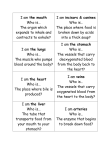
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
NSC 203 HUMAN PHYSIOLOGY (II) NATIONAL OPEN UNIVERSITY OF NIGERIA SCHOOL OF HEALTH SCIENCES COURSE CODE: NSC 203 COURSE TITLE: HUMAN PHYSIOLOGY II COURSE UNITS: 4 (3-1-2) CONFIDENTIAL – NO PART OF THIS DOCUMENT MAY BE COPIED – PROPERTY OF NOUN NURSING NSC 203 NSS 323 HUMAN PHYSIOLOGY (II) NATIONAL OPEN UNIVERSITY OF NIGERIA COURSE GUIDE NSC 203 – HUMAN PHYSIOLOGY II COURSE CODE: NSC 203 COURSE TITLE: Human Physiology II COURSE UNITS: 5 Credit units (36 hours of instruction online; 12 hours of Discussion forum online/tutorial; 48 hours of laboratory practical) YEAR: 2 SEMESTER: 1st Semester PRE-REQUISITE COURSES: All courses in the BNSC degree programme in the first year CON-CURRENT COURSES: All courses at the 200, NSC 201, 205, 207, 209 SESSION: _________________ COURSE WEBSITE: www.noun.edu.ng/ COURSE WRITERS Dr. Rufus O. Akomolafe, PhD Dr. Abiodun O. Ayoka, PhD Dr. Olumide S. Akinsomisoye - PhD Olaoluwa S. Olukiran (Co-writer) – MSc Department of Physiology, Obafemi Awolowo University, Ile-Ife COURSE EDITORS: Dr O.O. Irinoye and Dr T.O. Oladogba COURSE COORDINATOR: _________________________ COURSE FACILITATORS: _________________________ 2 COURSE GUIDE NSC 203 HUMAN PHYSIOLOGY (II) COURSE GUIDE Table of Contents General Introduction Course Aims Course Objectives Working through the Course Course Materials Study Units Reference Textbooks Equipment and Software Needed to Access Course Number and Places of Meeting Discussion Forum Course Evaluation Grading Criteria Grading Scale Schedule of Assignments with Dates Course Overview How to get the most from this Course 3 Page 4 4 4 4 5 5 6 6 6 7 7 7 7 7 8 8 NSC 203 HUMAN PHYSIOLOGY (II) COURSE GUIDE GENERAL INTRODUCTION Welcome to the second year course in Human Physiology, NSC 203 – Human Physiology II. In the first level course, we learn that Physiology is about how the body does the work of helping us to attain a state of health. We also learnt that the nurse must be adequately grounded in Physiology to help her/him in determining the functionality of the various organs that also work within the systemic framework. The knowldedge of normal function is also the basis of diagnosing disturbances of physiological processes. COURSE OBJECTIVES At the completion of this course, you should be able to: Apply the knowledge of respiratory and gastrointestinal physiology in analyzing health and nursing care needs and planning care of patients. WORKING THROUGH THIS COURSE The course will be delivered adopting the blended learning mode, 70% of online but interactive sessions and 30% of face-to-face during laboratory sessions. You are expected to register for this course online before you can have access to all the materials and have access to the class sessions online. You will have hard and soft copies of course materials, you will also have online interactive sessions, face-to-face sessions with instructors during practical sessions in the laboratory. The interactive online activities will be available to you on the course link on the Website of NOUN. There are activities and assignments online for every unit every week. It is important that you visit the course sites weekly and do all assignments to meet deadlines and to contribute to the topical issues that would be raised for everyone’s contribution. You will be expected to read every module along with all assigned readings to prepare you to have meaningful contributions to all sessions and to complete all activities. It is important that you attempt all the Self Assessment Questions (SAQ) at the end of every unit to help your understanding of the contents and to help you prepare for the in-course tests and the final examination. You will also be expected to keep a portfolio where you keep all your completed assignments. COURSE MATERIALS Course Guide Course Text in Study Units Textbooks (Hard and electronic) Book of Laboratory Practical Assignment File/Portfolio STUDY UNITS This course is made up of 2 modules comprising 17 units as listed below: MODULE 3- Respiratory Physiology Unit 1: Components, muscles of respiration and mechanism of breathing Unit 2: Surfactant and compliance of the lungs, the dead space, lung volumes and capacity 4 NSC 203 HUMAN PHYSIOLOGY (II) Transport and Exchange of Gases, Oxygen, Carbon dioxide and the Oxygen – haemoglobin dissociation curve Unit 4: Control of Respiration and Respiratory Adaptations in Unusual Environment MODULE 4- Gastrointestinal Physiology Unit One Organization and Secretions of the Gastrointestinal Tract Unit Two Gastrointestinal Motility Unit Three Gastrointestinal Hormones, Digestion And Absorption Unit 3: REFERENCE TEXTBOOKS Fox SI. (2012). Human Physiology. 12th edition, Mc Graw Hill, New York. Ganong WF. (2010). Review of Medical Physiology. 23rd edition, Mc Graw Hill, New York. Guyton AC, Hall JE. (2001). Textbook of Medical Physiology. Harcourt International Edition, 10th edition, W.B. Saunders, Philadelphia. Oyebola DO. (2002). Essential Physiology, Vol 1, Nihort Press. COURSE REQUIREMENTS AND EXPECTATIONS OF YOU Attendance of 95% of all interactive sessions, submission of all assignments to meet deadlines; participation in all CMA, attendance of all laboratory sessions with evidence as provided in the log book, submission of reports from all laboratory practical sessions and attendance of the final course examination. You are also expected to: 1. Be versatile in basic computer skills 2. Participate in all laboratory practical up to 90% of the time 3. Submit personal reports from laboratory practical sessions on schedule 4. Log in to the class online discussion board at least once a week and contribute to ongoing discussions. 5. Contribute actively to group seminar presentations. EQUIPMENT AND SOFTWARE NEEDED TO ACCESS COURSE You will be expected to have the following tools: 1. A computer (laptop or desktop or a tablet) 2. Internet access, preferably broadband rather than dial-up access 3. MS Office software – Word PROCESSOR, Powerpoint, Spreadsheet 4. nBrowser – Preferably Internet Explorer, Moxilla Firefox 5. Adobe Acrobat Reader NUMBER AND PLACES OF MEETING (ONLINE, FACE-TO-FACE, LABORATORY PRACTICALS) The details of these will be provided to you at the time of commencement of this course DISCUSSION FORUM There will be an online discussion forum and topics for discussion will be available for your contributions. It is mandatory that you participate in every discussion every week. Your participation links you, your face, your ideas and views to that of every member of the class and earns you some mark. COURSE EVALUATION There are two forms of evaluation of the progress you are making in this course. The first are the series of activities, assignments and end of unit, computer or tutor marked assignments, and laboratory practical sessions and the report. These constitute the continuous assessment that all carry 30% of the total mark. The second is a written examination with multiple choice, short 5 NSC 203 HUMAN PHYSIOLOGY (II) answers and essay questions that take 70% of the total mark that you will do on completion of the course. Students’ evaluation: The students will be assessed and evaluated based on the following criteria: o In-Course Examination: In line with the university’s regulation, in-course examination will come up in the middle of the semester These would come in form of Computer Marked Assignment. This will be in addition to 1compulsory Tutor Marked Assignment (TMA’s) and three Computer marked Assignment that comes after every module….. o Laboratory practical: Attendance, record of participation and other assignments will be graded and added to the other scores form other forms of examinations. o Final Examination: The final written examination will come up at the end of the semester comprising essay and objective questions covering all the contents covered in the course. The final examination will amount to 60% of the total grade for the course. Learner-Facilitator evaluation of the course This will be done through group review, written assessment of learning (theory and laboratory practical) by you and the facilitators. GRADING CRITERIA Grades will be based on the following Percentages Tutor Marked Individual Assignments 10% Computer marked Assignment 10% Group assignment 5% 30% Discussion Topic participation 5% Laboratory practical 10% End of Course examination 70% GRADING SCALE A = 70-100 B = 60 - 69 C= 50 - 59 F = < 49 SCHEDULE OF ASSIGNMENTS WITH DATES To be provided for each module by the facilitator in addition to the ones already spelt out in the course materials. SPECIFIC READING ASSIGNMENTS To be provided by each module COURSE OVERVIEW NSC 203 - Human Physiology (II) This course is in continuation of NSC 104, Human Physiology (I) where we covered the functional cell, cardio-vascular/cardio-pulmonary physiology. In this course we would cover respiratory and gastrointestinal physiology. Respiration is the commonly acknowledged sign of life. 6 NSC 203 HUMAN PHYSIOLOGY (II) HOW TO GET THE MOST FROM THIS COURSE 1. Read and understand the context of this course by reading through this course guide paying attention to details. You must know the requirements before you will do well. 2. Develop a study plan for yourself. 3. Follow instructions about registration and master expectations in terms of reading, participation in discussion forum, end of unit and module assignments, laboratory practical and other directives given by the course coordinator, facilitators and tutors. 4. Read your course texts and other reference textbooks. 5. Listen to audio files, watch the video clips and consult websites when given. 6. Participate actively in online discussion forum and make sure you are in touch with your study group and your course coordinator. 7. Submit your assignments as at when due. 8. Work ahead of the interactive sessions. 9. Work through your assignments when returned to you and do not wait until when examination is approaching before resolving any challenge you have with any unit or any topic. 10. Keep in touch with your study centre, the NOUN, School of Health Sciences websites as information will be provided continuously on these sites. 11. Be optimistic about doing well. 7 NSC 203 HUMAN PHYSIOLOGY (II) COURSE CONTENTS Table of Contents MODULE 3- Respiratory Physiology Unit 1: Components, muscles of respiration and mechanism of breathing Unit 2: Surfactant and compliance of the lungs, the dead space, lung volumes and capacity Unit 3: Transport and Exchange of Gases, Oxygen, Carbon dioxide and the Oxygen – haemoglobin dissociation curve Unit 4: Control of Respiration and Respiratory Adaptations in Unusual Environment Page 9 9 13 MODULE 4- Gastrointestinal Physiology Unit 1 Organization and Secretions of the Gastrointestinal Tract Unit 2 Gastrointestinal Motility Unit 3 Gastrointestinal Hormones, Digestion And Absorption 39 39 50 59 8 19 29 NSC 203 HUMAN PHYSIOLOGY (II) MODULE 3 - RESPIRATORY PHYSIOLOGY Introduction A person is alive when he or she demonstrates the act of respiration. Several processes are involved in the exchange of gases at different levels of the organs involving diverse structures. Effective functioning of the organs of respiration, especially the lungs are measurable and this module provides you with the information about how the lungs perform the functions of gaseous exchange. Module Objectives At the end of this module, you should be able to: Discuss the various processes involved in act of exchange of gases in the body for sustenance of life at the levels of the cells, organs and the body as a whole. CONTENT Unit 1: Unit 2: Unit 3: Unit 4: UNIT 1: Components, muscles of respiration and mechanism of breathing Surfactant and compliance of the lungs, the dead space, lung volumes and capacity Transport and Exchange of Gases, Oxygen, Carbon dioxide and the Oxygen – haemoglobin dissociation curve Control of Respiration and Respiratory Adaptations in Unusual Environment COMPONENTS, MUSCLES OF RESPIRATION AND MECHANISM OF BREATHING CONTENT 1.0 2.0 3.0 3.1 3.2 3.3 4.0 5.0 6.0 6.1 6.2 7.0 Introduction Objectives Main Contents Components of the respiratory system Muscles of respiration Mechanism of respiration Conclusion Summary Tutor Marked Assignments Activity Tutor Marked Tests References and other resources 1.0 Introduction Respiration has two major components namely (a) the transport of oxygen from the outside air to the cells within tissues, and the transport of carbon dioxide from the cells to the outside air and (b) the utilization of oxygen within the body cells for the liberation of energy from food substances. The former is known as external respiration or gaseous exchange while the latter is called internal or tissue respiration. The respiratory system functions in close collaboration with 9 NSC 203 HUMAN PHYSIOLOGY (II) the circulatory system which acts as the transport system that conveys oxygen from the respiratory apparatus to the tissues and carbon-dioxide from the tissues to the respiratory apparatus for expulsion from the body. The respiratory system is thus an ―air pump‖ while the cardiovascular system is a ―blood pump‖. This unit covers the n 2.0 Objective At the end of this unit, you should be able to: i. Explain the component parts of the respiratory system ii. Explain the composition of the categories of muscles involved in respiration iii. Describe the mechanism of respiration 3.0 Main Content 3.1 The pulmonary circulation Venous blood from tissues of the body is returned to the right atrium of the heart. From here, the blood enters the right ventricle. The right ventricle pumps the blood out of the heart through the pulmonary artery. The pulmonary arterial trunk divides into the right and left pulmonary arteries and these supply blood to the right and left lungs respectively. The pulmonary arteries divide several times just like the trachea until the pulmonary capillaries are formed. The pulmonary circulation is a low pressure circulation. Blood is pumped out of the right ventricle at a pressure of 25/0 mmHg. By the time blood reaches the pulmonary capillaries, the pressure has fallen to an average of 10mmHg. Since the plasma oncotic pressure is 25mmHg, no fluid moves out of the pulmonary capillaries into the interstitial space. This is important because if fluid moves out of the arterial end of the pulmonary capillaries as it does in the systemic circulation, then, the small diameter air sacs (alveoli) can become ―flooded’’ with tissue fluid and this will adversely affect the transport of gases in the lungs. The blood pumped into the pulmonary circulation at rest is 5L/min, the same as that pumped out of the left side of the heart per minute at rest. Blood from the pulmonary capillaries is returned to the left atrium of the heart through the pulmonary vein. The pulmonary artery carries deoxygenated blood while the pulmonary vein carries oxygenated blood. About 2% of the blood flow to the lungs is through the bronchial arteries and veins. The left bronchial artery arises directly from the aorta while the right bronchial artery arises from the first right intercostal artery. The bronchial arteries run along the bronchi and follow them into the lung. They supply the air passages, their glands and sub pleural connective tissue. The bronchial veins, which carry deoxygenated blood join the pulmonary vein so that the latter which was 100% saturated with oxygen in the lungs is diluted by the addition of venous blood from the bronchial veins. The oxygen in the blood that is delivered to the left atrium is therefore about 97%. This mixing of deoxygenated bronchial venous blood with oxygenated pulmonary venous blood is called physiological shunting. 3.2 Capillary/alveolar membrane The capillary/alveolar membrane (Figure 2-2) is very thin and this makes it easy for gases to diffuse across the membrane. The large surface area of the alveoli is matched by an even larger surface area presented by the capillaries. Since the capillaries are 10 micrometer in diameter and the red blood cells are 7.2 micrometer in their larger diameter, many of the red blood cells will cross the pulmonary capillaries in a ―queue’’ of single cells. This also exposes a large surface area of RBC for gaseous exchange. 