National Science Foundation, CBET • UH Research Computing Center & High Performance Computing. ACKNOWLEDGEMENTS • Prof. M.Waters (Univ. of Queensland). • National Science Foundation, CBET NSF/CBET 1067356. • UH Research Computing Center & High Performance Computing. • IBM BlueBioU and Rice. RESEARCH AIM 1. Extends studies in multicomponent membranes • Recent published experimental data support the active role of the membrane environment for GpA 2. Multiscale Modeling of TMD association 3. Linking TMD arrangement to function CONTACT INTERFACE • In pure DPPC, contact interface GxxxG motif • 10% CHOL, contact at PHE78 and ALA82 • PHE interaction increases contact interface area, stronger proteinprotein interaction Contact interface in pure DPPC GLY79 GLY83 GLY86 Contact interface in 10% CHOL PHE78 ALA82 EXPERIMENTAL RESULTS • Asymmetric ECD structure with GH binding (active configuration) • Proximity of upper TMD leads to activation Crossing linking before 260EED (CEED) proximity of upper TMD • Mutations EED by alanines, activation concomitant to increased separation of the intracellular sequences • Crosslinking experiment without GH binding, inactive TMD dimer interface: with Phe 276 and 283, without GxxG motif • 150ps umbrella COM pulling for each configuration – average force • 32 configurations for each separation, moving average for all configuations, decreased standard deviation • CG model softer lipids, AA more repulsive forces -41kJ/mol/nm -187kJ/mol/nm