Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
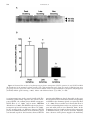
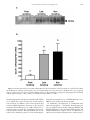
Changes in gene expression of glucose transporters in lactating and nonlactating cows1 T. Komatsu*2, F. Itoh*, S. Kushibiki*, and K. Hodate† *Department of Animal Physiology and Nutrition, National Institute of Livestock and Grassland Science, Tsukuba, Ibaraki 305-0901, Japan; and †School of Veterinary Medicine and Animal Science, Kitasato University, Towada, Aomori 034-8628, Japan amount of GLUT1, whereas the adipose tissue of peaklactating cows did not (P < 0.05). There were no significant differences in the abundance of GLUT4 mRNA in adipose tissue and muscle, whereas GLUT4 mRNA was not detected in the mammary gland. The plasma insulin concentration was greater (P < 0.05) in nonlactating cows than in peak- and late-lactating cows. The results of the present study indicate that in lactation, GLUT1 expression in the mammary gland and adipose tissue is a major factor for insulin-independent glucose metabolism, and the expression of GLUT4 in muscle and adipose tissue is not an important factor in insulin resistance in lactation; however, the plasma insulin concentration may play a role in insulin-dependent glucose metabolism. Factors other than GLUT4 may be involved in insulin resistance. ABSTRACT: Glucose delivery and uptake by the mammary gland are a rate-limiting step in milk synthesis. It is thought that insulin-independent glucose uptake decreases in tissues, except for the mammary gland, and insulin resistance in the whole body increases following the onset of lactation. To study glucose metabolism in peak-, late-, and nonlactating cows, the expression of erythrocyte-type glucose transporter (GLUT1) and the insulin-responsive glucose transporter (GLUT4) in the mammary gland, adipose tissue, and muscle were assessed by Western blotting and realtime PCR. Our results demonstrated that the mammary gland of lactating cows expressed a large amount of GLUT1, whereas the mammary gland of nonlactating cows did not (P < 0.05). On the other hand, adipose tissue of late and nonlactating cows expressed a large Key Words: Adipose Tissue, Cows, Glucose Transporter, Lactation, Mammary Gland 2005 American Society of Animal Science. All rights reserved. Introduction J. Anim. Sci. 2005. 83:557–564 cose transporter in the lactating bovine mammary gland (Zhao et al., 1996), but whether GLUT1 expression changes during the period of lactation is unknown. Insulin-responsive glucose transporter (GLUT4) is the transporter isoform primarily responsible for insulin-stimulated glucose transport and is therefore found mainly in insulin-sensitive tissues, such as fat and muscle (James et al., 1989). In lactation, it is considered that the increase in glucose supply to the mammary gland is partially mediated by an increase in wholebody insulin resistance (McDowell et al., 1987), but the change of expression of GLUT4 during lactation has not been extensively investigated. In ruminants, GH is a galactopoietic hormone (i.e., the systemic administration of GH to cows markedly increases milk production). Growth hormone augments peripheral insulin resistance in dairy cows following the i.v. injection of insulin (Rose et al., 1997). It has been reported that the plasma insulin concentration is lower in lactating cows than in nonlactating cows (Peel et al., 1983). The objectives of this study were to investigate changes in expression of GLUT1 and GLUT4 related During lactation, lactose synthesis from glucose in the mammary epithelial cells seems to be the rate-limiting step in milk synthesis. To supply glucose to the mammary gland, insulin-independent glucose uptake decreases in tissues, except for the mammary gland, and insulin resistance in the whole body increases at peak lactation (McDowell et al., 1987); however, the mechanisms during lactation are still unclear. Insulin-independent glucose transporter (GLUT1) is expressed in many tissues, including the brain, kidney, and mammary gland (Burant et al., 1991). It is reported that GLUT1 may be the predominant facilitative glu- 1 The authors thank the staff of the Ruminants and Field Management Section, Natl. Inst. of Livest. and Grassl. Sci., for technical assistance and animal management. 