10 NSC 203 HUMAN PHYSIOLOGY (II) Figure 1: Ultrastructure of the respiratory membrane 3.2 Muscles of respiration These can be divided into two broad groups: i. Muscles of inspiration ii. Muscles of expiration The main muscles of inspiration are the diaphragm (innervated by the phrenic nerve, from C3, 4 and 5) and the external intercostal muscles. The accessory muscles of inspiration are sternocleido-mastoids, scalenes, serratus anterior, levator scapulae, erectus spinae and pectoralis major and minor. Expiration is normally a passive process under quiet breathing. But in strenuous exercise, when there is difficulty with breathing or in forced expiration as in sneezing, the internal intercostal muscles are used. The accessory muscles of expiration are the abdominal recti and posterior inferior serratus muscle. There are two types of intercostal muscles in each of the eleven intercostal spaces, the internal and external intercostal muscles. They are supplied by intercostal nerves from adjacent intercostal nerve roots. The external intercostal run forwards and downwards and they pull the ribs forward and upward. The internal intercostal run backwards and downwards and pull the ribs backward and downward. The diaphragm is the dome-shaped musculo-tendinous partition between the thorax and abdomen, forming the roof of the abdomen and the floor of the thorax. The bony thorax, the intercostal muscles and the diaphragm form a cone shaped structure, called the thoracic cavity (Figure 2.1). 11 NSC 203 HUMAN PHYSIOLOGY (II) The distance from the thoracic inlet to the diaphragm is the vertical diameter of the thorax. The vertical diameter can increase in size when the diaphragm contracts and moves downwards. The distance between the posterior surface of the sternum and the anterior surface of the vertebral column is the antero-posterior (AP) diameter. When the ribs are pulled upward and forward, the AP diameter increases. Figure 2: Contraction and expansion of the thoracic cage during expiration and inspiration. 3.3 Mechanism of Breathing When the diaphragm contracts, it moves down, thereby increasing the vertical diameter of the thoracic cavity. The diaphragm is capable of vertical excursion of 2.5cm to 10cm depending on the depth of breathing. An increase in thoracic volume due to diaphragm causes the intrathoracic pressure to be 2 to 6 mmHg less than atmospheric pressure. Since pressure outside (atmospheric air) is higher than the pressure in the thoracic cavity, atmospheric air rushes into the lungs and fills the lungs. In deep breathing, contraction of the external intercostal muscles will pull the ribs upwards and move the sternum forward thereby increasing the AP diameter. This will increase the thoracic volume further and make the intrathoracic pressure more negative, so that more air will fill the lungs. As air rushes in to fill the lungs, the lungs and the chest wall expand. This expansion stretches the elastic tissues of the lungs and the chest wall. At the end of inspiration, the stretched elastic tissues relax and this causes the lungs and the thoracic wall to recoil passively. At the end of inspiration, the diaphragm relaxes and it is pushed up to a dome-shaped position by the abdominal viscera. This leads to a reduction of the vertical diameter of the thoracic cage. The thoracic cage and its elastic recoil and the lung tissues makes the thoracic volume to become smaller so that the pressure inside the thorax is now greater than the pressure outside (Po). This higher pressure compresses the lungs so that air is expelled from the lungs. During quiet breathing expiration is a passive process, relying on the elastic recoil of the lung and chest wall. When ventilation is increased, such as during exercise, expiration becomes active with contraction of the muscles of the abdominal wall and the internal intercostals muscle. 4.0 Conclusion 12 NSC 203 HUMAN PHYSIOLOGY (II) The respiratory system serves as the air pump that allows for external and internal exchange of gases from the external environment and the internal environment of the body. The two groups of muscles are for inspiration and expiration. The mechanism of labour is facilitatated by the movement of the diaphragm as it contracts, moves down or up, thereby increasing or decreasing the vertical diameter of the thoracic cavity to allow air entry or air push-out. 5.0 Summary In this unit, you have learnt that air exchange is facilitated through pulmonary circulation, and at the level of the capillary membrane/alveolar membrane. The main muscles of inspiration are the diaphragm and the external intercostal muscles and are supported by the accessory muscles of inspiration including the sternocleido-mastoids, scalenes, serratus anterior, levator scapulae, erectus spinae and pectoralis major and minor. 6.0 6.1 Tutor Marked Assignments Activity: This will be provided by the Facilitator 6.2 1. Explain how the structural makeup of the pulmonary circulation, the capillary and alveolar membranes facilitate exchange of gases from the outside of the body and within the body. 2. Enumerate the muscles in their groupings involved in respiration 3. Describe the mechanism of respiration 7.0 References and other resources Fox, S.I. (2012). Human Physiology. 12th edition, Mc Graw Hill, New York. Ganong W.F. (2010). Review of Medical Physiology. 23rd edition, Mc Graw Hill, New York. Guyton, A.C& Hall J.E. (2001). Textbook of Medical Physiology. Harcourt International Edition, 10th edition, W.B. Saunders, Philadelphia. Oyebola, D.O. (2002). Essential Physiology, Vol 1, Nihort Press UNIT 2: SURFACTANT AND COMPLIANCE OF THE LUNGS, THE DEAD SPACE, LUNG VOLUMES AND CAPACITY CONTENT 1.0 Introduction 2.0 Objectives 3.0 Main Content 3.1 Surfactant and compliance of the lungs 3.2 Anatomic and Physiologic dead Space 3.3 Lung Volumes 3.4 Lung Capacity 4.0 Conclusion 5.0 Summary 6.0 Tutor Marked Assignments 6.1 Activity 6.2 Tutor Marked Tests 13 NSC 203 HUMAN PHYSIOLOGY (II) 7.0 References and other resources 1.0 Introduction The lung is able to perform its function of facilitating air intake and exchange by the presence of a special fluid and this helps in getting the lungs to hold some volume of air. The amount of air in the lungs can be measured using various measures. In this unit, you will cover these and how some of the measures are taken. 2.0 Objectives At the end of this unit, you should be able to: i. Explain in detail the Surfactant and compliance of the lungs ii. Describe the Anatomic and Physiologic dead Space iii. Describe the process of Spirometry iv. Describe lung volumes and lung capacity as measures of pulmonary function. 3.0 Main Content 3.1 Surfactant and compliance of the lungs Surfactant is a surface acting agent that is responsible for lowering the surface tension of a fluid. The surfactant that lines the epithelium of the alveoli is the pulmonary surfactant and it decreases the surface tension on the alveolar membrane. The pulmonary surfactant is secreted by the Type II alveolar epithelial cells in the lungs. Surfactant is a lipoprotein complex formed by lipids especially phospholipids, proteins and ions. In infants, lack or absence of surfactant causes Respiratory Distress Syndrome or Hyaline Membrane Disease. Figure 4.1 is a diagram relating lung volume changes to changes in transpulmonary pressure. Compliance means the volume change in the lungs produced by a unit change of pressure. It is a measure of the distensibility (elasticity) of the lungs and thoracic structures. The extent to which the lungs expand for each unit increase in transpulmonary pressure is called their compliance. The total compliance of both lungs together in normal adult human being averages about 200 millimeters of air per centimeter of water transpulmonary pressure. That is, every time the transpulmonary pressure increases by 1 centimeter of water, the lung volume expands 200 millimeters. Tissues of the lungs and thorax exhibit elastic properties so that when a force (pressure) is applied, the resulting volume change is proportional to the applied force within limits. Thus, under static conditions, when pressure is increased, volume is increased and the lungs and thoracic wall are stretched. When the increase in pressure is removed, the elastic properties of the tissue restore the original volume. Compliance (C) is given by the ratio: Change in volume Change in pressure The unit of compliance is Liters/cmH20. 3.2 Anatomic and Physiologic dead Space In the lungs, exchange of gases occurs only in the respiratory bronchioles and the alveoli. Air in the air passages does not take part in gaseous exchange. The anatomical structures that make up the air passages in which no gaseous exchange occurs are the nasal cavity, the pharynx, the 14 NSC 203 HUMAN PHYSIOLOGY (II) larynx, the trachea, the bronchi and the bronchioles up to the terminal bronchioles. The volume of air inside this conduit is the anatomic dead space (ADS). Physiologic dead space is the anatomic dead space plus the volume of areas of the lungs that are not taking part in gaseous exchange. Such non-functional areas result from poor or absent perfusion in a well-ventilated lung. In a perfectly healthy person, there is no such non-functional area in the lungs, so that the anatomic dead space is equal to the physiologic dead space. Spirometry The volume of air that moves into and out of the lungs under different conditions can be measured using a spirometer. The process is called spirometry. A typical basic spirometer is shown in Figure 3-3. It consists of a drum containing air or oxygen inverted over a slightly bigger drum containing water. A string is attached to the inverted drum and this is passed over pulleys to a counter balancing weight. A writing point is attached to the device so that movements of the inverted drum can be recorded on a kymograph. The apparatus has a mouthpiece through which a subject breathes in and out. The end of the mouthpiece is above the water level. When the subject inspires from the spirometer, the volumes of air inside the inverted drum is reduced, the inverted drum sinks lower into the water and the writing point on the other side moves up. The reverse occurs during expiration. So an upward deflection is inspiration and a downward deflection is expiration. During spirometry, many of the volumes are measured starting from the resting expiratory level. The resting expiratory level is the volume of air present in the lungs following quiet expiration. In the latter condition, elastic recoil of the lungs and thoracic cage causes the lungs to be passively compressed to a relaxed state. Figure 3: Spirometer 3.3 Lung Volumes 15 NSC 203 HUMAN PHYSIOLOGY (II) Lung volumes are the volumes of air breathed by an individual. There are four volumes; i.) Tidal Volume (TV): This is the volume of air breathed in and out of the lungs during normal quiet respiration. It has a value of 500ml (0.5L) ii) Inspiratory Reserve Volume (IRV): This is the volume of air that can be forcefully inspired after a normal inspiration. It has an average value of 3300ml (3.3L) iii.) Expiratory Reserve Volume (ERV): This is volume of air that can be forcefully expired after normal expiration. iv) Residual Volume: This is the volume of air remaining in the lungs even after a most forceful expiration. This volume of air cannot be emptied from the lungs. It has a value of 1200ml (1.2L). 3.4 Lung Capacities Lung capacities are the combination of two or more lung volumes. There are four types of lung capacities; i. Inspiratory Capacity (IC): This is the maximum volume of air that is inspired after normal expiration. This is also TV + IRV. It has a value of about 3800ml (3.8L). ii. Vital Capicity (VC): This is the maximum volume of air that can be forcefully expired after a maximal inspiratory effort. This is also TV + IRV + ERV. It has a value of about 4800ml (4.8L). iii. Functional Residual Capacity (FRC): This is the volume of air remaining in the lungs after normal tidal expiration. This is also ERV + RV. It has a value of about 2200ml (2.2L). iv. Total Lung Capacity (TLC): This is the volume of air present in the lungs after a inspiratory effort. This is also IRV + TV + ERV + RV. It has a value of about 6000ml (6.0L). capacity plus the residual volume (VC + RV). 16 NSC 203 HUMAN PHYSIOLOGY (II) Figure 4: Diagram showing respiratory excursions during normal breathing and during maximal inspiration and maximal expiration. The most common parameters measured in spirometry are vital capacity (VC), forced vital capacity (FVC), forced expiratory volume (FEV) at timed intervals of 0.5, 1.0 (FEV1), 2.0, and 3.0 seconds, forced expiratory flow 25–75% (FEF 25–75) and maximal voluntary ventilation (MVV), also known as maximum breathing capacity. Forced vital capacity (FVC) is the volume of air (measured in liters) that can forcibly be blown out after full inspiration. FVC is the most basic maneuver in spirometry tests. Forced expiratory volume in 1 second (FEV1): Average values for FEV1 in healthy people depend mainly on sex and age. Values of between 80% and 120% of the average value are considered normal. FEV1/FVC (FEV1%) is the ratio of FEV1 to FVC. In healthy adults this should be approximately 75–80%. In obstructive diseases (asthma, COPD, chronic bronchitis, emphysema) FEV1 is diminished because of increased airway resistance to expiratory flow; the FVC may be decreased as well, due to the premature closure of airway in expiration, just not in the same proportion as FEV1 (for instance, both FEV1 and FVC are reduced, but FEV1 is more affected because of the increased airway resistance). This generates a reduced value (<80%, often ~45%). In restrictive diseases (such as pulmonary fibrosis) the FEV1 and FVC are both reduced proportionally and the value may be normal or even increased as a result of decreased lung compliance. 3.3 Minute respiratory volume and alveolar ventilation The normal respiratory rate is 12 to 16 times per minute. The volume of fresh air moved into the respiratory system per minute is the minute respiratory volume or total ventilation. This volume is the product of the tidal volume (T.V.) and the respiratory rate (R.R.) i.e T.V x R.R 500ml/breath x 12 breaths/minute = 6000ml/min or 6L/min. Alveolar ventilation is the volume of fresh air that enters the alveoli per minute. It is equal to the tidal volume minus the dead space volume (VD) multiplied by respiratory rate (RR). i.e. Alveolar ventilation= (TV- VD) x RR Thus, if the TV is 500ml, and VD is 150ml and RR is 12/min, Then, Alveolar ventilation = (500-150) x12 = 350 x 12 ml/min = 4200ml/min. 4.0 Conclusion The tissues of the lungs and thorax exhibit elastic properties that enables measurable amount of air to be inhaled and exhaled. These measurable volumes of air are determined to affirm the quality of functioning of the lungs. 5.0 Summary In this unit, you have leant about the use of surfactant in getting compliant lungs. You have also learnt about the structures of the respiratory system that are not actively involved in respiration. You have learnt about the various measures that can be used to measure lung functions. 6.0 6.1 Tutor Marked Assignments Activity: Compile the patients with respiratory disorders in you place of work. Share 17 NSC 203 HUMAN PHYSIOLOGY (II) their diagnoses in your discussion forum and in groups as will be assigned to you, explain the homeostatic imbalance associated with their diseases. In your reference textbooks, or from the Internet look for a diagram that shows compliance of the lungs in a healthy person. 6.2 Answer the following questions: 1. What is the disease associated with the defect of surfactant and how will you explain what happens? 2. Describe how spirometry is conducted. 3. What are the lung function tests that can be performed and what are the normal values of such measures? 7.0 References and other resources Fox, S.I. (2012). Human Physiology. 12th edition, Mc Graw Hill, New York. Ganong W.F. (2010). Review of Medical Physiology. 