Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
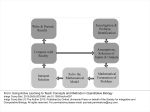
ARTICLE IN PRESS Progress in Biophysics and Molecular Biology ] (]]]]) ]]]–]]] www.elsevier.com/locate/pbiomolbio Review Modelling methodology in physiopathology Jean-Pierre Boissela,b,c,, Benjamin Ribbab,d, Emmanuel Grenierb,e, Guillemette Chapuisatb,f, Marie-Aimée Dronnea,b a CNRS, UMR5558, Lyon, France Institute for Theoretical Medicine, UMR5558, Claude Bernard University Lyon 1, RTH, Laennec School of Medicine, rue Guillaume, Paradin, 69008 Lyon, France c Léon Bérard Anticancer Centre, Lyon, France d EA, Lyon-Sud School of Medicine, Lyon, France e UMPA, UMR5669, ENS Lyon, 46 allée d’Italie, 69364 Lyon, France f CMLA, ENS Cachan, CNRS, Pres UniverSud, 94235 Cachan, France b Abstract Diseases are complex systems. Modelling them, i.e. systems physiopathology, is a quite demanding, complicated, multidimensional, multiscale process. As such, in order to achieve the goal of the model and further to optimise a rathertime and resource-consuming process, a relevant and easy to practice methodology is required. It includes guidance for validation. Also, the model development should be managed as a complicated process, along a strategy which has been elaborated in the beginning. It should be flexible enough to meet every case. A model is a representation of the available knowledge. All available knowledge does not have the same level of evidence and, further, there is a large variability of the values of all parameters (e.g. affinity constant or ionic current) across the literature. In addition, in a complex biological system there are always values lacking for a few or sometimes many parameters. All these three aspects are sources of uncertainty on the range of validity of the models and raise unsolved problems for designing a relevant model. Tools and techniques for integrating the parameter range of experimental values, level of evidence and missing data are needed. r 2007 Elsevier Ltd. All rights reserved. Keywords: Systems physiopathology; Modelling; Methodology; Strategy Contents 1. 2. Beyond systems biology, systems physiopathology . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1.1. Why should we pay attention to methodology? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1.2. Challenges and strategy . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1.3. A few general principles . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Knowledge and uncertainty management . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2 2 4 5 6 Corresponding author at: Institute for Theoretical Medicine, UMR5558, RTH Laennec School of Medicine, rue Guillaume, Paradin, 69008 Lyon, France. E-mail address: [email protected] (J.-P. Boissel). 0079-6107/$ - see front matter r 2007 Elsevier Ltd. All rights reserved. doi:10.1016/j.pbiomolbio.2007.10.005 Please cite this article as: Boissel, J.-P., et al., Modelling methodology in physiopathology. Progress Biophys Mol Biol (2008), doi:10.1016/j.pbiomolbio.2007.10.005 ARTICLE IN PRESS J.-P. Boissel et al. / Progress in Biophysics and Molecular Biology ] (]]]]) ]]]–]]] 2 3. 4. 5. 6. 7. 8. 9. Sub-models . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 6 3.1. Splitting the discursive model in sub-models . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 6 3.2. Chronology and organisational levels . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 7 Perhaps the most challenging: parameter valuation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 7 Phenomenological or mechanistic?. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 9 Top-down or bottom-up? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10 Validation. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10 Robustness . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 11 Conclusion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 11 Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 12 References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 12 1. Beyond systems biology, systems physiopathology 1.1. Why should we pay attention to methodology? Systems biology has been devised to tackle the barrier of complexity in life science. Beyond the complexity issue, numerical modelling has been introduced in physiopathology and therapeutic research because of the decreasing innovation efficiency in therapeutics and the growing difficulties for physicians to make optimal treatment decision for a given patient through combining the many available modestly efficacious treatments. Altogether, these two worrying developments result in a loss of chance for patients when compared to the amount and quality of available knowledge. The ultimate goal of ‘‘in silico’’ models in physiopathology is to improve health. Through designing numerical models of a disease and running computer simulations, we expect a better understanding of the links between its various components, weighting the respective influence of its various factors, deciphering new and more relevant targets for innovative medicines and a quicker way of testing a new therapeutic intervention at the very beginning of its development, before a sizeable amount of resources has been invested (early proof of concept). Altogether, these developments should result in improving the way new therapies are discovered and in providing doctors with tools predicting the outcome of combining therapies. Also, following the introduction of patient parameter values into the numerical models of his/her disease and of all the available therapies, a reasonable choice among all the alternatives and their combinations would be facilitated. Actually, in terms of complexity, physiopathology is far beyond systems biology. Disease mechanisms are much more complex than anything else. The main reason is that a disease develops throughout a diversity of levels, from gene expression up