2 Correspondence: Ikenodai 2 (phone: 81-29-838-8645; fax: 81-29838-8606; e-mail: [email protected]). Received June 24, 2004. Accepted November 29, 2004. 557 558 Komatsu et al. to glucose metabolism in peak-, late-, and nonlactating cows, and to clarify the mechanisms of the decrease of insulin-independent glucose uptake and the increase of insulin resistance during lactation. Materials and Methods Animals and Tissues Four peak-lactating (8 to 11 wk after parturition), four late-lactating (40 to 50 wk after parturition), and four nonlactating (dried off for 3 to 10 wk) Holstein cows were used in this experiment. Cows were fed a diet based on a total mixed ration, so as to maintain BW (63.7% DM, 66.9% TDN, 13.5% CP, and 42.8% NDF; DM basis) and milk yield (88.0% DM, 79.7% TDN, 18.75% CP, and 26.4% NDF; DM basis) according to the Japanese Feeding Standard for Dairy Cattle (Agriculture, Forestry, and Fisheries Research Council Secretariat, 1999). The diets for lactating and nonlactating cows were quartered and supplied at 0400, 1000, 1600, and 2200 each day using an automatic feeder, and cows were fasted for 12 h before slaughter. Lactating cows were milked at 0830 and 1800. The mean milk yield of the cows on the day before slaughter was 27.9 ± 2.51 kg/d in peak-lactating cows and 18.5 ± 2.16 kg/day in late-lactating cows. The mean BW of the cows were 580 ± 62.6 kg in peak-lactating cows, 665.8 ± 33.4kg in latelactating cows, and 657 ± 41.5 kg in nonlactating cows. All animals received humane care as outlined in the Guide for the Care and Use of Experimental Animals (NILGS, 2002). Two 5-g of samples of intestinal adipose tissue, mammary gland, and skeletal muscle (pectoralis prefundis) were collected immediately after slaughter, and frozen in liquid N. The biopsy site of mammary gland selected was in the basal (upper) portion of the left udder. Fat and large s.c. blood vessels were avoided whenever possible during incision. Samples were stored at −80°C before protein and RNA extraction. RNA Isolation and cDNA Synthesis Total RNA was extracted with TRIzol reagent (Invitrogen, Leek, The Netherlands) according to the manufacturer’s protocol, then treated with DNase I to remove residual genomic DNA, and quantified spectrophotometrically by light absorbance at a wavelength of 260 nM; the ratio of absorbance at 260 nM to that at 280 nM was always greater than 1.8. The method was modified and optimized for extraction of RNA from adipose tissue. As opposed to the manufacturer’s protocol, we used 10 mL of TRIzol/g of adipose tissue and added 4 mL of chloroform. The cDNA was synthesized from 100 ng of total RNA using a random hexamer (TaKaRa, Tokyo, Japan) and Murine Moloney leukemia virus reverse transcriptase (Invitrogen, Carlsbad, CA). Western Blotting Tissue GLUT1 was analyzed by Western blotting as reported previously (Abe et al., 2001). Tissues were ho- mogenized in PBS containing 1 g/mL of aprotinin and 1 mM phenylmethylsulfonyl fluoride using a Polytron homogenizer, and centrifuged at 3,000 × g for 20 min at 4°C. Protein concentrations of the supernatant fraction were measured using a BCA protein assay reagent kit (Pierce, Rockford, IL) and BSA as a standard. The supernatant fraction was subjected to 10% SDS-PAGE, electrophoretically transferred to a polyvinylidene difluoride membrane, and blocked overnight at 4°C in blocking buffer (Tris-buffered saline with 0.1% Triton X-100 Sigma, St. Louis, MO] containing 5% nonfat dried milk [Meiji, Tokyo, Japan]). Membranes were incubated with diluted (1:500) GLUT1 antiserum (Calbiochem, San Diego, CA) in blocking buffer at room temperature for 1 h. The membranes were then washed five times at room temperature for 10 min in Tris-buffered saline with 0.1% Triton X-100 and incubated for 1 h at room temperature in blocking buffer, with a 1:3,000 dilution of goat anti-rabbit immunoglobulin G conjugated with horseradish peroxidase (Zymed, San Francisco CA). The luminescent signals on membranes were visualized using an enhanced chemiluminescence detection kit (Amersham, Buckinghamshire, U.K.) according to the manufacturer’s instructions, followed by exposure to x-ray film for 5 min. The resulting films were analyzed by scanning densitometry. All samples of mammary gland and adipose tissue were treated on each blot. Real-Time PCR from cDNA GLUT1 and GLUT4 mRNA. The abundance of GLUT1 and GLUT4 mRNA was measured with the use of gene-specific double-fluorescent-labeled probes, TaqMan Universal PCR Mix and a 7700 Sequence Detector (Applied Biosystems, Foster, CA). Six-carboxyfluorescein and VIC (Applied Biosystems, Foster, CA) were used as the 5′-fluorescent reporters, while 6-carboxy-tetramethyl-rhodamine (TAMRA) was added to the 3′ end as a quencher. To compensate for variations in input RNA amounts and the efficiency of reverse transcription, the housekeeping gene’s glyceraldehyde3-phosphate dehydrogenase mRNA was quantified and results were normalized to these values. The PCR amplification was performed using the primer and probe (Applied Biosystems) sets outlined in Table 1. Primers and TaqMan probes were designed using the computer program Primer Express (Applied Biosystems). The PCR was performed at 95°C for 10 min, followed by 45 cycles at 94°C for 30 s, 59°C for 30 s, and 72°C for 30 s. Use of the TaqMan probe resulted in reliable and sensitive quantification of the real-time PCR (RT-PCR) product with good linearity (Pearson correlation coefficient r > 0.99, not shown). All samples were all within the standard curve. Growth Hormone Receptor mRNA. Growth hormone receptor (GHR) mRNA expression was measured by RT-PCR using a LightCycler (Roche Diagnostics, Mannheim, Germany) instrument with the QuantiTect 559 Gene expression of GLUT in lactating cows Table 1. Primers and probes for real-time polymerase chain reactiona Item Genbank Accession No. GLUT1 NM_174602 5′-GTGCTCCTGGTTCTGTTCTTCA-3′ 5′-GCCAGAAGCAATCTCATCGAA-3′ 5′-FAM-TCAAAGTTCCCGAGACAAAAGGCCG-TAMRA-3′ GLUT4 D63150 5′-GGACCGCGAATAGAAGAAAGAC-3′ 5′-CAACTTCATCATCGGCATGG-3′ 5′-FAM-TAGGGACCCATAGCATCCGCCACA-TAMRA-3′ GAPDH BTU85042 5′-TGACCCCTTCATTGACCTTCA-3′ 5′-GCCTTGACTGTGCCGTTGA-3′ 5′-VIC-TTCCAGTATGATTCCACCCACGGCA-TAMRA-3′ GHR NM_176608 5′-ACAACGCTTACTTCTGCGAGGTA-3′ 5′-TTCCTGGTTAAAGCTTGGCTCTAC-3′ GAPDH BTU85042 5′-CACCCTCAAGATTGTCAGCA-3′ 5′-GGTCATAAGTCCCTCCACGA-3′ Primer Probe a FAM = six-carboxy-fluorescein; TAMRA = 6-carboxy-tetramethyl-rhodamine; GAPDH = glyceraldehyde-3-phosphate dehydrogenase; GLUT1 = insulin-independent glucose transporter; GLUT4 = insulin-responsive glucose transporter; GHR = growth hormone receptor. SYBR Green PCR system (Qiagen K. K., Tokyo, Japan). The PCR amplification was performed using the primer sets outlined in Table 1. The primers were chosen with an online software package (www-genome.wi.mit.edu/ cgi-bin/primer/primer3_www.cgi). The PCR was performed at 95°C for 15 min, followed by 50 cycles at 94°C for 15 s, 55°C for 20 s, and 72°C for 10 s. All samples were all within the standard curve. GH and Insulin Assays Venous blood samples were collected at 0800 on the day of slaughter. Plasma was harvested from the blood samples and stored at −20°C for later determination of GH and insulin. Plasma GH concentrations were determined by RIA as described by Johke (1978). Plasma insulin concentrations were determined using a commercially available RIA kit (Eiken