23rd edition, Mc Graw Hill, New York. Guyton, A.C& Hall J.E. (2001). Textbook of Medical Physiology. Harcourt International Edition, 10th edition, W.B. Saunders, Philadelphia. Oyebola, D.O. (2002). Essential Physiology, Vol 1, Nihort Press UNIT 3: TRANSPORT AND EXCHANGE OF GASES - OXYGEN, CARBON DIOXIDE AND THE OXYGEN–HAEMOGLOBIN DISSOCIATION CURVE CONTENT 1.0 Introduction 2.0 Objectives 3.0 Main Content 3.1 Transport and exchange of gases 3.2 The oxygen–haemoglobin dissociation curve 4.0 Conclusion 5.0 Summary 6.0 Tutor Marked Assignments 6.1 Activity 6.2 Tutor Marked Tests 7.0 References and other resources 1.0 Introduction The process of exchange of oxygen with carbon dioxide and vice-versa uses pressure gradients that are supported by some factors. The red blood also play important role in oxygen transport to 18 NSC 203 HUMAN PHYSIOLOGY (II) tissues. In this unit the steps by step of the processes and the chemical reactions with the equations are explained in this unit. 2.0 Objectives At the end of this unit, you should be able to: i. Explain the process of Gaseous transport and exchange ii. Explain the process of oxygen transportation iii. List various steps involved in oxygen transportation iv. List the major forms through which oxygen can be transported in the blood. v. Describe the Give detailed explanation about Oxygen dissociation curve. vi. Enumerate the various factors that can cause oxygen- hemoglobin dissociation curve to shift to the left or shift to the right. vii. Explain the various steps involved in the transportation of the Carbon-dioxide. viii. Explain the Ventilation – Perfusion relationship citing relevant examples. 3.0 Main Content 3.1 Transport and exchange of gases Gaseous exchange is the process by which oxygen is transferred from the atmosphere to the tissue for use in metabolism; and the gas produced by metabolism, carbon dioxide, is transferred from tissues to the atmosphere. Gaseous exchange is divided into the processes of alveolar ventilation (bringing air into the lungs for transfer of oxygen and carbon dioxide) and pulmonary circulation (bringing blood to the lungs to take up oxygen and excrete carbon dioxide). The process of gas exchange has several steps. The following is a summary of the steps: i. Ventilation (breathing) ii. Interchange of CO2 and O2 between air in the lungs' alveoli and blood in lung capillaries by diffusion iii. Transport of CO2 and O2 through the bloodstream iv. Interchange of CO2 and O2 between blood in lung capillaries and alveolar air by diffusion v. Use of O2 and production of CO2 by cells through metabolism. Upon inhalation, gas exchange occurs at the alveoli, the tiny sacs which are the basic functional component of the lungs. The alveolar walls are extremely thin (approximately 0.2 micrometers). These walls are composed of a single layer of epithelial cells (type I and type II epithelial cells) close to the pulmonary capillaries which are composed of a single layer of endothelial cells. The close proximity of these two cell types allows permeability to gases and, hence, gas exchange. This whole mechanism of gas exchange is carried by the simple phenomenon of pressure difference. Whenever the atmospheric pressure is lower than the pressure inside the lungs, the air from lungs flows out, but when the pressure in the lungs is lower than atmospheric, air rushes into the lungs. To accomplish gas exchange the air that is inhale is delivered, via the mouth and nose, to tiny sacs, called alveoli, which are the terminal or end units of the airways. Oxygen from the air diffuses across a thin membrane into tiny blood capillaries surrounding the alveoli. At the same time CO2 diffuses from the blood capillaries into the alveoli and out of the lungs with each exhalation. The combination of one alveolus (containing air) and its surrounding capillaries 19 NSC 203 HUMAN PHYSIOLOGY (II) (containing blood) is called an alveolar-capillary unit. At the alveolar-capillary membrane, gas exchange takes place. Oxygen is delivered to, and carbon dioxide removed from, the capillary blood. This gas exchange converts the oxygen-poor blood entering the pulmonary capillary into oxygen-rich blood. Partial pressures of O2 and CO2 in the body (normal, resting conditions): PO2 = 100 mm Hg PCO2 = 40 mm Hg Alveolar capillaries Entering the alveolar capillaries PO2 = 40 mm Hg (relatively low because this blood has just returned from the systemic circulation and has lost much of its oxygen) PCO2 = 45 mm Hg (relatively high because the blood returning from the systemic circulation has picked up carbon dioxide). While in the alveolar capillaries, the diffusion of gasses occurs: oxygen diffuses from the alveoli into the blood and carbon dioxide from the blood into the alveoli. Leaving the alveolar capillaries PO2 = 100 mm Hg PCO2 = 40 mm Hg Blood leaving the alveolar capillaries returns to the left atrium and is pumped by the left ventricle into the systemic circulation. This blood travels through arteries and arterioles and into the systemic, or body, capillaries. As blood travels through arteries and arterioles, no gas exchange occurs. Entering the systemic capillaries PO2 = 100 mm Hg PCO2 = 40 mm Hg Body cells (resting conditions) PO2 = 40 mm Hg PCO2 = 45 mm Hg Because of the differences in partial pressures of oxygen and carbon dioxide in the systemic capillaries and the body cells, oxygen diffuses from the blood and into the cells, while carbon dioxide diffuses from the cells into the blood. Leaving the systemic capillaries PO2 = 40 mm Hg PCO2 = 45 mm Hg Blood leaving the systemic capillaries returns to the heart (right atrium) via venules and veins (and no gas exchange occurs while blood is in venules and veins). This blood is then pumped to the lungs (and the alveolar capillaries) by the right ventricle. 20 NSC 203 HUMAN PHYSIOLOGY (II) Figure 5: Partial pressures of gases in blood 3.2 Transport of oxygen and carbon-dioxide Transport of oxygen The process by which atmospheric oxygen gets to the tissues for use in metabolic processes is referred to as oxygen transport. Oxygen transport consists of four important steps: i. Movement of oxygen from atmospheric air into the alveoli (inspiration) ii. Diffusion of oxygen from the alveolar sac into the blood in the pulmonary capillaries iii. Transport of oxygen in the blood from the lungs to the tissues. iv. Delivery of oxygen from the systemic capillary blood to the tissues. Fresh atmospheric air moves into the alveoli during inspiration. This is an active process involving contraction of the diaphragm with or without contraction of the external intercostal muscles. In the alveoli, the PO2 is 100mmHg, while PO2 in pulmonary arterial capillaries is 40mmHg. There is therefore a gradient for oxygen to diffuse from the alveolar sacs into the pulmonary capillary blood. The capillary/alveolar membrane is very thin and the surface area of the alveolar sacs and pulmonary capillaries is very large. These factors favour rapid diffusion of oxygen from the alveoli into the capillaries. Oxygen is transported in two forms in the blood, as shown in Figure 8.1: (i) O2 dissolved in plasma (ii) O2 carried in combination with hemoglobin in the red blood cell. 21 NSC 203 HUMAN PHYSIOLOGY (II) Dissolved O2 in plasma The plasma is a poor carrier of oxygen. At a PO2 of 100 mmHg, 100 ml of plasma can carry only 0.3ml of oxygen, while at a PO2 of 40 mmHg, dissolved oxygen is 0.13 ml/100ml. Nonetheless, oxygen diffuses from the region of high tension in the alveolar sacs (100 mmHg) into the region of low tension in the plasma (40mmHg). The oxygen that diffuses into the plasma dissolves in the plasma and the diffusion continues until the PO2 in plasma rises to 100mmHg. At this point of equilibration, the plasma contains only 0.3ml of dissolved O2 per 100ml. However, the saturation of plasma with oxygen exposes the red blood cells suspended in the plasma to a high oxygen tension. Oxygen diffuses rapidly from the plasma into the red blood cells and combines with hemoglobin. 3.3 Oxygen in combination with hemoglobin The haem portion of the hemoglobin molecule contains four molecules of ferrous iron. This iron is capable of a reversible combination with oxygen, with the iron still remaining in the ferrous form. Since the iron is not converted to ferric iron, the reaction is not an oxidation, but an oxygenation. The normal hemoglobin concentration is about 14.5g/dl. Each gram of hemoglobin is capable of carrying 1.34ml of oxygen at full saturation. Then, 100ml of blood at full saturation will carry 19.7ml of oxygen (19.4ml combined with Hb and 0.3ml dissolved in plasma). Each molecule of hemoglobin (which contains 4Hb units) combines with four molecules of oxygen. This can be written thus: Hb4 + 4O2 Hb4O8 The above reaction however occurs in stages, with one O2 molecule combining with Hb at a time. The sequence of the reaction is as follows: Hb4 + O2 Hb4O2 Hb4O2 + O2 Hb4O4 Hb4O4 + O2 Hb4O6 Hb4O6 + O2 Hb4O8 The above reaction is very rapid. It requires less than 0.01 second. The deoxygenation of Hb 4O8 is also rapid. The combination of oxygen with hemoglobin is a self- catalytic reaction. The formation of Hb4O2 is relatively slow, but Hb4O2 catalyzes the combination of the next O2 molecule with Hb4O2 so that Hb4O4 is formed at a faster rate than Hb4O2 and so on. This is why the oxygen- hemoglobin dissociation curve has a steep rise. The PO2 of pulmonary capillary blood when fully oxygenated is 100mmHg, i.e. equal to alveolar PO2. Before the blood gets to the left atrium, it mixes with venous blood from the bronchial vein (physiological shunting) so that the PO2 of the blood entering the left atrium is about 97mmHg. 22 NSC 203 HUMAN PHYSIOLOGY (II) Figure 6: Transport of oxygen in the blood 3.4 Oxygen dissociation curve The oxygen dissociation curve is a graph that shows the percent saturation of hemoglobin at various partial pressures of oxygen (Figure 1-50). The oxygen–haemoglobin dissociation curve plots the proportion of haemoglobin in its saturated form on the vertical axis against the prevailing oxygen tension on the horizontal axis. The oxyhemoglobin dissociation curve is an important tool for understanding how blood carries and releases oxygen. Specifically, the oxyhemoglobin dissociation curve relates oxygen saturation (SO2) and partial pressure of oxygen in the blood (PO2), and is determined by what is called "hemoglobin’s affinity for oxygen"; that is, how readily haemoglobin acquires and releases oxygen molecules into the fluid that surrounds it. The oxygen- hemoglobin dissociation curve can shift to the left or shift to the right. The factors that can cause a shift to the right are: (a) increase in temperature (b) increase PCO2 (c) increased acidity (fall in pH) (d) increase in 2,3 diphosphoglycerate (2,3 DPG) A shift to the right due to increased acidity is called Bohr’s effect. The factors that can cause a shift to the left are the opposite of those that cause shift to the right. These are: (a) decrease in temperature (b) decrease PCO2 23 NSC 203 HUMAN PHYSIOLOGY (II) (c) reduced acidity (increase in pH) (d) decrease in 2,3- DPG (e) presence of foetal hemoglobin Variation of the hydrogen ion concentration This changes the blood's pH. A decrease in pH shifts the standard curve to the right, while an increase shifts it to the left. This is known as the Bohr’s effect. A reduction in the total binding capacity of haemoglobin to oxygen (i.e. shifting the curve down, not just to the right) due to reduced pH is called the root effect. Effects of carbon dioxide Carbon dioxide affects the curve in two ways: first, it influences intracellular pH (the Bohr’s effect), and second, CO2 accumulation causes carbamino compounds to be generated through chemical interactions, which bind to haemoglobin forming carbaminohaemoglobin. Low levels of carbamino compounds have the effect of shifting the curve to the right, while higher levels cause a leftward shift. However, this isn't the overriding effect of CO2 accumulation. Only about 5–10% of the total CO2 content of blood is transported as carbamino compounds. Most of the CO2 content (80–90%) is transported as bicarbonate ions. The formation of a bicarbonate ion will release a proton into the plasma. Hence, the elevated CO2 content creates a respiratory acidosis and shifts the oxygen–haemoglobin dissociation curve to the right. Effects of 2,3-D.P.G. 2,3-Disphosphoglycerate or 2,3-DPG is an organophosphate, which is created in erythrocytes during glycolysis. The production of 2,3-DPG is likely an important adaptive mechanism, because the production increases for several conditions in the presence of diminished peripheral tissue O2 availability, such as hypoxaemia, chronic lung disease, anaemia, and congestive heart failure, among others. High levels of 2,3-DPG shift the curve to the right, while low levels of 2,3-DPG cause a leftward shift, seen in states such as septic shock and hypophosphatemia. Temperature Temperature does not have such a dramatic effect compared to the previous factors, but hyperthermia causes a rightward shift, while hypothermia causes a leftward shift. Carbon monoxide Haemoglobin binds with carbon monoxide 200-250 times more readily than with oxygen. The presence of carbon monoxide on one of the 4 haem sites causes the oxygen on the other haem sites to bind with greater affinity. This makes it difficult for the haemoglobin to release oxygen to the tissues and has the effect of shifting the curve to the left (as well as downward, due to direct competitive effects of carbon monoxide). With an increased level of carbon monoxide, a person can suffer from severe tissue hypoxia while maintaining a normal PO2. 24 NSC 203 HUMAN PHYSIOLOGY (II) Figure 7: Oxygen-haemoglobin dissociation curve 3.5 Carbon-dioxide transport Since cellular (internal) respiration is a continuous process, carbondioxide is produced continuously in the body tissues. This CO2 must be transported from the tissues to the lungs where it is expired. The transport of CO2 involves: (a) diffusion of CO2 from the tissues into the blood (b) transport of CO2 in the blood to the lungs (c) diffusion of CO2 from the pulmonary capillaries into the alveoli (d) movement of CO2 from the alveoli into atmospheric air The PCO2 in the tissues is 46mmHg and PCO2 in the arterial blood is 40mmHg. Carbondioxide is about 20 times more diffusible than oxygen at body temperature. So, CO2 diffuses rapidly from the tissues along its concentration gradient into the blood. Transport of CO2 in blood Carbondioxide is transported in the blood in three forms (a) as dissolved CO2 -10% (b) in combination with plasma protein and haemoglobin- carbamino compound- 30% (c) as bicarbonate- 60% CO2 in solution Since CO2 is very soluble, about 10% of the CO2 transported is in solution in the plasma. 25 NSC 203 HUMAN PHYSIOLOGY (II) CO2 as Carbamino-compounds Carbondioxide combines with the free NH2 groups in plasma proteins and free NH2 groups in haemoglobin inside the red blood cell to from carbamino compounds. Thus, CO2 + protein-NH2 = protein – NHCOOH= protein-NHCOO- + H+ The H+ produced in this reaction is buffered by plasma proteins and phosphates. Similarly, CO2 + Hb - NH2 = Hb- NHCOOH= HbNHCOO- + H+ The H+ produced is buffered by haemoglobin and phosphate esters in the red cells. CO2 as bicarbonate In the plasma CO2 combines with water to form carbonic acid. CO2 + H2O H2CO3 H+ + HCO3The reaction is very slow unless the enzyme carbonic anhydrase which acts as a catalyst is present. Carbonic anhydrase is not found in the plasma, but it is abundant in the red cells. Thus, when CO2 diffuses into red blood cells, it is very rapidly converted to bicarbonate and hydrogen ions. Carbonic anhydrase increases the speed of this reaction about 5000 times compared with that in the plasma. Because of the high rate of its formation, the concentration of bicarbonate in the red blood cell becomes higher than that in the plasma. Consequently, HCO3diffuses from the red cell into the plasma. Since the H+ produced by the same reaction has been buffered by intracellular haemoglobin and phosphate, H+ cannot move out with the HCO3- . Also, neither Na+ nor K+ can move of the cell to accompany the HCO3 so as to maintain electrical equilibrium. This is because the Na+- K+ ATPase pump maintains strictly the intra and extra- cellular Na+ and K+ concentrations. Chloride is the only free anion that can move. So, chloride moves from the plasma into the red cell to replace the HCO3 that has moved out so as to maintain electrical neutrality. This movement of chloride into the red cells is called chloride shift or the Hamburger effect. As a result of this chloride shift, the concentration of chloride in venous blood is about 2% lower than in arterial blood. Also water moves into RBC so that red blood cell in venous blood is about 3% bigger than in arterial blood. 