to population, whereas systems biology limits itself at a couple of levels, molecules and cell, sometimes a population of cells. The tissue level is seldom accounted for, and neither physiology nor anatomy is paid attention to. Diseases encompass several piled up organisational levels of complex phenomenon, from genes to population (Fig. 1). Time scales vary from nanoseconds to several decades, with chemical interactions for the former scale and evolution to clinical events for the latter. In chronic diseases, such as cancer or atherosclerosis, the sequence of events at the molecule, cell, tissue and target organ levels takes decades to achieve in death or myocardial infarction. In addition, new knowledge is emerging at a high rate: data from clinical research are coming in on top of evolving biological knowledge (Fig. 2). This observation has three consequences: (i) the ever-extensive amount of knowledge one should incorporate in a numerical model of disease; (ii) the need of a careful knowledge management tool, and (iii) models should be conceived flexible enough in order to integrate any new relevant knowledge. At first sight, it seems to be an easy task: just collect all the necessary pieces of information in the literature, and put them on a computer to draw pictures. But everyone who tried this knows that many difficulties arise. First we must find all the pieces, and even if only one is lacking, we may not achieve our objective well. Second, we must be sure that all the mathematical solutions represent properly the real world we are interested in. Third, we should solve all the methodological and technological hurdles that we faced along the process by achieving a compromise between a too stringent reduction which would make the model irrelevant to its goal Please cite this article as: Boissel, J.-P., et al., Modelling methodology in physiopathology. Progress Biophys Mol Biol (2008), doi:10.1016/j.pbiomolbio.2007.10.005 ARTICLE IN PRESS J.-P. Boissel et al. / Progress in Biophysics and Molecular Biology ] (]]]]) ]]]–]]] 3 Integration axis population organism tissue System physiopathology subcellular cellular interactions between organisational levels } molecular System biology Ph en om en on ax is Time axis interactions between phenomenon (sub-systems) Fig. 1. Complexity of systems physiopathology and systems biology. Number of published original reports 16000 14000 12000 10000 8000 6000 4000 2000 0 1970 1975 1980 1985 1990 1995 2000 2005 2010 Years Fig. 2. Number of clinical trial reports recorded in PubMed from 1970 as of today. and a too big amount of details which will make the objective impossible to reach. Fourth, it takes time to build the model and to make it run properly and in an efficient way. In addition, beyond pure science, numerical modelling of diseases is science applied to improving care to patients. As a consequence, the researcher should pay attention to both quality and transparency. For ethical reason, the former is compulsory, whereas the latter is required by regulators. Actually, regulations are already mandatory, and one can bet they will be even more stringent in the future in order to protect consumers. Models and processes which they are part of should comply with the regulations. Overall, modelling diseases develop along four axes: the phenomenon or sub-systems axis, the time axis and the integration axis (Fig. 3). Clearly, there is no unique way to design the model which would help achieving Please cite this article as: Boissel, J.-P., et al., Modelling methodology in physiopathology. Progress Biophys Mol Biol (2008), doi:10.1016/j.pbiomolbio.2007.10.005 ARTICLE IN PRESS J.-P. Boissel et al. / Progress in Biophysics and Molecular Biology ] (]]]]) ]]]–]]] 4 Integration axis Model Modelaim(s) aim(s) • Mathematical solutions • Embedding the description levels xis • Integrating sub-models a l ca eri m Time axis Ph en om en ol og ica la xi s Nu Fig. 3. The four axes along with the numerical modeller in physiopathology plays. Table 1 The challenges of systems physiopathology Amount of data Quality of data Lack of data Diversity of time scales Number and diversity of complexity levels (from molecules to population) Number and diversity of components and types of relations at each level Availability of relevant mathematical tools Computing time and precision Model validation the researcher aims. All this makes modelling in physiopathology different. It explains why an appropriate strategy and relevant methodologies are needed. 1.2. Challenges and strategy The challenges with numerical modelling in systems physiopathology are listed in Table 1. In order to take them up, one needs to follow a research strategy. The first, inescapable step is to answer the question: what do we want to model and why? A precise formulation of the objective(s) is required for a proper fixing of many choices that arise during the process. This is not solely to obtain a pure numerical representation of what is known in human physiology or to aim at a global understanding of the real system. It is not merely to build a new tool for the researcher in human biology. It is a practical objective, such as the discovery of new therapies for human diseases. The next major step is the integration of the approach in the virtuous circle shown in Fig. 4. In practice, this means bringing together a variety of expertise from mathematicians to clinicians, including computation scientists, molecular biologists and pharmacologists, and to ensure living connections with other groups working on the same theme. In short, be multidisciplinary. The other steps are tied together specifically according to every research topic. However, it is always structured along the main steps that are shown in Table 2. Unlike what comes up from this table, the sequence Please cite this article as: Boissel, J.