Chemical, Tokyo, Japan). Statistical Analyses All values were expressed as means ± SEM. The data were analyzed by one-way ANOVA with the StatView 5 (SAS Inst., Inc., Cary, NC) software package. When the ANOVA showed a significant effect for the phase, phases were compared by the Fisher’s protected LSD as a multiple comparison test. A P-value less than 0.05 denoted the presence of a statistically significant difference. Results The expression of GLUT1 protein and mRNA was assessed in the bovine mammary gland and adipose tissue at various stages of lactation. It has been reported that the antibody used in this study recognized GLUT1 protein in bovine tissues (Abe et al., 2001). In the mammary glands of peak- and late-lactating cows, Western blotting analysis showed that GLUT1 was detected with similar intensity, whereas in nonlactating cows, it was barely detectable (Figure 1A). The antibody of GLUT1 could specifically recognize separate 42- and 45-kDa proteins. In the mammary gland, RT-PCR analysis also showed that the mRNA level of bovine GLUT1 in peak- and late-lactating cows was three times greater (P < 0.05) than that in nonlactating cows (Figure 1B). In adipose tissue, GLUT1 protein expression was detected in late- and nonlactating cows, whereas it was barely detectable in peak-lactating cows (Figure 2A). Unlike in the mammary gland, it was detected as a single band in adipose tissue. Use of the TaqMan probe resulted in reliable and sensitive quantification of the RT-PCR product, with good linearity (Person correlation coefficient r > 0.99, not shown). Values were standardized to glyceraldehyde-3-phosphate dehydrogenase and expressed as a percentage of the control. Realtime PCR analysis also showed that the mRNA level of GLUT1 in late and nonlactating cows was six times greater (P < 0.05) than that in peak-lactating cows (Figure 2B). In muscle, GLUT1 protein and mRNA were not detected in at any stage cows (data not shown). There were no significant differences in the abundance of GLUT4 mRNA among the peak-, late-, and nonlactating cows in the adipose tissue (Figure 3A) or muscle (Figure 3B), and GLUT4 mRNA was not detected in the mammary gland of the cows in any lactating stage (data not shown). There were no significant differences in the abundance of GHR mRNA among cows in the various lactating stages in the mammary gland or adipose tissue (Figures 4A and B). The insulin concentration was greater (P < 0.05) in nonlactating than in peak- and late-lactating cows (Figure 5A). In contrast, there were no significant differences in the concentrations of GH among the cows in the various stages (Figure 5B). Discussion Glucose is the major precursor for lactose synthesis. Therefore, the supply of glucose to the mammary gland 560 Komatsu et al. Figure 1. Western blot analysis of erythrocyte-type glucose transporter (GLUT1) protein (A) and GLUT1 mRNA (B) abundance in the mammary gland in peak-, late-, and nonlactating cows. Data are means ± SEM for four cows per group and are expressed as the ratio of GLUT1 mRNA relative to glyceraldehyde-3-phosphate dehydrogenase (GAPDH) mRNA (peak lactating = 100%). Means with different letters (a, b) differ, P < 0.05. is an important factor in the control of milk yield. The mammary gland mainly expresses the glucose transporter GLUT1, the sodium glucose-linked transporter SGLT (Zhao et al., 1999), and in mouse, GLUT12 (Macheda et al., 2003). The GLUT1 is the major glucose transporter species in lactating cows (Zhao et al., 1996). In the present study, no difference between GLUT1 protein and mRNA abundance in the mammary gland was detected at peak vs. late lactation, whereas GLUT1 protein and mRNA were barely detectable in dry cows. Western blot analysis showed two bands corresponding to GLUT1 in the mammary gland, as reported by Zhao et al. (1996). Tracer studies have shown that there is a linear relationship between