26 NSC 203 HUMAN PHYSIOLOGY (II) Figure 8: Movement of gases at alveolar level. Figure 9: Movement of gases at tissue level. 27 NSC 203 HUMAN PHYSIOLOGY (II) Carbon-dioxide dissociate curve The volumes of CO2 in 100 volumes of blood is plotted against the PCO2 (Figure 1-53). A separate CO2 dissociation curve is usually drawn from reduced whole blood and oxygenated whole blood. Carbon-dioxide in the three forms is carried in the venous blood (where the PCO2 is 46mmHg, same as the in the tissues because of equilibration) to the right side of the heart and from there to the lungs. In the lungs, there is a reversal of the reactions at the tissue level so that CO2 at a partial pressure of 46mmHg in the pulmonary arterial capillary diffuses into the alveoli where the PCO2 is 40mmHg. From the alveoli, the CO2 is expelled to the atmosphere during expiration. Factors such as rate of tissue metabolism (which determines rate of CO 2 production), rate of blood flow, degree of deoxygenation of haemoglobin at the tissues level and the rate/ depth of pulmonary ventilation will affect the rate of CO2 transport. Figure 10: Carbon-dioxide dissociation curve 3.6 Ventilation – Perfusion relationship Ventilation is the movement of air into and out of the lungs; while perfusion is the flow of blood through the lungs. In the normal lung, in upright position, distribution of ventilation is greatest to the top and decreases slightly towards the bottom of the lung, while in a similar position, distribution of perfusion is greatest at the bottom and decreases markedly towards the top of the lung. For exchange of gases to occur normally in the lungs, a normal ratio of ventilation to perfusion must be maintained. Ventilation/perfusion imbalance is said to exist when this ratio is abnormal. An extreme example is a situation where a main bronchus is totally blocked by a foreign body that has been accidentally inhaled. In this situation, although there is an adequate blood flow to the affected lung, because there is no ventilation of the lung, there is no exchange of gases in that lung. Also, there could be obstruction to blood flow to a whole lung due to clot embolism. Although there will be adequate ventilation of the affected lung, because there is no perfusion of 28 NSC 203 HUMAN PHYSIOLOGY (II) lung, no exchange of gases takes place in that lung. Very often, there are small areas in the lungs that may be underperfused. Such areas constitute physiologic dead spaces and increase the total dead space of the lung. 5.0 Summary In this unit, you have leant about the process of Gaseous transport and exchange with emphasis on how the body receives oxygen and gives out carbon dioxide applying physical and chemical principles of pressure gradients and combining with haem and supported by some factors. 6.0 6.1 6.2 Tutor Marked Assignments Activity: As provided by the facilitator and the practical workbook. Please answer the following questions: i. Explain the process of gaseous exchange. ii. Explain the 4 steps of the Oxygen transport process iii. Describe the Oxygen-haemogobin combination and how this occurs in the vessels and the heart using appropriate equations. 7.0 References and other resources Fox, S.I. (2012). Human Physiology. 12th edition, Mc Graw Hill, New York. Ganong W.F. (2010). Review of Medical Physiology. 23rd edition, Mc Graw Hill, New York. Guyton, A.C& Hall J.E. (2001). Textbook of Medical Physiology. Harcourt International Edition, 10th edition, W.B. Saunders, Philadelphia. Oyebola, D.O. (2002). Essential Physiology, Vol 1, Nihort Press UNIT 4: CONTROL OF RESPIRATION AND RESPIRATORY ADAPTATIONS IN UNUSUAL ENVIRONMENT CONTENT 1.0 Introduction 2.0 Objectives 3.0 Main Content 3.1 Neural control of breathing 3.2 Chemical control of breathing 3.3 Respiration at High Altitude 3.4 Deep Sea Diving 4.0 Conclusion 5.0 Summary 6.0 Tutor Marked Assignments 6.1 Activity 6.2 Tutor Marked Tests 7.0 References and other resources 29 NSC 203 HUMAN PHYSIOLOGY (II) 1.0 Introduction Normal respiration has a rhythmic pattern, with inspiration alternating with expiration. It is unusual to find a normal person breathing in all the time without inspiring and expiring. The normal rhythmic pattern of breathing is possible because of precise control of breathing. The control of breathing can be divided into two broad groups, neural (nervous) control and chemical control. In this unit you will learn about control of breathing in usual and unusual environment. 2.0 Objectives At the end of this unit, you should be able to: i. Explain the process of neural control of breathing ii. Explain the process of the chemical control of breathing iii. Describe different groups of chemo-receptors iv. Explain the ventilatory responses to changes in pH, carbon dioxide and oxygen lack. 3.0 Main Content 3.1 Neural control of breathing Neural control is the influence of nerve cells of the central nervous system and their efferent and afferent fibres on the regulation of breathing. The neural control can be sub-divided into two main types: i. voluntary control; ii. involuntary or autonomic control Voluntary control Voluntary control of respiration occurs when an individual decides to hold his breath or to hyperventilate. It is well known that when a swimmer wants to dive into the deep end of the swimming pool, he first takes a deep breath, dives into the water and holds his breath for the period he is under the water. If he breaths-in while under the pool, he will aspirate water into his lungs and may drown. Athletes are trained to consciously breathe in and out deeply before the start of a race. The acts of breath-holding and voluntary hyperventilation constitute voluntary control of breathing. It is carried out under the control of the motor cortex. Involuntary control of breathing The involuntary control of breathing is through the activities of collection of neurons referred as centres in the pons and the medulla and stretch receptors in the lung. The brain stem centres and the peripheral stretch receptors in the lungs are connected by afferent and efferent fibres into a network of functionally inter-related and well-connected neuronal circuit. In the medulla oblongata, there are two centres, the inspiratory centre and the expiratory centres. In the pons, there are also two centres. In the lower part of the pons is the apneustic centre, while the pneumotaxic centre is located in the upper part of the pons. These brain stem respiratory centres are connected to each other and to the inspiratory muscles. The inspiratory neurons (I neurons) at the inspiratory centre are capable of spontaneous discharge of nervous impulses. This impulse is transmitted in the spinal cord to the anterior horn cells of C3, 4 and 5 and efferent fibres that emerge from there is the phrenic nerve which supplies the diaphragm. Also, some of the impulses go to the anterior horn cells in the thoracic region and the efferent from these innervate the intercostal muscles. Discharge of impulses from 30 NSC 203 HUMAN PHYSIOLOGY (II) the inspiratory centre causes inspiration to occur, and this leads to expansion of the lungs. During inspiration, I-neurons inhibit the expiratory neurons (E-neurons) of the expiratory centre. The depth of inspiration produced by the activity of the I-neurons is shallow. The apneustic centre in the lower pons send facilitatory impulses to the inspiratory centre and this leads to an increase in the depth of inspiration. At the same time as this increase in depth of respiration is going on, the following events are occurring (i) Facilitatory impulses pass from the inspiratory centre to the pneumotaxic centre, causing its stimulation (ii) Expansion of the lungs following inspiration causes the stretch receptors in the lungs to be stimulated. Afferent impulses from the stretch receptors are transmitted through the vagus nerve to the apneustic centre and cause its inhibition. (iii) The pneumotaxic centre sends inhibitory impulses to the apneustic centre. (iv) At the same time, the pneumotaxic centre sends facilitatory impulses to the expiratory centre. (v) The inhibitory impulses reaching the apneustic centre from the pneumotaxic centre cause a reduction in the inspiratory drive of the apneustic centre on the inspiratory centre. (vi) Also, the inhibitory impulses from the I-neurons to the E-neurons are removed. (vii) As a result of inhibition of I-neurons and the stimulation of E-neurons, expiration occurs. Figure 11: Respiratory neurons in the brain stem When expiration has occurred, the inhibitory impulses from the lungs on the apneustic centre are removed. The inhibition of the I-neurons during expiration removes the facilitatory impulses from the I-neurons to the pneumotaxic centre, which in turn stops inhibiting the apneustic centre. The apneustic centre is free once more to drive the inspiratory centre and the cycle starts all over again. This is how the alternating phases of inspiration and expiration occur. 31 NSC 203 HUMAN PHYSIOLOGY (II) Figure 12: The regulation of ventilation by the central nervous system. 3.2 Chemical control of breathing Some chemical components of blood can affect respiration. The three main substances involved are CO2, H+ and O2. A rise in PCO2 or H+ concentration of arterial blood or a fall in its PO2 increases the activity of the respiratory centre, while a decrease in PCO2 or H+ concentration have inhibitory effect on respiratory centre activity. Increase in PO2 has no effect on respiratory centre. The changes in the chemical composition of blood are detected by chemoreceptors. There are two groups of chemoreceptors: i. Peripheral chemoreceptors ii. Central chemoreceptors Peripheral chemoreceptors The carotid and the aortic bodies are the peripheral chemoreceptors. The carotid body is located near the bifurcation of the common carotid artery into internal and external carotid arteries. There are usually two or more aortic bodies near the arch of the aorta. The carotid body has the glossopharyngeal nerve as its afferent fibre, while the vagus nerve in the afferent fibre from the aortic body. The carotid bodies have a very high blood flow, about 2000 ml/100g of tissue per minute. Because of this large blood flow, the oxygen needs of the carotid bodies can be met by dissolved oxygen in plasma. For this reason, situations like anaemia or carbon monoxide poisoning which reduce the oxygen- carrying power of the blood do not cause stimulation of the carotid chemoreceptors. The peripheral chemoreceptors respond to decrease in PO2 or increase in H+. 32 NSC 203 HUMAN PHYSIOLOGY (II) Figure 13: Respiratory control by in the carotid and aortic bodies Central chemoreceptors The central chemoreceptors are located on the floor of the 4th ventricle in the medulla oblongata. They respond to changes in the H+ concentration of the cerebrospinal fluid (CSF) or possibly, brain interstitial tissue. Hydrogen ion cannot penetrate the blood brain barrier easily; but CO2 does. The CO2 that enters the brain and CSF is hydrated to form H2CO3. The H2CO3 dissociates to H+ and HCO3. Therefore, the H+ concentration in the CSF rises and this stimulates the central chemoreceptors. Chemoreceptor regulation of breathing in response to changes in PCO2 is illustrated in Figure 3-13. Ventilatory responses to changes in pH When there is an increase in the concentration of H+ ions in the body, as may occur in uncontrolled diabetes mellitus which results in accumulation of acid ketone bodies, there is marked stimulation of respiration, mainly through the central chemoreceptors. The increased ventilation resulting from this leads to increased expulsion of CO2 from the body and this leads to a compensatory fall in blood H+ concentration. Conversely, when the H+ concentration in blood falls, there is no stimulation of the central chemoreceptors, leading to a decreased ventilation and a consequent rise in arterial PCO2. This leads to a rise in H+ concentration. Thus, the blood concentration of H+ is important in the regulation of respiration. Persistent hyperventilation that is not secondary to an increase in H+ concentration leads to a fall in blood H+ concentration (due to excessive washing out of CO2). This is respiratory alkalosis. 33 NSC 203 HUMAN PHYSIOLOGY (II) Also, hypoventilation that is not secondary to a fall in plasma H+ concentration leads to increase in H+ concentration (due to accumulation of CO2). This is respiratory acidosis. Ventilatory responses to CO2 When the arterial PCO2 is increased, this stimulates the peripheral chemoreceptors (and indirectly, through increased H+ concentration the central chemoreceptors are also stimulated) leading to increased pulmonary ventilation. The increased ventilation results in increased ―washing out‖ of CO2 from the body so that the PCO2 is reduced. If the arterial PCO2 falls too low, the CO2 drive on the chemoreceptors is reduced or stopped. This leads to reduced excretion of CO2. This allows CO2 to accumulate in the body and causes a rise in PCO2. The operation of this feed-back mechanism of respiratory control keeps CO2 excretion and production in balance. The normal arterial PCO2 is 40 mmHg. Moderate increases in this concentration causes increased ventilation. If the arterial CO2 level becomes too high, rather than stimulate respiration, it leads to depression of the central nervous system, including the respiratory centre, and it also produces headache, confusion and eventually coma. This is CO2 narcosis. Ventilatory responses to oxygen lack When the arterial PO2 is reduced, this causes stimulation of the peripheral chemoreceptors. The normal arterial PO2 is 97 mm Hg and slight decreases in arterial PO2 causes a slight stimulation of the chemoreceptors. This slight reduction in PO2 does not usually lead to an increase in ventilation for two reasons. The first is that reduced arterial PO2 means that haemoglobin is less saturated with oxygen. HbO2 is more acidic than Hb. Therefore, less HbO2 means that the blood will become slightly more alkaline and this will tend to inhibit respiration. This tends to cancel the stimulatory effect of the reduced PO2. Secondly, any increase in ventilation that may occur lowers PCO2 and this tends to inhibit respiration. Because of the two reasons above, the effect of fall in arterial PO2 is not obvious until the O2 falls to 60 mmHg or less. Under the latter condition, the rate and depth of respiration is increased. When PO2 comes back to normal, this stimulus due to oxygen lack is removed and respiration comes back to normal. The above account shows the importance of the three chemical substances, H+, PCO2 and PO2, in the chemical control of respiration. 34 NSC 203 HUMAN PHYSIOLOGY (II) Figure 14: Chemoreceptor control of breathing 3.3 Respiration at High Altitude The higher one goes into a high altitude, the lower the amount of air available in the environment. For example, the PO2 in inspired air at sea-level is 159 mm Hg and alveolar PO2 is 104 mm Hg, while at an altitude of about 10,000 meters, PO2 in inspired air is 47 mm Hg and alveolar PO2 is 21 mm Hg. There is a corresponding decrease in the total barometric pressure. The hypoxia at such an altitude stimulates the peripheral chemoreceptors in the carotid body leading to an increase in pulmonary ventilation. The resulting hyperventilation leads to a lot of CO2 being expelled from the body and this produces respiratory alkalosis. In order to be able to survive at such an altitude the body makes some adjustments, referred to as acclimatization. Acclimatization to high altitude includes a great increase in pulmonary ventilation, an increase in red blood cell count brought about by stimulation of the bone marrow by erythropoietin, increase in 2, 3–DPG level which increases oxygen delivery to the tissues, excretion of alkaline urine to 35 NSC 203 HUMAN PHYSIOLOGY (II) correct the alkalosis, an increase in the number of mitochondria in the cells and increased vascularity of the tissues. Figure 15: Respiratory adaptations to a high altitude Cyanosis This is a bluish discoloration of the skin and mucous membrane due to presence of a large quantity of deoxygenated haemoglobin in the blood. Blueness is not often easy to observe in the skin of black Africans except those who are very fair in complexion and in neonates. However, cyanosis can be observed in the tongue, nail bed and buccal mucosa of black Africans. Usually, cyanosis becomes noticeable when the arterial blood contains 5g or more of deoxygenated haemoglobin per 100ml (dl) of blood. 