-P., et al., Modelling methodology in physiopathology. Progress Biophys Mol Biol (2008), doi:10.1016/j.pbiomolbio.2007.10.005 ARTICLE IN PRESS J.-P. Boissel et al. / Progress in Biophysics and Molecular Biology ] (]]]]) ]]]–]]] 5 Knowledge in biology and Mathematical and medicine computing tools Observation and Numerical models experiments: in vivo, ex vivo, in vitro Experiments: in silico Fig. 4. The virtuous circle. Table 2 Major steps of a model-building strategy Setting the objective Making up and organising the multidisciplinary team Organising connections with in vitro and in vivo expert groups Selecting the knowledge management tool(s) Collecting the data Writing the discursive model Sub-setting the discursive model Choosing a phenomenological approach or a mechanistic one (or both) Finding the mathematical solutions Modelling the sub-sets of the discursive model (sub-modelling) Arranging the computational tools Writing the numerical solutions Integrating the sub-models Exploring the model robustness Reducing the model Validating the model Using the model: ‘‘in silico’’ experiments of steps is not fully linear. Step connection should not be looked at as a no-return process. Quite often, if not always, it is necessary to look back from a step to a former one. In the following we will consider some of these steps. Others, such as computing, are dealt with or illustrated elsewhere in this issue of the journal (Descombes and Dumont, 2007; Grenier et al., 2007; Dronne et al., 2007a). 1.3. A few general principles The way the challenges are dealt with depends on the main objective of the research. However, there are a few general principles that are quite helpful in finding appropriate ways to solve the problems: (i) modelling of biological processes can be piecewise, clearly identified input and output with for each piece or sub-model and biomarkers that could be used for the sake of validating the global model (this is the ‘‘Legos-like’’ principle); (ii) each piece can be numerically solved at a different level of complexity from the others; (iii) at anytime in the progress of the modelling process, a sub-model can be replaced by a more detailed one (the ‘‘plug-in’’ Please cite this article as: Boissel, J.-P., et al., Modelling methodology in physiopathology. Progress Biophys Mol Biol (2008), doi:10.1016/j.pbiomolbio.2007.10.005 ARTICLE IN PRESS J.-P. Boissel et al. / Progress in Biophysics and Molecular Biology ] (]]]]) ]]]–]]] 6 principle). However, replacement by a more detailed sub-model should be prepared (consistency in input and output) and may require new developments and re-calibration of the whole model. 2. Knowledge and uncertainty management Systems physiopathology requires collection and analysis of all the available evidence and data, before selecting those relevant for the model. Their uncertainty and strength of evidence should be weighted and recorded. The knowledge needed to model diseases such as acute stroke or cancer is enormous and diverse. Building a model in physiopathology relies on the basis of evidence that are considered as sound enough and that look crucial for integration into the model. This is a kind of ‘‘basis of knowledge’’ which will be represented in the discursive model. However, these data are embedded in uncertainty. They range from in vitro experimental results in basic biology, including structural biology, to randomised clinical trials and clinical imaging data. The first idea consists of scanning the whole literature, collecting all the possible experimental data, together with the experimental conditions, the type of cells and species, then to compare all these data and try to exclude the erroneous ones. This would give an up-to-date basis of knowledge. Because of the diversity of experimental and observational settings, values of a given parameter may vary over a wide range. Thus, it seems important to score the record by its variability and strength of evidence. In addition, identifying erroneous values is a hard task because most of the time the experimental setting is not described in such details that wrong measurements can be detected for sure. In addition, the measurements might well be correct, but the experimental setting may not be representative of the situation of interest. Instead, we may better take all the available knowledge. If moreover one takes into account the experimental errors and the individual variations, one realises that it is impossible to sort out the ‘‘true data’’. Thus, we cannot view the data extracted from the literature as definite values. An illustration of the challenge facing model builder, because of uncertain and insufficient knowledge is given in other articles in this journal issue, in published material and in Section 4. Bearing this major hurdle in mind, all the data must be collected, reviewed and stored in formats and in repositories that allow repeated access by all the team members. The development of databases or knowledge bases is thus needed to house quantitative, qualitative and structural information with a scoring of their strength of evidence (Ribba et al., 2006c). Having said that, the way of achieving such scoring remains an open question for biochemical and genetic data. One of the solutions may be to address the problem by adopting a statistical approach, i.e. meta-analysis, and a conceptualisation of the fundamentals of the empirical process by which the parameter values are determined (Grenier et al., 2007; Dronne et al., 2007a; Dronne et al., 2006). However, a lot of development is needed to achieve a relevant and valid array of tools capable of solving this critical issue. Besides data from literature, imaging and clinical data are often required (e.g. for building sham patients). These data are available somewhere. The issue here is to know where there are stored and to assess their quality. One needs an international network of such databases open to modellers. 