Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
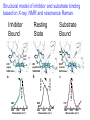
Enzyme Kinetics Michaelis-Menten Theory Dehaloperoxidase: Multi-functional Enzyme NC State University Michaelis-Menton kinetics The rate of an enzyme catalyzed reaction in which substrate S is converted into products P depends on the concentration of the enzyme E even though the enzyme does not undergo any net change. k aon k bcat E + S ↔ ES → P + E k ′ aoff Michaelis-Menton rate equations ka kb E + S ↔ ES → P + E k ′ a Steps in the Michaelis-Menton mechanism Step 1. Bimolecular formation of the enzyme E and and substrate S: E+S ES rate of formation of ES = kon[E][S] Step 2. Unimolecular decomposition of the complex: ES E + S rate of decomposition of ES = -koff[ES] Step 3. Formation of products and release from the enzyme: ES P+E rate of formation of P = kcat[ES] The rate law of interest is the formation of the product in terms of E and S. The enzyme substrate complex can be eliminated The enzyme substrate complex is formed transiently and can be approximated using the steady state approximation. The result of this approximation is Pseudo-first order Michaelis-Menton kinetics In an experiment we know the total enzyme concentration [E]0 and not the unbound enzyme [E]. The total concentration of enzyme [E]0 = [E] + [ES]. which rearranges to Pseudo-first order Michaelis-Menton kinetics At this point it is convenient to define the Michaelis constant and to rearrange the equations as Michaelis-Menton parameters The rate of formation of product can be written where Km is the Michaelis constant and kcat is the maximum turnover number. We often make the definitions which permit us to write the equation as Limiting conditions of enzyme reactivity • Maximal rate: If there is excess substrate present the rate is limited by the rate at which the ES complex falls apart. The rate of formation of products is a maximum and vmax = kcat[E]0is called the maximum velocity. • Second order regime: If [S] << KM then the rate of formation of products is d[P]/dt = kcat/Km [E]0[S]. The rate depends on [S] as well as [E]0. • A plot of 1/k yields kcat and Km but not the rate constants kon and koff. The latter rate constants can be obtained from stopped-flow experiments. General expression for reaction velocity Based on the previous analysis the velocity at an arbitrary substrate concentration is: Lineweaver-Burke Plots • The Michaelis-Menton expression is non-linear. • The Lineweaver-Burke plot is linearized plot of data. 1 = K M + [S ] = 1 + K M 1 v [S ]vmax vmax vmax [S ] • This expression has the form of an equation for a line: y = intercept + slope x • Such plots are not necessary today with common non-linear fitting programs. Transition State Stabilization The original idea of the enzyme having maximum complementarity to the TS was put forward by Linus Pauling in 1946. It wasn't until the early 70's that the idea was put on a more solid grounding. As put forward by Lienhard and Wolfenden the idea is as follows: Kn‡ kn E+S E + S‡ E+P Ks ES Kc‡ Kt ES‡ kc E+P Transition State Stabilization Defining the equilibrium constants as association constants: Kn‡ = [S‡]/[S] , Kt = [ES‡]/[E][S‡] from TS theory: ∆G‡ = -RT ln K‡ and kobs = (kBT/h)e-∆G‡/RT Thus, kn = (kBT/h)Kn‡ and kc = (kBT/h)Kc‡ where c means catalyzed and n means uncatalyzed. From the scheme you can see that Ks Kc‡ = Kn‡ Kt hence Kt/Ks = Kc‡/Kn‡ however, kc/kn = Kc‡/Kn‡ Therefore the observed rate enhancement kc/kn = Kt/Ks >> 1 Therefore the transition state geometry S‡ must bind more tightly than the substrate S in its equilibrium geometry! Transition State Analogs The transition state stabilization hypothesis was tested by designing so-called transition state analogs, molecules which mimick the real TS as closely as possible. One of the first enzymes examined was proline racemase: - N H COO H N H H COO- N COO- H The compound on the right is a planar TS state analog. This molecule was found to be a good inhibitor, with Ki some two orders of magnitude smaller than Km. The Role of Entropy In a seminal paper Page and Jencks showed that the loss in entropy in going from a bimolecular to a unimolecular reaction, i.e. E + S <=> ES, could account for as much as 108 of the observed rate enhancement. In other words, this much free energy would come from the intrinsic binding energy. The entropy loss arises from the loss of translational and rotational degrees of freedom when the substrate is bound. The configurational entropy is: S = kB lnW where W is the number of degrees of freedom available to a molecule. Inhibition An inhibitor is any compound that causes a decrease in the catalytic rate. We will consider non-covalent ligands that can bind to the enzyme. The general scheme is shown below: S E I EI S kc ES I EIS ki E+P EI + P I = inhibitor Inhibition occurs if ki[EIS] < kc[ES] Competitive Inhibition Competitive inhibition results from the direct competition between the I and S for the substrate binding site. There is an additional equilibrium constant: EI E +I [E ][I ] KI = [EI ] The velocity under these conditions turns out to be: [S ]vmax v= αK M + [S ] α=1+ [I ] KI Uncompetitive Inhibition Uncompetitive inhibition arises when I can bind at site that is not the same as the substrate binding site. There is an additional equilibrium constant: EI E +I [E ][I ] KI = [EI ] Here the complex IE indicates that the inhibitor does not bind in the same site as the substrate. The velocity under these conditions is: vmax[S ] v= K M + α[S ] α=1+ [I ] KI DHP has a natural peroxidase function Engineered globin peroxidases Mauk group Watanabe group DHP O X - O X + H2O2 X Trihalogenated Phenol (X = I, Br, Cl, F) DHP X X + H2 O + X O Horseradish Peroxidase Dihalogenated Quinone - Structural model of inhibitor and substrate binding based on X-ray, NMR and resonance Raman. Inhibitor Bound Resting State Substrate Bound TXP 4-XP (Int) H55 (equilibrium) 5cHS/6cHS 1494 5cHS ν3 c ν3 1480 6cHS b ν3 1483 6cHS a H55 (closed) 6cHS Heme 1494 5cHS H55 (open) 5cHS Heme 4BP DHP TCP 1460 1500 1480 Wavenumber (cm-1) 1460 1500 1480 Wavenumber (cm-1) 1460 1500 1480 Wavenumber (cm-1) Kinetic analysis showing competitive inhibition under native conditions Absorbance Product/4-BP Product 2,4,6-TBP 2,4,6-TBP Wavelength (nm) Wavelength (nm) OH Br Br + H2O2 Br OH OH Br Br + Br + H2O2 Br Kinetic analysis showing competitive inhibition mechanism a 1.5 b 1.5 1.0 2,4,6-TCP 0.5 0.0 2,6-DCQ 250 300 350 0.2 400 2,4,6-TCP 0.5 250 300 350 400 Wavelength (nm) Vmax 30 Vo 20 1/Vo 10 Fe(IV)=O inhibited 360 Product/4-BP 1.0 d DHP DHP Cmp ES Cmp ES + 20µM 4BP Cmp ES + 40µM 4BP Cmp ES + 100µM 4BP Cmp ES + 200µM 4BP Cmp ES + 400µM 4BP 0.4 DHP + Substrate + Inhibitor 0.0 400 Wavelength (nm) c 0.6 Absorbance DHP + Substrate Absorbance Absorbance TCP Substrate Substrate + Inhibitor 1/[S] 0 440 Wavelength (nm) 480 0 5 10 15 Concentration TCP (mM) 20