3.4 Deep sea diving When human beings descend beneath the sea, the pressure around them increases tremendously. For every 10 meters of depth in sea-water, pressure on the diver increases by 1 atmosphere. So, at a depth of 31m (100ft) in the ocean, a diver is exposed to a pressure of 4 atmospheres (normal atmospheric pressure + 3 atmospheres due to 31m depth of sea water). In order to prevent the lungs from collapsing, the air breathed by the diver must be supplied under high pressure that is hyperbaric air and the condition of breathing air under high pressure is called hyperbarism. The 36 NSC 203 HUMAN PHYSIOLOGY (II) gases present in the air the diver normally breathes are nitrogen, oxygen and carbon-dioxide. When these gases are breathed under high pressure, especially nitrogen and oxygen, serious physiological effects can result. CO2 content of inspired air is very low and if there is no rebreathing, CO2 does not create problems when breathed under high pressure. As the diver descends deeper into the sea, the increased pressure to which he is subjected causes compression of the gases being inspired, leading to a decrease in volume and an increase in pressure according to Boyle’s law. The increased pressure causes a lot of nitrogen and oxygen to dissolve in the body fluids and in the tissues. As the depth of descent increases, the quantities of dissolved nitrogen and oxygen increase. Nitrogen narcosis At sea level, nitrogen has no effect on body functions, but nitrogen breathed at high pressures can cause varying degree of narcosis. When a diver remains beneath the sea for one hour or more, at about 120ft, the first symptoms of mild nitrogen narcosis appears. The diver becomes unduly jovial and loses many of his cares. At 150 to 200 feet, he becomes drowsy and at 200 to 250 feet, he becomes very weak and becomes too clumsy to perform the work he is supposed to do. At depths greater than 250 feet, the diver becomes almost useless as he can no longer perform any function. The features of nitrogen narcosis are similar to those of alcoholic intoxication, hence it is often called ―raptures of the depths‖. The mechanism of the narcotic effect of nitrogen is believed to be similar to that of anaesthetic gases. Decompression sickness (also called bends, Caisson disease, Diver’s paralysis). If a diver stays beneath the sea for a long time, a large quantity of nitrogen will dissolve in his body. If the diver suddenly comes back to the surface of the sea, large quantities of the nitrogen that has been forced into solution will come out of solution and form gas bubbles in the body fluids, both in the intracellular and extracellular fluid compartments. Those bubbles that are in the plasma will flow along with the blood and block some of the small blood vessels (air embolism). These air bubbles can cause damage to any part of the body. The extent of the damage and the associated symptoms depend on the number and sizes of bubbles formed. The latter gives rise to ―decompression sickness‖. The symptoms of decompression sickness are due largely to the resultant air embolism. This leads to tissue ischaemia and sometimes, tissue death. About 90% of people suffering from decompression sickness develop pain in the joints and muscles of the legs or arms. The joint pain is the reason this condition is also called ―bends‖. Various degrees of disorders of the nervous system such as dizziness, paralysis, collapse and unconsciousness may develop. A small percentage may develop massive pulmonary microembolism, with associated shortness of breath and later, pulmonary oedema. This can lead to death. Decompression sickness can be avoided if the diver is made to ascend to the surface of the sea gradually over a period of 2 to 5 hours. If a diver is brought slowly to the surface, the dissolved nitrogen is exhaled rapidly enough through the lungs to prevent decompression. Slow ascents over one hour will elimination of about 70% of the dissolved nitrogen, while about 90% will have been eliminated if ascent is carried out in 6 hours. Another method used to prevent decompression sickness is to bring the diver to the surface quickly and put him in pressurized tank and then lower the pressure inside the tank gradually 37 NSC 203 HUMAN PHYSIOLOGY (II) back to normal atmospheric pressure over a couple of hours. Pressurized tank can also be used to treat the diver in whom the symptoms of decompression sickness have occurred. The diver is recompressed in the tank and then slowly decompressed over several hours. The use of helium instead of nitrogen in the air mixture breathed by divers engaged in deep dives and stay underwater for long periods has minimized the problem of decompression sickness. The introduction of SCUBA apparatus (self-contained underwater breathing apparatus) has turned diving into a popular sport. However, because of the limited amount of air a diver can carry in his SCUBA gear, the diver cannot remain for a long time underwater, otherwise, he will run out of air. 4.0 Conclusion Breathing has voluntary, neural and chemical control with appropriate ventilatory responses to oxygen, carbon dioxide concentrations. Respiratory adaptations also occur appropriately in high altitude and in deep sea diving. 5.0 Summary In this unit, you have leant about the following voluntary and autonomic control of respiration. You also learn about peripheral and central chemoreceptors regulation and ventilatory responses to pH, oxygen lack and carbon dioxide. Th i. Neural control of breathing ii. Chemical control of breathing 6.0 6.1 Tutor Marked Assignments Activity: Conduct the experiments on (1) respiratory movements and effects of various factors (2) measuring lung volumes and capacities 6.2 Answer the following questions: i. Explain the process of neural control of breathing. ii. Explain the process of Chemical control of breathing iii. Describe the chemoreceptors iv. How do the body respond to changes in pH, carbon dioxide and oxygen lack?. 7.0 References and other resources Fox, S.I. (2012). Human Physiology. 12th edition, Mc Graw Hill, New York. Ganong W.F. (2010). Review of Medical Physiology. 23rd edition, Mc Graw Hill, New York. Guyton, A.C& Hall J.E. (2001). Textbook of Medical Physiology. Harcourt International Edition, 10th edition, W.B. Saunders, Philadelphia. Oyebola, D.O. (2002). Essential Physiology, Vol 1, Nihort Press 38 NSC 203 HUMAN PHYSIOLOGY (II) MODULE 4: GASTROINTESTINAL PHYSIOLOGY Introduction Beyond air exchange, the body depends on nutrients that should be taken, digested and absorbed to supply energy and other micronutrients needed. This module covers the organization of the gastrointestinal system, the secretions, the hormones and the processes of digestion and absorbtion. Module Objectives At the end of this module, you should be able to: 1. Discuss the organization of the gastro-intestinal system 2. Discuss the secretions of the various organs of the gastro-intestinal tracts. 3. Describe the hormones and their actions along the GIT 4. Discuss the process of digestion and absorption along the GIT CONTENT Unit One Unit Two Unit Three UNIT ONETRACT - Organization and Secretions of the Gastrointestinal Tract Gastrointestinal Motility Gastrointestinal Hormones, Digestion And Absorption ORGANIZATION AND SECRETIONS OF THE GASTROINTESTINAL CONTENT 1.0 2.0 3.0 3.1 3.2 3.3 3.4 4.0 5.0 6.0 6.1 6.2 7.0 Introduction Objectives Main Contents Organization of the gastrointestinal tract The sphincters Characteristics of sphincters Functions of sphincters Conclusion Summary Tutor Marked Assignments Activity Tutor Marked Tests References and other resources 1.0 Introduction The gastrointestinal system performs digestive functions that provide the nutrients needed for energy and other organic functions. The GIT system is organized with different organs that also allow for control release of contents through sphincters. In this unit, you will learn about the organization and the functions of some of the organs in the GIT. 39 NSC 203 HUMAN PHYSIOLOGY (II) 2.0 Objective At the end of this unit, you should be able to: i. Explain the Organization of the gastrointestinal tract GIT ii. Describe the sphincters in the GIT iii. Describe the salivary glands and the secretions. iv. Describe other secretions (pancreatic, intestinal, gall bladder) of the GIT and their functions 3.0 Main Content 3.1 Organization of the gastrointestinal tract The gastrointestinal system include alimentary canal, extending from pharynx to anus and accessory organs like salivary gland, liver and pancreas. Figure 1 shows the entire alimentary tract. Figure 1: The Alimentary tract 40 NSC 203 HUMAN PHYSIOLOGY (II) Organizational structure The digestive tract includes the mouth, pharynx, esophagus, stomach, small intestine, large intestine, rectum and anus. The histology is similar in all part except the esophagus and the anus where serous attachment is not present. Figure 1.2 shows a typical cross section of the intestinal wall. The basic structures include: (i) serous coat (ii) muscular layer – longitudinal and circular layer (iii) sub-mucosa (iv) Mucosa Figure 2: Typical cross section of the gut Innervation The wall of gastrointestinal tract (GIT) has intrinsic nervous system (enteric nervous system) beginning from the esophagus extending down the anus. It is composed mainly of two plexus. i. The outer one lies between the longitudinal and circular layer and is called Auerbach’s or Myenteric plexus. ii. submucosa plexus or Meissner’s plexus lies in the submucosa layer. The myenteric plexus control GIT movement. It is sensitive to stretch while the submucosal or meissner’s control secretion and local blood flow. It is sensitive to osmolar changes, pH changes and chemical composition of food. The enteric nervous system can function on its own, independently of the extrinsic nerves supply to the gut. The extrinsic innervation can modify the activity of the enteric nervous system. The extrinsic supply comes from the autonomic nerves. The sympathetic fibres gives relaxation of smooth muscle and vasocontraction of smooth muscle, vasodilation and secretion of the digestive juice. 3.2 The sphincters The alimentary tract is divided into functional compartment by sphincter. They include: i. Upper esophageal sphincter 41 NSC 203 ii. iii. iv. v. vi. vii. HUMAN PHYSIOLOGY (II) Lower esophageal sphincter Pyloric Sphincter of Oddi Ileocecal sphincter Internal sphincter (smooth muscle/involuntary) Outer sphincter (skeletal muscle/voluntary) Characteristics of sphincters i. They have high tension/pressure area. The tensions within the sphincter are very high. ii. The resting tone is greater than the other two adjacent segments which result in intraluminal high pressure zone that separate the two lumen to compartments. iii. It relaxes in response to appropriate stimulus, so that flow may occur from one compartment to the next. iv. It regulates or maintains aurocaudal (mouth to anus) flow of GIT contents Functions of sphincters i. The upper esophageal sphincter prevent air into the esophagus during respiration. ii. The lower esophageal sphincter prevents irritant gastric from refluxing into the esophagus iii. The pyloric sphincter separates the acidic environment of the stomach from the alkaline environment of the duodenum. iv. The ileocecal sphincter separates ileum from the cecum, disallowing the faeces in the cecum from refluxing back into the ileum. v. The sphincter of Oddi allows intermittent flow of pancreatic secretion and bile. vi. The internal sphincter has smooth muscle and it shows involuntary movement of defecation, while the outer anal sphincter has skeletal muscle and shows voluntary movement. 3.3 Salivary glands and secretions The digestive secretion in the mouth comes from salivary gland. There are three major salivary glands, as shown in Figure 2.1. i. Parotid (behind the tongue) ii. Submaxillary (Submandibular) iii. Sublingual 42 NSC 203 HUMAN PHYSIOLOGY (II) Figure 3: Formation and secretion of saliva by a submandibular salivary gland They are exocrine glands. Each has acini and duct cells. In the acini, two types of secreting cells have being found: i. Serous secretion containing an alpha-amylase; this is the enzyme for the digestion of starches. ii. A mucous secretion, containing mucin, which is important for lubrication purposes There is also lingual lipase, secreted by the Ebner’s glands. The parotid glands produce entirely the serous type of secretion; the submandibular glands secrete both the serous and mucous types and the sublingual produce only the mucous type of secretion. The submaxillary produces 10% secretion, sublingual produced 5% secretion and parotid produced 25%. Innervation The salivary glands are supplied by both sympathetic and parasympathetic nerves. The sympathetic action gives vasoconstriction which causes secretion to be viscous and scanty. The parasympathetic effect causes vasodilation and causes copious secretion. The parasympathetic fiber of cranial nerve-7 supplies some submaxillary and sublingual and cranial nerve-9 supplies the parotid gland. The parasympathetic is mediated via the release of acetylcholine that can be blocked by atropine. Composition of saliva i. The secretion of saliva per day ranges from 1000 ml- 1500ml. ii. It has a pH of 6.8 which range from 6.7- 7. iii. It contains 99.5% water and 0.5% solid The solid consists of organic and organic substances. The major inorganic constituent are: Na+, K+, Cl-, HCO3-, Ca++ and Mg2+. The organic substances include the enzyme- α- amylase, mucin, lysozymes, IgA, blood group antigens, urea, uric acid, etc. 43 NSC 203 HUMAN PHYSIOLOGY (II) Duct cells function The saliva in the duct is isotonic while the saliva in the mouth is hypotonic. This is due to the change that takes place in the lumen of the ducts. The duct-epithelial cell show active reabsorption of sodium ion in exchange for potassium ion. There is also reabsorption of Cl- in exchange for HCO3- since the duct cells are impermeable to water, the removal of sodium and chloride ions makes the saliva to be hypotonic. Aldosterone which is a mineralocorticoid acts on salivary duct to cause sodium ion reabsorption in exchange for potassium ion. The concentration of Na+ and K+ in saliva depends on flow rate. At high flow rate, less time is allowed for transfer of ions and hence Na+ is more than K+ but the saliva still remains isotonic at high flow rate. Regulation of secretion Salivary secretion is regulated mainly by neural mechanisms. It shows both conditional and unconditional reflexes. Conditional reflex is established by learning, and the secretion can be seen from sight, smell and thought of food. Unconditional reflex comes from presence of food in the mouth. The secretion of saliva in both sides of reflexes is caused by activity of parasympathetic nerves (VII) and (IX) supplying the glands. Secretion is almost abolished (reduced) during sleep. There is a decrease in resting flow rate when there is dehydration, anxiety, fear and severe mental effort. Functions of the saliva i. The enzyme α- amylase acts on boiled starch and convert it to maltose. The beginning of carbohydrate digestion occurs in the mouth. ii. The mucin present in the saliva lubricate the food which helps in mastication iii. Saliva is necessary for swallowing iv. Helps in taste perception of food materials by dissolving them v. It facilitates speech. Speech is difficult in dry mouth vi. Lysozymes and IgA present in the saliva gives birth to bactericidal and immunity functions respectively. vii. It neutralizes the gastric acid that refluxes into the esophagus and release heart burn. 3.4 Gastric secretion Gastric juice is secreted by gastric glands present in the gastric mucosa of fundus and body of the stomach. There are 3 types of cells namely: i. Neck and surface epithelial cell that secrete mucous ii. The chief cell that secretes enzyme iii. Parietal or Oxyntic cells that secrete Hydrochloric acid (HCl) and intrinsic factors. Composition of gastric secretion The volume of secretion per day is about 2 liters and the pH varies between 1.8 – 2.0. The inorganic constituents include Na+, Cl-, PO4-, K+ and SO4-. The organic substances present in the secretion are digestive enzyme (pepsinogen, renin, lipase), mucin and intrinsic factors. 