3. Sub-models 3.1. Splitting the discursive model in sub-models The discursive model is a text or a chart (or, better, both) that brings together all the components of the disease and their interactions that are thought to be relevant enough to the objective of the modelling process. The discursive model comes out from the knowledge review mentioned above. It is the basis for the later steps (Table 2). Components and their connections will show up in the final numerical model. It is presented as a text, summarised by a chart and several series of rules. An example of the chart is given in Fig. 4 (Dronne et al., 2004). The discursive model comes in a variety of embedded forms: at the molecular level, at the cellular level, etc. Use of a dedicated software may help in mastering this step. The final discursive model could be quite intricate. Often, in physiopathology, it consists of thousands of pieces and even more functional relations. Please cite this article as: Boissel, J.-P., et al., Modelling methodology in physiopathology. Progress Biophys Mol Biol (2008), doi:10.1016/j.pbiomolbio.2007.10.005 ARTICLE IN PRESS J.-P. Boissel et al. / Progress in Biophysics and Molecular Biology ] (]]]]) ]]]–]]] 7 In the numerical modelling steps, the challenge is therefore to integrate this large amount of heterogeneous knowledge and data. A solution to this is splitting the discursive model by identifying independent subsystems. These are characterised by their studiability and the possibility of modelling them independently as sub-models, while respecting the global dynamic system. For example, in modelling acute stroke, apoptosis is described as an entire process by itself. Independence of a sub-system is defined on the basis of a couple of rules: (i) the underlying biological phenomenon has a recognised specific functional status; (ii) there are wellcharacterised signals connecting with other sub-systems (input/output of the sub-system); and (iii) it encompasses at least a biomarker which is measurable in vitro or in vivo (for the sake of validation of simulation results). Clearly, apoptosis can be viewed as a sub-system with relatively simple input/output signals, e.g. calcium concentration, energy stores as inputs, energy consumption and eventually cell death as output. 3.2. Chronology and organisational levels However, like the whole system, sub-systems have two other characteristics: one should pay attention to a chronological component and an organisational level. The molecular level is the lower level, whereas the population level is the higher level. As critical pieces of the whole discursive model, sub-systems should be organised along two dimensions, the time and organisational axes. Both axes are descriptors of the sequence of events, i.e. chronology and causal relationship. As an example, multiscale mathematical models of cancer growth have been recently proposed (Ribba et al., 2005, 2006a, b). In acute stroke, cells die first by necrosis, caused by a cellular oedema, resulting in abnormal ion exchanges because of energy deprivation (Fig. 4). Then cells of the penumbra area may die through a completed apoptotic process (Dronne et al., 2004). However, apoptosis occurs later, and can last several days. These examples show that sub-systems can be linked by causal relations and can have different chronologies. Cell and tissue mechanical properties are increasingly recognised as regulating factors of many biological processes ranging from gene transcription to tissue re-modelling. Cell elasticity is a key parameter for mechanical signal transduction (Huang et al., 2004; Janmey and Weitz, 2004), while extracellular matrix stiffness regulates cell adhesion and migration (Gray et al., 2003). Environmental mechanical forces are known to affect many cellular functions, such as cell growth, proliferation, protein synthesis and gene expression. Thus, any numerical modelling at tissue level would have to point to different sub-systems at different organisational levels that account for, e.g., cell proliferation, genetic expression and protein synthesis. It is thus clear that one of the challenges is to allow the integration of a variety of organisational levels. Moreover, computational power will soon allow the integration of cell proliferation regulated at molecular and genetic levels, as well as macroscopic changes of tissue compression and deformation. Thus, the connections between sub-models support causality and chronology also, as well as pure functional relations. In Fig. 1, the phenomenon axis is arranged according to causality. 4. Perhaps the most challenging: parameter valuation A numerical model is a series of equations or a series of rules characterised by algebraic or logical functions and parameters. The choice of functions depends on the connections or interactions between the system components or entities. Parameter values determine the spatial and temporal behaviour of the model. Thus, parameter selection and valuation are critical. Because of the uncertainty and inadequacy of available knowledge, parameter valuation is a challenging issue. This issue arises in two different settings: (i) the parameter values are drawn from the literature, i.e. the experimental data they were derived from are not accessible and (ii) the parameters are adjusted on a set of experiments to a statistical method (e.g. maximum likelihood) that allows fitting the model to the data. As the appropriate experimental data are often not accessible, the former is the most frequently used approach. We will limit this section to it. A practical example is given in Dronne et al. (2004). To illustrate the challenge and ways to make it, let us take a case where ordinary differential equations are the choice for the mathematical solution. The equation system is parameter sensitive. As in acute ischaemic stroke, let us consider a series of ionic channels of sub-system 2 in Fig. 5. The ordinary differential equations Please cite this article as: Boissel, J.