the rate of glucose transport and milk yield in cows (Kronfeld, 1982). In the mammary gland, we found no change in the expression of GLUT1 between peak and late lactation, implying that a decrease of the glucose supply to the mammary Gene expression of GLUT in lactating cows 561 Figure 2. Western blot analysis of insulin-independent glucose transporter (GLUT1) protein (A), and GLUT1 mRNA (B) abundance in adipose tissue in peak-, late-, and nonlactating cows. Data are means ± SEM for four cows per group and are expressed as the ratio of GLUT1 mRNA relative to glyceraldehyde-3-phosphate dehydrogenase (GAPDH) mRNA (peak lactating = 100%). Means with different letters differ, P < 0.05. gland and apoptosis of mammary epithelial cells (Wilde et al., 1997) may cause the decrease of milk yield at late lactation. In addition, it has been reported that resistin mRNA abundance decreases in the lactating mammary gland (Komatsu et al., 2003). Therefore, resistin may serve to make the regulation of glucose uptake by insulin effective. Because GLUT4 was not detected in the mammary gland, a novel insulin-depen- dent glucose transporter (e.g., GLUT12; Macheda et al., 2003) may be involved in glucose uptake. In ruminants, triacylglycerol is synthesized from glycerol-3 phosphate, a metabolite derived from glucose, and adipose tissue mainly expresses GLUT1 and GLUT4. Although we also showed GLUT1 protein and mRNA abundance in adipose tissue, no difference of expression was observed between non- and late-lactat- 562 Komatsu et al. Figure 3. Insulin-responsive glucose transporter (GLUT4) mRNA abundance in adipose tissue (A) and muscle (B) in peak-, late-, and nonlactating cows. Data are means ± SEM for four cows per group and are expressed as the ratio of GLUT4 mRNA relative to glyceraldehyde-3-phosphate dehydrogenase (GAPDH) mRNA (peak lactating = 100%). The GLUT4 mRNA was not detected in the mammary gland. ing cows, whereas hardly any expression was detected in peak-lactating cows. Western blot analysis showed a single band, in contrast with the case of mammary glands. The cause of this difference is unknown. The Figure 4. Growth hormone receptor (GHR) gene abundance in mammary gland (A) and adipose tissue (B) in peak-, late, and nonlactating cows. Data are means ± SEM for four cows per group and are expressed as the ratio of GHR mRNA relative to glyceraldehyde-3-phosphate dehydrogenase (GAPDH) mRNA (peak lactating = 100%). decrease of GLUT1 expression in adipose tissue in peak lactation may contribute to the increased supply of plasma glucose to the mammary gland. The GLUT4 mRNA levels detected in the adipose tissue and muscle did not vary significantly among the various stages of lactation in cows. It is considered that insulin resistance 563 Gene expression of GLUT in lactating cows GHR mRNA levels detected in the mammary gland and adipose tissue did not significantly vary in the various stages of lactating cows. It has been reported that administration of GH to dairy cows increases milk yield (Pocius et al., 1986), so the effect may be caused via GHR activation in addition to the GH concentration, not by regulation of the abundance of GHR mRNA and protein. In conclusion, GLUT1 seems to be the major glucose transporter isoform in the bovine mammary gland and adipose tissue during lactation. The change in insulinindependent glucose uptake during lactation was associated with a change in the expression of GLUT1. We did not detect any change of GLUT4 expression in adipose tissue or muscle, which suggests that the change of insulin-dependent glucose uptake in lactation is not regulated by GLUT4 expression. Insulin concentration and GLUT4 activation may be involved in regulating lactation. Plasma insulin concentration may play a role in regulating insulin-dependent glucose metabolism, but the expression of GLUT4 in the mammary gland and adipose tissue is not an