44 NSC 203 HUMAN PHYSIOLOGY (II) Function of HCl (Hydrochloric acid) The concentrated Hcl in the gastric juice is necessary to activate pepsinogen to pepsin, the extreme acidity is bactericidal. The acid pH in the upper part of duodenum facilitates ion absorption. Function of gastric juice The beginning of protein digestion takes place in the stomach. Pepsin acts on protein and convert it to peptone. The enzyme is secreted by chief cell as active pepsinogen. Gastric renin Is a milk curdling enzyme which is absent in human but present in cows. Gastric lipase: Is a weak fat-splitting enzyme. Intrinsic factor Is secreted by the parietal cell of the fundus. It is required for the absorption of vitamin B 12 (cyanocobalamin or extrinsic factor). The absorption of vitamin B12 occurs in the terminal ileum. Mucus Is secreted by surface epithelial cell and neck cell of the gland. The surface epithelial cell also secrete bicarbonates, the mucus and the bicarbonate form gel in the lining of the gastric mucosa. This gel protects the mucosa from the action of the acid. In fact, there is a pH gradient from the lumen to the mucosal wall of the stomach. The pH in the mucosal is 7 and that of the lumen is 2. The presence of HCO3- and mucus form the acid mucosa barrier. Certain condition like chronic stress, alcohol and aspirin has the tendency to arose the acid mucosa barrier. Hydrochloric acid secretion Hydrochloric acid (HCl) is the gastric juice secreted from the parietal cells. The secretion of Hcl is an active process and it is transported against the electrochemical gradient. The concentration of Hcl in gastric juice is 150 mEq/L whereas it is 00004 mEq/L in plasma. The source of H+ is from the dissociation of H2CO3. The H2CO3 is formed from hydration of CO2 in the presence of carbonic anhydrase enzyme. Carbonic acid dissociates to H+ and HCO3-. The hydrogen ion formed is exchanged with potassium ion and the transport carrier is H+- K+ ATPase. The energy released from the breakdown of ATP is utilized for the active transport of H+. Chloride ion is also actively transported from the cell into the lumen and coupled with H+ to form Hcl. The active transport of Hcl is followed by passive transport of water into the lumen. The secretion of H+ leaves bicarbonate ion within the cell. The HCO3- combines with Na+ and enters the blood as NaHCO3. It is known during digestion, alkaline level in the blood and urine rise and it is called Postprandial alkaline tide. Agents causing secretion of Hydrochloric acid i. Gastrin is released from pyloric antrum ii. Acetylcholine is secreted from the vagal ending iii. Histamine comes from enterochromaffin cell lining the mucosa 45 NSC 203 HUMAN PHYSIOLOGY (II) When vagus is activated it releases acetylcholine. This stimulates the release of GRP (Gastric Releasing Peptide) followed by secretion of gastrin. Vagal stimulation also causes release of histamine. All these cause release of Hcl. When there is peptic ulcer, treatment could be induced by inhibiting acid secretion, and this could be achieved from the following i. Inhibiting H2 receptors by cimetidine ii. Blocking hydrogen ion – potassium ion ATPase by omeprazole. Regulation of gastric secretion Gastric juice secretion is regulated by both neural and hormonal mechanisms. Neural regulation is mediated via the vagus. Acetylcholine is released by vagal ending. The acetylcholine activates phospholipase C which in turn raises intracellular Ca2+. The intracellular Ca2+ promotes the secretion of gastric juice. Hormonal regulation is by gastrin which is secreted from the pyloric antrum. 3.4.1 Phases of gastric secretion There are 3 phases of gastric secretion: (1) Cephalic phase (2) Gastric phase (3) Intestinal phase Cephalic phase Conditional reflexes like sight, smell and thought of food causes secretion of gastric juice. Presence of food in the mouth also causes secretion in the stomach. The cephalic phase occur by activity of the vagus. Shaming-feeding experiments in animals like dog gives an example of cephalic secretion. The quantity of juice secreted is small. The cephalic phase accounts for about 10% of the total secretion associated with a meal. Gastric phase The gastric phase accounts for about 80% of the total secretion of gastric juice. This phase is initiated by the presence of food in the stomach. The food stretches the stomach wall and this distension stimulates the gastric glands via both the extrinsic (vagal) and intrinsic (nerve plexuses) reflex pathways to produce gastric juice. Distension of the pyloric antrum also results in the release of gastrin into the blood by an intrinsic reflex. In addition, some substances in the food, known as secretagogues, elicit release of gastrin by the intrinsic reflex. Such substances include meat extracts, protein digestion products, alcohol, bile acids and caffeine. During this phase, maximum secretion occurs. Intestinal phase The arrival of food and product of digestion in the intestine also stimulates gastric secretion. The quantity produced is very small. However, the presence of food in the duodenum inhibit secretion of gastric juice. This inhibition is mediated through enterogastric reflex. The presence of acid and fat in the duodenum causes release of secretin and cholecystokinin. Also, there is release of VIP, GIP- all of which are enterogastrones. i.e. they cause inhibition of gastric secretion. 46 NSC 203 HUMAN PHYSIOLOGY (II) 3.3 Pancreatic secretion The pancreas has both exocrine and endocrine functions. The digestive enzymes are secreted from exocrine pancreas. The exocrine pancreas consists of acini and ducts. Composition About 1500 ml of pancreatic juice are secreted daily. Pancreatic juice is a watery alkaline fluid; isotonic with plasma and rich in digestive enzymes. The alkalinity is due to HCO3- secreted from the duct epithelial cells. Other inorganic substances are Na+, K+, Cl- and SO4-. The organic constituents includes: peptidase, amylase, lipase, nuclease. Peptidases It includes: Trypsin and Chymotrypsin Trysinogen enterokinase Trypsin Chymotrysinogen Trypsin Other peptidases include carboxypeptidase and elastase Nuclease RNA and DNA Nucleotide Lipase Phospholipase Triglycerides Fatty acids Regulation of pancreatic secretion The regulation is mainly from hormones. The 2 types of hormone regulating 2 types of secretion. i. Secretin is produced by the duodenal mucosa. Presence of acid chyme in the duodenum is the main stimulus resulting in secretion of watery fluid which in bicarbonate that helps to neutralize the acid pH. ii. Cholecystokinin (CCK) causes secretion of the pancreatic juice which in digestive enzymes. It also acts on the gut bladder. Stimulus is product of food digestion entering the duodenum. 3.5 Bile Bile is secreted by the liver cells (liver lobules) and stored in the gallbladder. Secretion of bile The volume secreted per day is 500-700 ml, and secretion occurs when chyme enters the duodenum. PH is 7.6-7.8. Composition of bile Bile released from the gall bladder into the duodenum has the following composition. Water - 92% Bile Salts - 6% Bilirubin - 0.3% Cholesterol – 0.3% Fatty acids - 0.3% Lecithin – 0.3% Other lipids – 0.2% Na+ - 130mEq/L 47 NSC 203 HUMAN PHYSIOLOGY (II) K+ - 12mEq/L Ca2+ - 23mEq/L Cl- - 25mEq/L HCO3- - 10mEq/L Bile salts They are secretory products of the liver. They are formed from cholesterol which gives rise to primary bile acid which becomes conjugated into Na+ and K+ to form taurocholate and glucocholate. Bile pigments The breakdown of haemoglobin gives rise to bilirubin and biliverdin which are bile pigment. They are excretory products of the liver. The golden yellow of the bile is due to the presence of the pigment. Phospholipids Lecithin is the chief phospholipids present in the bile. It is insoluble in water. It becomes solubilized in micelles. Cholesterol Cholesterol in bile is solubilized in micelles. The presence of bile salt keeps cholesterol in solution and prevents its precipitation to form stones. Functions of the bile Digestion and absorption of fats and fat soluble vitamins (ADEK) depends on presence of bile. The fat is lipid soluble while the digestive enzyme lipase is enzyme soluble. In order to facilitate the action of lipase on lipids, bile shows the following effects: i. The bile produces emulsification of fat. By this, large molecules are broken down into smaller ones. ii. They show hydrotropic effects. This action of bile enables lipase enzyme to digest the fat. iii. Bile reduces surface tension. This effect facilitates the lipase enzyme action. iv. The micelles that is formed after digestion promotes absorption. The micelles consist of digested glycerides combined with bile. Enterohepatic circulation The bile salts that are secreted into the duodenum are reabsorbed and recirculated. About 90% of the bile salt that enter the small intestine are absorbed from the terminal ileum, and enter the liver through the portal circulation. From the liver, it is recirculated into the duodenum; this forms enterohepatic circulation. The bile salts in circulating pool are only 3-6gm but the quantity required for digestion is 4—8 gram. Normal digestion and absorption of fat is able to take place by recirculation. The total circulating pool circulate twice during digestion of each meal. The rate of synthesis depends on the rate of returns to the liver. About 0.2 gm/day of bile is lost in faeces. Any condition that affects enterohepatic circulation and decrease bile pool and causes mal absorption of bile and fat soluble vitamins may result in steatorrhea. 48 NSC 203 HUMAN PHYSIOLOGY (II) Regulation of bile Bile secretion is regulated by hormones. There are 2 mechanisms: i. Hormone secretin produced from duodenum when acid chyme enters acts on the biliary duct cells to increase secretion of water and electrolytes which helps to neutralize the acid chyme, and this action is known as hydrocholeretic effects. a. The bile salt in the bile causes stimulation of the liver to secrete more bile. This action is called choleretic effect. ii. Hormone cholecystokinin (CCK) also acts on gall bladder smooth muscle causing contraction and expulsion of bile. This action of CCK on gall bladder is called chologugue effect. Gall bladder Gall bladder stores, releases and makes the bile into concentration by active reabsorption of Na+ and HCO3-, passively by the reabsorption of water. Gall stones There are 2 types of gall stone- cholesterol stone and pigment stone. Normally, cholesterol and lecithin are found in solution by the formation of micelles. When there is alteration in concentration, cholesterol crystallizes to form stones. Cholesterol stones are radiolucent. Pigment stones are formed due to infection or obstruction of the biliary tree. The conjugation of bilirubin makes it insoluble resulting in precipitation. Pigment stones are radiopaque. Intestinal secretion The small intestine consists of duodenum, jejunum, and ileum. The small intestinal secretion is mainly water, mucus and electrolytes. It is alkaline in nature and ranges between 10001500ml/day. Digestive enzymes are not secreted into the lumen, they are present in the apical surface of the villi. At the base of the villi are glands called Crypts of Lieberkuhn which contains cells that secrete mucus. The intestinal gland of the duodenum are called Brunner’s gland which also secrete mucus. The mucus secreted by the Brunner’s gland provides protection to the mucosa lining against mechanical damage and also lubricate the mucosa lining. Regulation of intestinal secretion Regulation is neural and hormonal. Vagal stimulation during digestion causes secretion of intestinal hormones. Vasoactive intestinal peptide (VIP) hormones increase secretion of intestinal glands. The most important is the local enteric nervous system. 5.0 Summary In this unit, you have leant about the organization of the gastrointestinal tract and the various sphincters with different characteristics and functions. You have also learnt that the salivary glands of different types produce secretions that also perform digestive functions. Gastric, pancreatic, gall bladder secretions also perform different functions. 6.0 6.1 6.2 i. Tutor Marked Assignments Activity: - See Laboratory manual and the experiments on gastrointestinal system Please answer the following questions: Describe the organization of the gastrointestinal tract 49 NSC 203 ii. iii. iv. v. vi. 7.0 HUMAN PHYSIOLOGY (II) Describe the sphincters in the GIT Explain what gastric secretion is and list its major composition Describe the three phases of gastric secretion Discuss the pancreatic secretion and its major composition Describe the composition and functions of the bile. References and other resources Fox, S.I. (2012). Human Physiology. 12th edition, Mc Graw Hill, New York. Ganong W.F. (2010). Review of Medical Physiology. 23rd edition, Mc Graw Hill, New York. Guyton, A.C& Hall J.E. (2001). Textbook of Medical Physiology. Harcourt International Edition, 10th edition, W.B. Saunders, Philadelphia. Oyebola, D.O. (2002). Essential Physiology, Vol 1, Nihort Press UNIT TWO - GASTROINTESTINAL MOTILITY CONTENT 1.0 Introduction 2.0 Objectives 3.0 Main Contents 3.1 Gastrointestinal motility 3.2 Chewing 3.3 Deglutition (Swallowing) 3.4 Gastric motility 3.5 Motility of the small intestine 3.6 Defeacation 4.0 Conclusion 5.0 Summary 6.0 Tutor Marked Assignments 6.1 Activity 6.2 Tutor Marked Tests 7.0 References and other resources 1.0 Introduction The physical activity of processing the food items from the point of entry to the point of eliminating the waist takes a course in gastrointestinal motility. This unit traces these actions from the mouth to the anus. 2.0 Objective At the end of this unit, you should be able to: i. Explain the term, Gastrointestinal motility ii. Describe the act and use of Mastication (Chewing) iii. Describe the process of Swallowing (Deglutition) iv. List the functions of the stomach v. Describe the act of Gastro-intestinal motility vi. Describe the process of Defeacation 50 NSC 203 HUMAN PHYSIOLOGY (II) 3.0 Main Content 3.1 Gastrointestinal motility The main function of the alimentary tract is to ingest, digest and absorb food materials. For this to be carried out, appropriate mixing must be provided. The swallowing mechanism is shown in Figure 4.1. The mixing/propulsion are different at each compartmental stage of the tract. It requires both neural and hormonal mechanisms. i. Mastication (Chewing) ii. Swallowing (Deglutition) iii. Gastric motility iv. Motility of the small intestine v. Motility of the colon vi. Defeacation (passage of faeces) 3.2 Chewing Chewing is the process by which food brought into the mouth is broken down into smaller pieces by the teeth. In the process, the food is mixed with saliva. Chewing makes it easier to swallow the food and contributes to its enjoyment by homogenizing and mixing the food with saliva, thus releasing taste-producing substances. Chewing can be carried out voluntarily, but is more frequently a reflex activity. By subdividing the food into smaller particles, chewing makes it possible for the food to mix more readily with digestive secretions of the stomach and duodenum. The tongue and the cheek muscles are used to keep the food mass between the teeth during mastication. 3.3 Deglutition (Swallowing) It refers to the act of swallowing. It consists of oral, pharyngeal and esophageal stages. The 1 st stage is voluntary, the 2nd and 3rd stages are involuntary and reflex in nature. Oral stage It is voluntary. The food is masticated by mixing with saliva. The solid food is converted to soft bolus and positioned in the dorsum of the tongue. In this stage, the bolus passes through the oral cavity toward the pharynx assisted by the tongue pressing against the hard palate. Pharyngeal stage The presence of food at the entry of the pharynx stimulates receptors in the tonsil and epiglottiswhich initiate the reflex. This stage is involuntary: the efferent and afferent impulses are carried by cranial nerves V, IX, X and XII. The center is the swallowing center located in the medulla and the lower pons. This reflex allows the bolus to enter the esophagus and not the trachea. The entry of the bolus into the nasopharynx is prevented by: i. The soft palate raises and press against the posterior pharyngeal wall. ii. There is an upward and forward movement of the larynx which causes the glottis approximately with the epiglottis and seal with the larynx. The vocal cord also approximately inhibiting speech. 