-P., et al., Modelling methodology in physiopathology. Progress Biophys Mol Biol (2008), doi:10.1016/j.pbiomolbio.2007.10.005 ARTICLE IN PRESS J.-P. Boissel et al. / Progress in Biophysics and Molecular Biology ] (]]]]) ]]]–]]] 8 ococclusion c lu s ion 1 4 ¯O2 ¯O 2 ¯cellular ¯ cellular metabolismt me abolism Ri ¯ATP ¯A TP Ionic pumps, pumps, Ionic channels, channels, exchangers exchangers 5 spreading spreading depression depression glutamate glutamate uptake uptake reverse reverse glutamate --glutamate 3 -[K+]e - [K+]e oedema oedema -[Na+]i - [N a+ ]i [Ca2+ ]i --[Ca2+]i NMDA NM DA receptor receptor 2 necrosis necrosis rereperfusion p e rfu s io n AMPA-AMPA Kainate Kainate eptor receptor 9 8 apoptosis apoptosis -ATP - ATP -O2 - O2 NOS 22 NOS (astrocyte) (astrocyte) cellular cellular metabolism metabolism 10 NOS 11 NOS (neuron) (neuron) NO radicals free radicals --free inflammation inflammation 6 7 Fig. 5. Example of a chart representing a discursive model. Ten sub-systems are assumed to describe the whole process of an acute stroke. system is so large and depends on so many parameters (three equations per channel and between five and ten parameters per channel) that a small error on a parameter value may lead to a physiologically wrong behaviour. Also, different parameter values can lead to alternative, although correct, physiological behaviour. The same set of equations differing only by parameter values describes both action potential of neurons and repetitive firing and pacemaker behaviour of cardiac cells. As said above, the parameter values are not known precisely and even not known at all in many instances. Some data for glial cell ionic channels are lacking (Dronne et al., 2007b). Available data come from species or types of neurons that are the ones we want to model, and values differ across species (Hodgkin and Huxley, 1952). It is impossible to find the necessary data for all the channels of a given type of cells. There are so many possibilities that not all situations will be explored in vivo or even in vitro and most of the parameters of interest will remain unknown. Nevertheless, we have to play with all the parameters, whose values are ‘‘partially’’ known or ‘‘totally’’ unknown, in order to catch the physiologically correct behaviour. In practice, one should begin with an experimental set of parameter values for every parameter in the model and move on to a ‘‘reasonable’’ set. The experimental or observed set is directly drawn from the literature scan. It includes all the values that have been observed in rather similar experimental settings, as close as possible to the model the system is embedded in. The knowledge management tool mentioned above helps in finding the whole range. It remains to define what ‘‘reasonable’’ values mean. We can state that parameters are ‘‘reasonable’’ if the model shows physiologically relevant responses to stimuli, correct rest state and if the parameter values remain within plausible ranges. Note that this definition does not imply precise ‘‘real’’ values. It only assigns them in a range said plausible according to qualitative dynamic and resting properties of the model. These properties arise from pre-specified rules included in the discursive model and drawn from the knowledge basis. Hence the basis of knowledge must contain not only experimental data but also a qualitative description of the global behaviour. The reasonable set can be searched either using probabilistic methods or using deterministic methods. Probabilistic methods consist of choosing at random a set of parameters and checking whether it leads to correct macroscopic behaviour, i.e. whether it meet a series of rules that represent the qualitative knowledge. Please cite this article as: Boissel, J.-P., et al., Modelling methodology in physiopathology. Progress Biophys Mol Biol (2008), doi:10.1016/j.pbiomolbio.2007.10.005 ARTICLE IN PRESS J.-P. Boissel et al. / Progress in Biophysics and Molecular Biology ] (]]]]) ]]]–]]] 9 Table 3 Procedure for selecting the ‘‘reasonable’’ set Choose the experimental/observed range of all parameter values Choose at random a set of values for all parameters Test numerically the values with the mathematical model according to the rules If the rules are met, keep this set in the ‘‘reasonable’’ set With the deterministic methods, we first build a ‘‘distance’’ which measures the difference between the numerical result and the desired one, and we then try to minimise the distance. The former is easier to apply (Table 3). Taking again the ionic channels case, the basis of rules consists of: (i) existence of a stable rest equilibrium potential and (ii) a short (1 ms) strong enough imposed current leading to an action potential. These two rules can be translated into precise mathematical statements and can be easily checked by an automatic procedure. If the tested set meets the rules, it is stored. At the end of a run, a few thousands of parameter sets have been tested, leading to a few dozen of suitable sets, which can then be explored in more detail by adding more stringent criteria to the rules, e.g.: (i) an action potential is created for short-duration external stimuli; below, there should be no action potential; above, the cell is depolarised; and (ii) for longlasting external stimuli, either multiple action potentials are created or repetitive firing occurs. Of course, the more and more stringent rules we impose, the harder we get the reasonable set of parameters. In some cases it appears impossible: the rules are contradictory. If the number of rules is great then random trials are no longer efficient and must be helped by ‘‘local refinement’’ algorithms which locally increase the density of trials when almost all criteria are satisfied. As we see later in this article, there is a continuum between parameter valuation and exploration of the model robustness. 