important factor of insulin resistance in lactation. Literature Cited Figure 5. Plasma insulin (A) and GH (B) concentrations in peak-, late-, and nonlactating cows. Data are means ± SEM for four cows per group. Means with different letters (a, b) differ, P < 0.05. increases during lactation (Rose et al., 1997). Therefore, during lactation, the promotion of insulin resistance in adipose tissue may be caused by inhibition of GLUT4 translocation to the cytomembrane, not by inhibition of GLUT4 expression. It has been reported that plasma GH concentration is higher in lactating vs. nonlactating cows (Vasilatos et al., 1980), but our results showed no difference of GH concentration in lactating vs. nonlactating cows. A difference might be seen if the change in GH concentration of individual cows with time were studied. The Abe, H., Y. Kawakita, K. Hodate, and M. Saito. 2001. Postnatal development of glucose transporter proteins in bovine skeletal muscle and adipose tissue. J. Vet. Med. Sci. 63:1071–1075. Agriculture, Forestry, and Fisheries Research Council Secretariat. 1999. Japanese Feeding Standard for Dairy Cattle. Agric. Forestry Fisheries Res. Counc. Secretariat, Tokyo, Japan. Burant, C. F., W. I. Sivitz, H. Fukumoto, T. Kayano, S. Nagamatsu, S. Seino, J. E. Pessin, and G. I. Bell. 1991. Mammalian glucose transporters: Structure and molecular regulation. Recent Prog. Horm. Res. 47:349–387. James, D. E., M. Strube, and M. Mueckler. 1989. Molecular cloning and characterization of an insulin-regulatable glucose transporter. Nature 338:83–87. Johke, T. 1978. Effects of TRH on circulating growth hormone, prolactin and triiodothyronine levels in the bovine. Endocrinol. Jpn. 25:19–26. Komatsu, T., F. Itoh, S. Mikawa, and K. Hodate. 2003. Gene expression of resistin in adipose tissue and mammary gland of lactating and nonlactating cows. J. Endocrinol. 178:R1–R5. Kronfeld, D. S. 1982. Major metabolic determinants of milk volume, mammary efficiency, and spontaneous ketosis in dairy cows. J. Dairy. Sci. 65:2204–2212. Macheda, M. L., E. D. Williams, J. D. Best, M. E. Wlodek, and S. Rogers. 2003. Expression and localisation of GLUT1 and GLUT12 glucose transporters in the pregnant and lactating rat mammary gland. Cell Tissue Res. 311:91–97. McDowell, G. H., J. M. Gooden, D. Leenanuruksa, M. Jois, and A. W. English. 1987. Effects of exogenous growth hormone on milk production and nutrient uptake by muscle and mammary tissues of dairy cows in mid-lactation. Aust. J. Biol. Sci. 40:295–306. NILGS. 2002. Guide for the Care and Use of Experimental Animals. Natl. Inst. of Livest. and Grassl. Sci. Anim. Care Comm., Tsukuba, Japan. Peel, C. J., T. J. Fronk, D. E. Bauman, and R. C. Gorewit. 1983. Effect of exogenous growth hormone in early and late lactation on lactational performance of dairy cows. J. Dairy Sci. 66:776–782. 564 Komatsu et al. Pocius, P. A., and J. H. Herbein. 1986. Effects of in vivo administration of growth hormone on milk production and in vitro hepatic metabolism in dairy cattle. J. Dairy Sci. 69:713–720. Rose, M. T., Y. Obara, F. Itoh, H. Hashimoto, and Y. Takahashi. 1997. Non-insulin- and insulin-mediated glucose uptake in dairy cows. J. Dairy Res. 64:341–353. Vasilatos, R., and P. J. Wangsness. 1980. Changes in concentrations of insulin, growth hormone and metabolites in plasma with spontaneous feeding in lactating dairy cows. J. Nutr. 110:1479–1487. Wilde, C. J., C. V. Addey, P. Li, and D. G. Fernig. 1997. Programmed cell death in bovine mammary tissue during lactation and involution. Exp. Physiol. 82:943–953. Zhao, F. Q., W. T. Dixon, and J. J. Kennelly. 1996. Localization and gene expression of glucose transporters in bovine mammary gland. Comp. Biochem. Physiol. B. Biochem. Mol. Biol. 115:127–134. Zhao, F. Q., E. K. Okine, and J. J. Kennelly. 1999. Glucose transporter gene expression in bovine mammary gland. J. Anim. Sci. 77:2517–2522.