51 NSC 203 HUMAN PHYSIOLOGY (II) iii. There is ceasation of respiration at this stage. The bolus is therefore directed at the esophagus. Esophageal stage The bolus enters the esophagus from the pharynx through the upper esophageal sphincter and enters the stomach through the lower esophageal sphincter. Entry of food bolus causes distension and relaxation of upper esophageal sphincter. This is as a result of inhibition of the vagus nerve. This distension initiate a wave of peristalsis which spread along the length of esophagus pushing the bolus forward. These are: primary peristalsis wave travelling at 3-4cm/sec (frequency). The force of gravity helps the wave of movement. Liquid travels faster than solid to reach the stomach. When the bolus reaches the lower esophageal sphincter, it relaxes and allows the bolus to enter the stomach, after which it closes to prevent regurgitation into the esophagus; this action is regulated by the myenteric plexus which secretes VIP or NO (nitric oxide). If the primary peristalsis does not completely empty the esophagus, one or more secondary peristalsis arises from distal part of esophagus. Figure 4: Swallowing mechanism Disorders of swallowing 1. Dysphagia - difficulty in swallowing 2. Achalasia – difficulty in emptying the food from the esophagus to stomach due to absence of peristalsis in the lower 3rd or failure of cardiac sphincter to relax. 52 NSC 203 HUMAN PHYSIOLOGY (II) 3.4 Gastric motility Functions of the stomach i. Storage organ: stomach can store large quantity of food, as it shows receptive relaxation. ii. Beginning of protein digestion by pepsin occurs in the stomach. iii. It makes food into chyme by propulsive, mixing and retropulsive movement. iv. The acid in the stomach has bactericidal effect. The gastric secretion has intrinsic factor which is necessary for absorption of vitamin B12. v. The acid in the stomach convert cane sugar to fructose and glucose. The acid in chyme in the duodenum facilitates iron absorption. Figure 5: Physiologic anatomy of the stomach Movement in the stomach Receptive relaxation The stomach shows receptive relaxation, accommodating large volume of food. The receptors for this are present in the wall of pharynx and the stomach. The food is stored in the fundus and the body. The afferent and efferent are the vagus. It causes the plexus to secrete VIP. Vagotomy does not completely abolish reflective relaxation, it only decreases it. Digestive peristalsis (Mixing of food) The distal part of the stomach shows peristalsis. The distension of the wall of the distal part (antrum) stimulates intrinsic plexus. The smooth muscle first shows slow waves which is not propagatory, and it’s called basal electrical rhythm (BER). The membrane potential reaches the threshold level of pharynx. The entry of Na+ and Ca2+ causes depolarization, resulting in spike potential (action potential) which is propagatory and forms peristalsis. Peristalsis consists of wave of contraction followed by relaxation frequency of which is 3/min. 53 NSC 203 HUMAN PHYSIOLOGY (II) When the food reaches the pyloric sphincter, it retropul back to the antrum because the pyloric sphincter is closed. The propulsion, mixing and retropulsion in the pyloric breaks down the food to chyme and helps through mixing by the gastric juice. Each time the peristalsis arises at the sphincter, only 2-3 ml is empty to the duodenum. Gastric emptying Gastric emptying occurs mainly due to gastroduodenal plexus cycle caused by digestive peristalsis of the stomach. The transit time for the gastric empty of the food is 3-4 hrs. As peristalsis reaches the sphincter, the chyme is retropuls allowing only small quantity (2-3 ml) to emptied into the duodenum and closes back. Factors influencing emptying Liquid leaves the stomach earlier than solid. Carbohydrate empty faster than protein and fat is emptied last. Increase in acid pH inhibit emptying. Isotonic leaves earlier than hypo or hypertonic fluids. Enterogastric reflex Presence of fats, acid or hyper osmolar solution in the duodenum inhibit gastric emptying. Other duodenal factors include: i. The degree of distension of duodenum ii. Presence of any degree of irritant in the duodenal mucosa iii. Degree of acidity in the duodenal chyme iv. Degree of osmolarity of the chyme The inhibition is mediated by neural and hormonal mechanisms. The neural involves the inhibition of the vagus while hormonal mechanism includes release of secretin, VIP, CCK and GIP; these hormones inhibit gastric motility. 3.5 1. 2. 3. 4. Motility of the small intestine The movement that are seen in small intestine are as follows (Figure 4-3): Segmental contraction Peristalsis contraction Pendular contraction Villi movement Segmental contraction The frequently occurring movement in the small intestine is the segmental contraction. Slow waves develop in the circular smooth muscle of the wall due to stimulation of the plexus. When the slow wave reaches threshold, segmental contraction that is propagatory develops. The frequency of the slow wave is highest in the duodenum (12/min) and lowest in the ileum (8/min). This facilitates the bolus to be propelled aborally. The segmental contraction involves ring-like regular constriction along the length of the segment of the intestine. The constricted part latter relaxes and the relaxed part constrict. This process is repeated over and over again resulting in the bolus moving back and forth within the lumen. Functions of segmental peristalsis are as follows: i. Bolus mixes well with digestive enzymes and facilitate completion of digestion 54 NSC 203 HUMAN PHYSIOLOGY (II) ii. The segmental contraction causes exposure of digested food to the villi surface for absorption iii. The occurrence of segmental contraction in the proximal segment and inhibition in the distal segment facilitate propulsion of bolus toward the segment colon. Peristalsis contraction Sometimes the longitudinal muscle contraction gives pendular contraction which facilitates mixing of bolus with digestive enzymes. Figure 6: Pattern of gastrointestinal motility and propulsion Villi movement There is contraction of the smooth muscle of the villi that results into forward and backward movement. The hormone- villi kinin stimulates the movement which facilitates absorption of digested food. Gastroileal reflex Distension of the stomach by food causes relaxation of ileocecal sphincter and allows emptying of the ileal content into the caecum. Distension of the ileum will also cause the relaxation of the sphincter and emptying. On the other hand, the distension of the caecum will result in contraction of the caeca and prevent reflux of the caecal content into the ileum. This is facilitated by the ileocecal valve. The activity of the ileocecal sphincter is controlled by the myenteric plexus. Functions of the small intestine i. It contains pancreatic bile and intestinal secretion ii. Completion of digestion and absorption of digested food occurs in the small intestine iii. Presence of villi facilitates absorption of digested food iv. The duodenal mucosa secretes gastrointestinal hormones like secretin, CCK, VIP, motilin. etc. v. Payer’s patches in the ileum are lymphoid organs which helps in immunity. 55 NSC 203 HUMAN PHYSIOLOGY (II) Migrating motor complex It is an interdigesting peristalsis occurring in the stomach and small intestine. It occurs in between meal at every 70-90 mins interval. Each peristalsis last 10mins. It is developed due to activity of intrinsic myenteric system. It helps in sweeping the content of the stomach and small intestine towards colon toward interdigestive period. Motilin is the hormone responsible. The large intestine It includes the caecum, ascending colon, transverse colon and descending or sigmoid colon, rectum and anus. The mucosa secretes mucus but there are no villi, hence no absorption of food, no digestive enzymes. However, it has important functions: i. Absorption of water and electrolytes ii. Formation of faeces iii. Secretion of mucus to lubricate the faeces iv. The bacteria fluoride synthesis vit B and vit K. Motility of the large intestine i. The large intestine shows haustral shutting (segmental contraction) ii. Mass peristalsis (contraction) iii. Peristalsis Haustral shutting It is similar to segmental contraction of the small intestine. In the large intestine, the longitudinal muscle forming 3 bands taenia coli. The enteric plexus below this is greater and the region adjacent has a thin wall. It gives rise to sac-like pouches along the length of segment of the colon that are called haustrau. The back and forth movement causes the chyme to be exposed for absorption of water and electrolytes. Out of 1,500ml of the mixed chyme that enters the colon per day, all will be absorbed leaving 50-100ml in the faeces. The haustral shutting also facilitate propulsive movement of the faeces to the distal colon. Transit time in the colon is very slow (5-10cm/hr). Mass peristalsis It occurs in a large segment of the colon, the contraction is powerful enough to cause the colon to be in contracted state for a long period. It is stimulated by myenteric plexus. The main function is to sweep the faeces along the segment of the colon. The mass contraction occurs 3-5 times per day and usually lead to defeacation. 56 NSC 203 HUMAN PHYSIOLOGY (II) Figure 7: The human colon 3.6 Defeacation Defeacation is the act of passing faeces. It is a complex behavior involving both reflex and voluntary actions. The rectum is normally empty. When faeces are pushed into the rectum by mass movements, the urge to defeacate is felt. The anal sphincters however prevent escape of faeces unless the individual is prepared for defeacation. There are two anal sphincters, the internal and the external sphincters. The internal sphincters consist of a circular smooth muscle that is in the anal wall, while the external sphincter consists of striated voluntary muscle that surrounds the internal sphincter and also extends distal to it. The internal sphincter is supplied by parasympathetic nerves and the external sphincter, which is under voluntary control, is supplied by somatic nerves. The process of defeacation can be subdivided into main components: i. The part under the defeacation reflexes ii. The part under voluntary control The defeacation reflexes There are two types of defeacation reflexes i. The intrinsic defeacation reflex ii. The parasympathetic defeacation reflex The intrinsic defeacation reflex is mediated via the myenteric plexuses. When faeces enter the rectum, distension of the rectal wall initiate peristaltic waves via a local reflex circuit and these 57 NSC 203 HUMAN PHYSIOLOGY (II) peristaltic waves spread to the descending colon, sigmoid colon and rectum forcing faeces towards the anus. As the peristaltic waves approach the anus, the internal and external relaxes. If the external sphincter is also relaxed, defeacation will occur. The peristaltic waves produced by the intrinsic defeacation reflex is usually weak and may not be effective in causing defeacation. This weak contraction is often reinforced by contractions mediated by the parasympathetic defeacation reflex, which involves parasympathetic nerves in the sacral segment of the spinal cord. These parasympathetic impulses augment the ineffectual weak movements produced by the intrinsic defeacation reflex so that they become powerful and effective in emptying the bowel. In spite of the two reflexes above, defeacation can only occur if the circumstance is socially acceptable for the act. The ability not to defeacate in some circumstances is due to the fact that the conscious mind takes over voluntary control of the external sphincter. Relaxation of the internal sphincter and forward movement of the faeces towards the anus normally cause an instantaneous contraction of the external sphincter. Impulses from the cerebral cortex which pass through the somatic nerves to the external sphincter will either inhibit the sphincter to allow defeacation to occur or further contract it if the circumstance is not conducive to defeacation. When the circumstances is right for defeacation to occur, the defeacation reflex is followed by relaxation of the external anal sphincter. Intra-abdominal pressure is elevated to aid in the expulsion of faeces. Evacuation is normally preceded by a deep breath, so that the diaphragm descends towards the abdominal cavity. The glottis is closed and contraction of the respiratory muscles on full lungs raises both the intrathoracic and intra-abdominal pressure. Contraction of the muscles of the wall of the abdomen causes a further increase in intra-abdominal pressure. The additional pressure generated by this bearing down effort as well as the strong contractions of the defeacation reflex helps to force faeces out of the anus through the relaxed sphincters. Figure 8: Afferent and efferent pathways of the parasympathetic mechanism for enhancing the defeacation reflex 58 NSC 203 HUMAN PHYSIOLOGY (II) 5.0 Summary In this unit, you have leant about gastrointestinal motility, the acts of mastication, swallowing and movement of the bulk though the tract. 6.0 6.1 6.2 i. ii. iii. iv. v. 7.0 Tutor Marked Assignments Activity: See the Laboratory manual and directives from the Facilitator Please answer the following questions: Describe the act and use of Mastication (Chewing) Describe the process of Swallowing (Deglutition) List the functions of the stomach Describe the act of Gastro-intestinal motility Describe the process of Defeacation References and other resources Fox, S.I. (2012). Human Physiology. 12th edition, Mc Graw Hill, New York. Ganong W.F. (2010). Review of Medical Physiology. 23rd edition, Mc Graw Hill, New York. Guyton, A.C& Hall J.E. (2001). Textbook of Medical Physiology. Harcourt International Edition, 10th edition, W.B. Saunders, Philadelphia. Oyebola, D.O. (2002). Essential Physiology, Vol 1, Nihort Press UNIT THREE - GASTROINTESTINAL HORMONES, DIGESTION AND ABSORPTION CONTENT 1.0 Introduction 2.0 Objectives 3.0 Main Contents 3.1 Gastrointestinal hormones 3.2 Digestion 3.3 Absorption 4.0 Conclusion 5.0 Summary 6.0 Tutor Marked Assignments 6.1 Activity 6.2 Tutor Marked Tests 7.0 References and other resources 1.0 Introduction Gastrointestinal hormones are chemical agents secreted by the endocrine glands released into general circulation and acts on the alimentary tract. They include the following: (a) gastrin (b) secretin (c) cholecystokinin- pancreozymin (CCK-P2) (d) gastric-inhibitory peptide (GIP) (e) vasoactive intestinal peptide (VIP) (f) motilin (g) gastric releasing peptide (GRP). Importantly, hormones help with digestion. The food which the body needs can be classified into carbohydrate, proteins and fats. This cannot be absorbed in their natural forms through the 59 NSC 203 HUMAN PHYSIOLOGY (II) gastrointestinal tract. Before they can be made available for body use, they must be broken down into smaller molecules which can be absorbed. This process of breaking down is known as digestion. It is the end product of digestion that is now absorbed. The basic process in the digestion of all food types is called hydrolysis. This unit will improve your knowledge on gastric hormones, digestion and absrbtion. 2.0 Objective At the end of this unit, you should be able to: i.Describe the gastric hormones ii.Explain the process of digestion in human iii.Discuss the process of digestion of proteins and fats iv.Discuss the absorption processes of carbohydrate, fat, proteins and vitamins. 