5. Phenomenological or mechanistic? A phenomenological numerical model is reduced to a representation of the envelope of the phenomenon of interest. For instance, apoptosis in acute stroke could be modelled by any mathematical equation which increases along time up to a maximum, then levels off and eventually goes down to baseline after a couple of days (Chapuisat et al., 2007). In the same issue of the journal the approach of acute stroke by Chapuisat is an illustration of a phenomenological model, whereas the approach by Dronne of the same disease is a mechanistic model (Dronne et al., 2007a; Chapuisat et al., 2007). Another rather pure example of the phenomenological approach is presented in the article by Grenier et al. (2007) in the same issue of the journal. They assume migraine as a wave phenomenon and used a mathematic descriptor of a reaction–diffusion elementary phenomenon to explore how migraine spreads in the brain. Although parameters of phenomenological models may have a biological meaning, most of the time their links with biology are somewhat indirect, and often they have no meaning at all. However, the main advantage of phenomenological models is their simplicity. On the contrary, mechanistic models aim at bringing all known details of the system. Details mean molecules—genes, proteins, ARNs, ions,y—with functional capacities and relations, e.g. signalling. Parameters in mechanistic models have direct biological or biochemical meaning, e.g. affinity constant for an enzyme, ionic conductance for an ionic channel, and they can be measured in vitro or in vivo. Obviously, the difficulty with mechanistic models is the great amount of information they should integrate, and, as discussed above, the imprecision and the lack of parameter values. The choice between the two alternatives depends on: (i) the objective of the modelling process; (ii) the availability of information regarding the system; and (iii) the chosen strategy. However, other considerations may play a role in the choice. The plug-in principle makes it possible to use a phenomenological model for a sub-system while the other sub-systems are modelled mechanistically. If the need of detailing the sub-system arises later, the phenomenological sub-model is replaced by a mechanistic one with the same entries and outputs. In addition, it looks wise to build in parallel two global models, one phenomenological and one mechanistic, or, perhaps better, to begin with the phenomenological approach in order to explore which Please cite this article as: Boissel, J.-P., et al., Modelling methodology in physiopathology. Progress Biophys Mol Biol (2008), doi:10.1016/j.pbiomolbio.2007.10.005 ARTICLE IN PRESS 10 J.-P. Boissel et al. / Progress in Biophysics and Molecular Biology ] (]]]]) ]]]–]]] sub-systems should be detailed. In such a case, parameters (and thus equations) should be chosen as direct or almost direct representative of sub-systems. And, conversely, all sub-systems should be represented by at least one parameter in the phenomenological model. Exploring the robustness of the global phenomenological model will sort out the sensitive parameters, and hence the sub-systems that need more attention. This can only be done if entries and output, hence the objective, are well defined. 6. Top-down or bottom-up? In a seminal article, Noble (2003) has spotted two opposite strategies to get into modelling a living system, the bottom-up and the top-down approaches. The former starts at the lowest organisational level (e.g. genes) with the detailed components of the system, their properties and their interactions. Higher levels are constructed from this lowest level, by assembling its components and accounting for interactions between levels. This approach assumes that, to understand the functioning of the system, it is first necessary to decipher its lowest organisational levels. Behind it, we find the idea that living systems are built from a limited number of standard parts, each amenable to standard formal model and that interactions between levels are encoded in the lower levels. A step further, we get into an entirely different field, theoretical biology (Noble, 2002). As Brenner stated, ‘‘Although I am convinced that Schrödinger’s equation underlies the behaviour of everything, it does not provide a useful basis for the construction of motor cars or bridges’’ (Brenner, 1998). The bottomup strategy is somewhat similar to the mechanistic approach. It faces the same challenges, i.e. uncertainty and lack of knowledge, and computational problems. Also, it neglects the observation that lower levels functioning depends on higher levels (see Fig. 1). According to the top-down strategy, one starts at the highest level on line with the modelling process objective, and models functions rather than entities. Then, the modeller replaces each functional block with a model of the mechanism which implements it. This strategy resembles the phenomenological approach. However, the difference is that it is a strategy, addressing the various organisational levels from the top-down to the lowest level, whereas the phenomenological approach can be used at any organisational level. However, as a phenomenological approach, the top-down strategy has been proposed as a way to characterise the basic building pieces that up a biological system. Actually, neither strategy seems the one to be followed. A compromise or middle-out, as it is called by Brenner, is more sensible (Brenner, 1998). Both the objective and the availability of data will determine where the compromise is. 7. Validation A model cannot be used to achieve the researcher’s aim without a convincing enough validation. Having said that, we would be glad to give the reader precise and relevant methods aimed at validating a model. Unfortunately, we can only propose a few general principles as the validation issue is almost entirely dependent on the objectives of the research. Assessing the validity of a model is a complex operation, based on several approaches: (i) checking the internal consistency; (ii) checking the content of the model; (iii) checking the output. The methods are different. However, they are complementary and are all prepared after the beginning of the process, at the stage of building the discursive model. Internal consistency means that the arrangement of the pieces in the model does not misrepresent the content of the discursive model which is supposed to represent all the available knowledge about the system of interest. It is neither a mathematical nor a logical operation. It consists in reviewing the details of the numerical solutions and the parameter values to check whether they correctly translate the discursive model. A detailed account of all the hypotheses (i.e., simplifying or bridging—filling lack of knowledge) made is mandatory for this operation to be relevant. Finally, the model content should fit with a few general laws, such as there is no life without energy. The validity of content of the model is addressed by looking if all the sub-model markers behave as expected from available evidence. This operation refers to data from in vitro and in vivo experiments. New such Please cite this article as: Boissel, J.-P., et al., Modelling methodology in physiopathology. Progress Biophys Mol Biol (2008), doi:10.1016/j.pbiomolbio.2007.10.005 ARTICLE IN PRESS J.-P. Boissel et al. / Progress in Biophysics and Molecular Biology ] (]]]]) ]]]–]]] 11 experiments might be requested. Examples are given in Dronne et al. (2006) with the currents of the ionic channels involved in the model. Eventually, the outputs of the model will support its validity if it is possible to compare them to available data or to results of in vivo or in vitro experiments designed on purpose. Also, whatever they are, outputs should comply with our general understanding of the living system and the particular system of interest. However, this argument should be balanced by the expectation of entirely new knowledge arising from simulations with the model. 8. Robustness If we add a piece to a formal model that has been shown to work properly, we may completely destabilise it and observe non-physiologic behaviours. In the example above, the addition of simply one channel may alter the resting potential by a few millivolts. This may be sufficient to bifurcate into another type of behaviour, for instance, an action potential type cell turns into a pacemaker type one. This question appears to be crucial in formal modelling. To be confident in the results of the global model we need to study its dependence with respect to the parameter values of its own components. If the results do not change when the parameters take ‘‘reasonable’’ values, then the qualitative results of the global model may be considered as convenient. If they change, this may be an indication that some threshold is hidden within the combination of the sub-models. Further investigations are then necessary to precise the overall behaviour. The threshold may figure out a real ‘‘in vivo’’ threshold. More often, it simply points to a weakness of the model, possibly due to a lack of our knowledge. In such case, one must encourage further in silico or in vivo investigations. Actually, exploring the robustness of a model is not entirely different from searching for the ‘‘reasonable’’ set of parameter values. An example is given with the ionic model of acute stroke by Dronne et al. (2006). In this model, 28 ionic conductances had to be specified. Three rules were pre-specified: (i) steady state at t ¼ 0; (ii) steady state reached at equilibrium; and (iii) cell depolarised when pumps do not operate. One thousand sets of conductance values were randomly drawn from the plausible ranges. One hundred and eighty of the 1000 random sets met the three rules, defining the ‘‘reasonable’’ set. Robustness was explored by simulating model behaviour in experimental situations that were used to validate the model, i.e. their results could be compared to in vivo or in vitro data or were consistent with the knowledge content of the model. All 180 sets gave qualitative as well as quantitative results that were very close to the expected results. Hence, the model was said to be robust. As we are interested in a particular behaviour of a sub-model it is rather easy to find a set of parameter values which is physiologically reasonable and which gives the desired qualitative properties. However, several different sub-models would have to be integrated in the global model—for instance, models of ionic exchanges, of apoptosis, of vascular system, of energy management (Dronne et al., 2004). Now, the question is how far does the behaviour of the global system depend on the details of each of its pieces? Sensitivity of the model behaviour to a parameter value points to this parameter. However, interpreting this finding deserves some caution. Rather than meaning that the parameter is pivotal in the system under study, it could reflect a constructed parameter, i.e. a parameter which does not exist biologically speaking and which in the model is hiding several more real parameters. In any case, sensitivity is a marker of something that should be explored more deeply. 9. Conclusion Methodology of modelling in physiopathology is a mix of strategy, tasks, work packages and validating steps structured by and organised around the biological entity that is to be modelled and the objective of the research. With an entity of a few molecules, the methodology is rather straightforward. It is almost limited to the choice of the biomarker(s) for validation, the mathematical solution, the parameter values, the simulation procedure and a few other things. With an organ or, say, a disease, appropriate strategy and methodology can make the difference between a successful model and one which does not achieve the researcher’s objective(s). Hence, deep thoughts prior to start are required. At this stage, no time spent to think is wasted. Other major initial steps in elaborating on a model-building strategy are to define the objective, then to collect and analyse the available knowledge and data to identify those that will be integrated in the model. Please cite this article as: Boissel, J.-P., et al., Modelling methodology in physiopathology. Progress Biophys Mol Biol (2008), doi:10.1016/j.pbiomolbio.2007.10.005 ARTICLE IN PRESS 12 J.-P. Boissel et al. / Progress in Biophysics and Molecular Biology ] (]]]]) ]]]–]]] The uncertainty and the strength of evidence of these data should be accounted for. Then, the researcher has to identify the pieces, that is the biological processes and entities that can be put together and separated out from the others to constitute a sub-system which will be worked out independently. This is particularly needed in the case of a disease where knowledge is scattered across different organisational levels, such as genetic, clinical, or epidemiological. Eventually, methodology in this setting has to do with epistemology. Obviously, investigating the management of unknown parameter values, which leads to a reasonable model behaviour, rises the issue of the boundaries of real world as well as the place of in silico simulation on deciphering life. Thus, one important concluding remark is the need for more research on methodology and epistemology of numerical modelling in physiopathology. Unfortunately, there is no standard recipe which would ensure a successful achievement. Art is not a fashionable buzz-word in science. However, it could well be appropriate here, beyond expertise. Acknowledgements We thank Michel (i) Cucherat, who kindly passed on Fig. 2 to the authors, and (ii) all the juniors and seniors of the Institute of Theoretical Medicine, and particularly those, besides the authors of this article, who are involved in the elaboration of the textbook on methodology of numerical modelling in physiopathology: Amine Achaachi, Frédérique Billy, Loubna El Zein, Catherine Genty, Ivanny Marchant, Patrice Nony, Nancie Raymond and Diéré Sonko. References Brenner, S., 1998. Biological computation. In: Bock, G., Goode, J.A. (Eds.), The Limits of Reductionism in Biology. Proceedings of the Novartis Foundation Symposium. Chichester, Wiley, pp. 106–116. Chapuisat, G., Dronne, M.A., Grenier, E., Hommel, M., Gilquin, H., Boissel, J.P., 2007. A global phenomenological model of ischemic stroke with stress on spreading depressions. Progress Biophys. Mol. Biol., in press, doi:10.1016/j.pbiomolbio.2007.10.004. Descombes, S., Dumont, T., 2007. Numerical simulation of a stroke: computational problems and methodology. Progress Biophys. Mol. Biol., in press, doi:10.1016/j.pbiomolbio.2007.10.003. Dronne, M.A., Boissel, J.P., Grenier, E., 2006. A mathematical model of ion movements in grey matter during a stroke. J. Theor. Biol. 240, 599–615. Dronne, M.A., Boissel, J.P., Grenier, E., Gilquin, H., Cucherat, M., Hommel, M., Barbier, E., Bricca, G., 2004. Mathematical modelling of an ischemic stroke: an integrative approach. Acta Biotheor. 52, 255–272. Dronne, M.A., Grenier, E., Chapuisat, G., Hommel, M., Boissel, J.P., 2007a. A modelling approach to explore some hypotheses of the failure of neuroprotective trials in ischemic stroke patients. Progress Biophys. Mol. Biol., in press, doi:10.1016/j.pbiomolbio. 2007.10.001. Dronne, M.A., Grenier, E., Dumont, T., Hommel, M., Boissel, J.P., 2007b. Role of astrocytes in grey matter during stroke: a modelling approach. Brain Res. 1138, 231–242. Gray, D.S., Tien, J., Chen, C.S., 2003. Repositioning of cells by mechanotaxis on surfaces with micropatterned Young’s modulus. J. Biomed. Mater. Res. A 66, 605–614. Grenier, E., Descombes, S., Dronne, M.A., Jaillard, A., Hommel, M., Boissel, J.P., 2007. A numerical study of the blocking of migraine by Rolando sulcus. Progress Biophys. Mol. Biol., in press, doi:10.1016/j.pbiomolbio.2007.10.002. Hodgkin, A.L., Huxley, A.F., 1952. A quantitative description of membrane current and its application to conduction and excitation in nerve. J. Physiol. 117, 500–544. Huang, H., Kamm, R.D., Lee, R.T., 2004. Cell mechanics and mechanotransduction: pathways, probes, and physiology. Am. J. Physiol. Cell. Physiol. 287, C1–C11. Janmey, P.A., Weitz, D.A., 2004. Dealing with mechanics: mechanisms of force transduction in cells. Trends Biochem. Sci. 29, 364–370. Noble, D., 2002. Final discussion. Is there a theoretical biology? In: Bock, G., Goode, J.A. (Eds.), ‘‘In silico’’ Simulation of Biological Processes. Proceedings of the Novartis Foundation Symposium. Wiley, Chichester, pp. 244–252. Noble, D., 2003. The future: putting Humpty-Dumpty together again. Biochem. Soc. Trans. 31, 156–158. Ribba, B., Marron, K., Agur, Z., Alarcón, T., Maini, P.K., 2005. A mathematical model of Doxorubicin treatment efficacy for nonHodgkin’s lymphoma: investigation of the current protocol through theoretical modelling results. Bull. Math. Biol. 67, 79–99. Ribba, B., Colin, T., Schnell, S., 2006a. A multiscale mathematical model of cancer, and its use in analyzing irradiation therapies. Theor. Biol. Med. Model. 10, 3–7. Ribba, B., Saut, O., Colin, T., Bresch, D., Grenier, E., Boissel, J.P., 2006b. A multiscale mathematical model of avascular tumor growth to investigate the therapeutic benefit of anti-invasive agents. J. Theor. Biol. 243, 532–541. Ribba, B., Tracqui, P., Boix, J.-L., Boissel, J.-P., Thomas, S.R., 2006c. Generic database for modelling—QxDB: a generic database to support mathematical modeling in biology. Philos. Trans. A 364, 1517–1532. Please cite this article as: Boissel, J.-P., et al., Modelling methodology in physiopathology. Progress Biophys Mol Biol (2008), doi:10.1016/j.pbiomolbio.2007.10.005