3.0 Main Content 3.1 Gastric Hormones 3.1.1 Gastrin Gastrin is produced by G-cells of the pyloric antral mucosa. Some amounts are also secreted by duodenal mucosa. Gastrin secreting tumors are being found in the pancreases. There are three forms of gastrin which are being isolated. We have G-17-17 amino acid residue G-34-34 amino acid residue G-14-14 amino acid residue G-17 is the most biological active form, the half-life between G-14 and G-17 is 2-3mins in circulation while G-34 has 15mins as its half-life. Action of gastrin i. It stimulates gastric acid and pepsin secretion (more than 1000 times more potent than histamine) ii. It stimulates growth of gastric mucosa iii. It stimulates insulin and glucagon secretion after a protein meal but not after carbohydrate iv. It stimulates the contraction of cardiac sphincter v. It stimulates gastric motility Regulation of gastrin i. Distension of the stomach ii. Presence of protein and product of protein breakdown iii. Stimulation of parasympathetic supply, pyloric action and acetylcholine secretion iv. Blood borne factor like Ca2+ and epinephrine v. Presence of acid in the stomach has a negative feedback control which inhibits gastrin secretion vi. Substances like secretin, GIP, VIP, glucagon, and calcitonin inhibits gastrin secretion 3.1.2 CCK – P2 (cholecystokinin – pancreozymin) It is found in the nerves in many parts of the body, including distal ileum, colon, and brain. It is secreted in the mucosa of the upper intestine (duodenum). 60 NSC 203 HUMAN PHYSIOLOGY (II) Porcine CCK exists in various form, e.g. 58, 39, 33, 12, 8 and 4. Half –life is 5 mins. Action of CCK- P2 i. It stimulates secretion of pancreatic juice, rich in enzyme or rich in digestive juice. ii. It augment action of secretin in producing secretion of an alkaline pancreatic juice iii. It inhibits gastric emptying iv. It exerts a trophic effect on the pancreas stimulating the growth of the exocrine cells v. It increases the secretion of interleukins vi. It enhances motility of small intestine and colon vii. It stimulates the contraction of the gall bladder and the relaxation of the sphincter of Oddi viii. Along with secretin, it augments contraction of pyloric sphincter thus preventing the reflux of duodenal content into the stomach ix. It stimulates the glucagon secretion 3.1.3 Secretin Is stimulated by the entry of the product of food digestion into the duodenum, e.g. amino acid, protein and fat. There is a positive feedback regulation of secretin by presence of food. Action of Secretin i. It increases the secretion of HCO3- by pancreatic duct and biliary tract ii. It promotes the secretion of watery alkali pancreatic juice iii. It augment the action of CCK in producing pancreatic juice rich in digestive enzymes iv. It decreases gastric acid secretion v. It causes contraction of the pyloric sphincter vi. It increases insulin secretion Regulation of secretin i. Secretion of secretin is stimulated by acid chyme bathing the duodenum ii. Alkali secretion helps to neutralized the acidity iii. Products of protein digestion is also a stimulus 3.1.4 Gastric inhibitory peptide i. Gastric inhibitory peptides is produced in the K-cell in the mucosal of duodenum and jejunum. It consists of 43 amino acids. In large doses, it causes inhibition of gastric secretion and motility. In smaller doses, it does not show this action. ii. It stimulates insulin secretion and forms one of the important B-cells stimulating hormones. Regulation of gastric inhibitory peptide 1. Secretion of GIP is caused by present of glucose and fats in the duodenum. 3.1.5 i. ii. iii. iv. Vasoactive intestinal peptide (VIP) It has 28 amino acids It is found in the nerves of GI tract, autonomic nerves and brain It acts as a co-transmitter with acetylcholine Half-life is 2mins in circulation Action of vasoactive intestinal peptide 61 NSC 203 i. ii. iii. iv. v. HUMAN PHYSIOLOGY (II) It stimulates intestinal secretion of electrolytes and water It relaxes intestinal smooth muscles including the sphincter It causes dilatation of peripheral blood vessel It inhibits gastric acid secretion It potentiate action of acetylcholine of salivary gland. Enterogastrones They are hormones which causes inhibition of gastric secretion and motility. They are secreted in the duodenum in response to the presence of acid chyme in the lumen. They include secretin, GIP and VIP. 3.1.6 Motilin Is secreted from the duodenal mucosal. It consists of 22 amino acid. The concentration in the blood undergoes cyclic fluctuation during fasting. The peak blood level corresponds to the beginning of the activity of myoelectric motor complex (mmc). Action of motilin i. It stimulates gastric acid secretion ii. It causes contraction of the gall bladder to increase bile concentration. iii. It prepares the intestine for the next meal. Other hormones include somatostatin, glucagon, gastric releasing peptide (GRP), neurotensin and substance P. 3.2 Digestion Digestion of carbohydrate Almost all carbohydrate of the diet are large polysaccharide or disaccharide, which are combinations of monosaccharides bound together by the process of condensation. Hydrogen ion (H+) has been removed from one of the monosaccharide while a hydroxyl ion (OH-) has been removed from the next one. The two monosaccharides combines with each other at this site of removal and the H + and OHcombine to form water (H2O). The dietary carbohydrate is mainly; Amylopectin, which consists of chains of glucose molecules joined by 1,4 - α linkages, with some branches linked by 1,6 – α linkages. For digestion of carbohydrate to take place the specific ion returns the H and OH ions to the polysaccharide and thereby separate the monosaccharides from each other and this process is called hydrolysis. R1 R11 + H2O R1H + R11OH Three major sources of carbohydrate exists in the normal human diet and these are : sucrose; the disaccharide known as cane sugar, lactose; the disaccharide in milk and starches, large polysaccharides present in almost all non-animals foods, particularly grains and tubers. When food is chewed, it is mixed with saliva, which contains the enzymes ptyalin and this is an α-amylase from the parotid gland. This enzyme hydrolyzes starch into the disaccharide maltose and isomaltose. An isomaltose is small polymers of glucose containing 3 to 9 glucose molecules. Because the food remains in the mouth for only a short time, about 3 to 5% of all the starches eaten becomes hydrolyzed. Even though the food does not remain in the mouth long enough for ptyalin to complete the breakdown of starches into maltose, its actions can continue as long as an hour after the food has entered the stomach i.e. until the content of the fundus mix with the 62 NSC 203 HUMAN PHYSIOLOGY (II) stomach secretions. Then, the activities of the α-amylase is blocked by the acid of gastric secretions. Before food properly mix with this gastric secretions, about 30-40% of the starches would have been hydrolyzed into maltose. Pancreatic secretions contains a large quantity of α-amylase and immediately after the chyme empties from the stomach into the duodenum and mixes with pancreatic juice, starches that have not already been split and digested amyloses are converted before they pass the jejunum. The epithelial cells contains four enzymes: lactase, sucrase, maltase, and isomaltose, which are capable of splitting disaccharides: lactose, sucrose, maltose and isomaltose into their constituents monosaccharides. The enzymes are located in the brush border of the cells lining the lumen of the intestine. Lactose splits into a molecule of glucose and a molecule of galactose. Sucrose splits into a molecule of glucose and fructose. Maltose splits into 2 molecules of glucose and isomaltose into several molecules of glucose. Since the ordinary diet contains far more starches than sucrose and lactose. Glucose represents about 80% of the final product of carbohydrate, while galactose and fructose represents about 10% each. The digestion of carbohydrates is summarized in the diagram below Starches Maltose & Isomaltose Ptyalin (Saliva) – 20 to 40% Pancreatic amylase – 50 to 80% Lactose Maltase & Isomaltase (Intestine) Glucose Sucrose Lactase (Intestine) Galactose Sucrase Fructose Figure 9: Digestion of carbohydrates Digestion of fat The most common fat of the diet are the neutral fat known as triglyceride. Each molecule is composed of the glycerol nucleus and three fatty acids. Also present in the normal diet are small molecules of phospholipids, cholesterol and cholesterol esters, which are regarded as fats. Phospholipids and cholesterol esters contains fatty acids, while cholesterol have some of the physical and chemical properties of fats , but have no fatty acids. The Ebner’s glands on the dorsal surface of the tongue secrete lingual lipase and the stomach also secrete a gastric lipase. The gastric lipase does not play any important role in fat digestion, but the lingual lipase is active in the stomach and can digest as much as 30% of the triglycerides in the diet. 63 NSC 203 HUMAN PHYSIOLOGY (II) Essentially, all fat digestion occurs in the small intestine. The most important enzyme being the pancreatic lipase in the pancreatic juice. Minute quantities of lipase (enteric lipase) from the epithelial cells of small intestine also act to cause hydrolysis of fat. The first step in fat digestion is to breakdown fat globules to small sizes, so that water soluble digestive enzymes can act on the globule surfaces. This process is called emulsification of fat and this is achieved under the influence of bile (secretion of liver that does not contain any digestive enzymes). Bile contain large quantity of bile salt mainly in the form of ionized sodium salt that are very important for emulsification of fat. The carbonyl and other polar parts of the bile salt molecules are highly soluble in H2O, whereas most of the sterol portion of the bile salt are highly soluble in fat. The fat soluble portion of the bile salt dissolves in the surface layer of fat lobule with the polar portion of the salt projecting outward and soluble in the surround fluids. This effect greatly decreases the interfacial tension of the fat. The property of this bile acids is multiplied several times by the lecithin that are also in the bile. When the interfacial tension of the globule of non-miscible fluid is low, this fluid on agitation can be broken up into many minute particles. After emulsification, the following reaction occurs: i. Lipase hydrolyzes triglycerides to release two monoglycerides and two free fatty acids. ii. Esterase hydrolysis two monoglycerides to glycerol and free fatty acids. iii. Phospholipase, the main enzyme for hydrolysis of phospholipids to lysolecithin and free fatty acids Digestion of proteins Dietary proteins are derived entirely from meat and vegetables. Proteins are formed from long chains of amino acids bind together by peptide linkages. Digestion of proteins, unlike carbohydrate start in the stomach. Pepsin is capable of digesting essentially all the different types of proteins in diet. One of its important feature is to digest collagen and albuminoid that is affected little by other digestive enzyme. Collagen is a major constituent of intercellular connective tissue of meat. Pepsin, however only begins the process of protein digestion by providing as much as 10-30% of the total protein digestion. Most protein digestion occurs in the small intestine under the influence of a proteolytic enzymes of the pancreatic secretion. When protein leaves the stomach, they are in form of proteases, peptones and large polypeptides. On entering the small intestine, they are trapped by the pancreatic enzymes, trypsin, chymotrypsin and carboxypolypeptidase. Both trypsin and chymotrypsin transmit protein molecules into small polypeptides. Carboxypolypeptidase then cleaves individual amino acids from the carboxyl ends of the polypeptides. The brush border of small intestine contains several enzymes for hydrolyzing the final linkages of remaining dipeptides and other small polypeptides. These enzymes are; aminopolypeptidase and dipeptidase. 64 NSC 203 HUMAN PHYSIOLOGY (II) The digestion of proteins can be summarized thus: Protein Proteoses Peptones Polypeptidases Trypsin, Chymotrypsin, Carboxypolypeptidase Peptidase Polypeptides + Amino acids Amino acids Figure 10: Digestion of proteins 3.2 Absorption The end product of the digestion of different types of food ingested or secreted electrolytes, vitamins and large quantity of water secreted in various digestive juices must be moved from the lumen of the gut across the epithelium to the interstitial fluid. The process of transport from gut lumen into body’s interstitial fluid is called absorption. The main absorptive portion of the gut is the small intestine with a large surface area. The surface area is achieved by the following: i. The mucosal infolding, called the valvulae corniventis. ii. Billions of small villi projecting about 1mm from the surface of the mucosa iii. A brush border consisting of about 600 villi per cell The combination of the three above increase the absorptive area of the mucosal about 600 folds. The total area of small intestine is 250m2. Absorption occurs basically by active transport and diffusion. Absorption of carbohydrate Glucose, other hexoses and pentoses are rapidly absorbed across the wall of small intestine. These sugars are absorbed before the content of the small intestine reach the terminal part of the ileum. The transport is an active process, and this can be demonstrated by several important experimental observation and they are: i. Transport can be blocked by metabolic inhibitors such as IAA, cyanite and phlorhizin ii. Transport is selective, the order of preferences for transporting difference monosaccharides and their relative rate of transport in comparison with glucose are; galcatose-1.1, glucose-1.0, fructose-0.4, mannose-1.2, xylose-0.15, arabinose-0.1. iii. There is competition between certain sugars for the respective carrier system Glucose and galactose in small intestine enter the cells by secondary active transport with sodium ion. Transport of other hexoses are affected by the amount of Na ion in interstitial lumen. Glucose and Na share the same co-transporter or symport called Na-dependent glucose transporter (SGLT). There are two members of this family; SGLT 1 and SGLT 2. Since the intracellular concentration of Na ions is low in interstitial cell, Na diffuses into the cell along its concentration gradient. Glucose moves with the Na and is released in the cell. The Na ion is transported into the lateral intracellular spaces and the glucose from the inside of the cell by another transporter; the SGLT 2 into the interstitium and finally into the capillaries. 65 NSC 203 HUMAN PHYSIOLOGY (II) Absorption of glucose is called a secondary active transport because the energy for transport is provided indirectly by the active transport of Na out of the cell. Fructose, however, utilizes a different mechanism its absorption is independent of Na ions or transport of glucose and galactose. It is transported by facilitated diffusion from the intestinal lumen into the cells of the intestine by a transporter GLUT 5 and out of the cell into the interstitium by another transporter GLUT 2. Some fructose is converted to glucose in the mucosal cells. Pentoses are absorbed by simple diffusion. All monosaccharides are absorbed into the portal blood draining the small intestine. Absorption of lipids (fats) The digestive end product of fat dissolve in the lipid portion of bile acid micelles. These micelles are soluble in the chyme. In this form, the monoglycerides and fatty acids are transported to the surfaces of the brush border microvilli penetrating into the recesses of the moving agitating microvilli. Here, the monoglycerides and the fatty acids diffuse through the epithelial membrane, because they are soluble in the membrane. The bile acid micelles is now left to diffuse back into the chyme, absorbing more monoglycerides and fatty acids. Undigested triglycerides and diglycerides are both highly soluble in the lipid membrane of epithelial cells. However, only small quantity of these are normally absorbed, because the bile micelles will not ferry them to the epithelial membrane. On entering the epithelial cells, fatty acids and monoglycerides are taken up by the small endoplasmic reticulum and are recombined to form new triglycerides. Once form the triglycerides aggregate within the endoplasmic reticulum into globules along with absorbed cholesterol and phospholipids globules are called chylomicrons. These globules diffuses to the side of the epithelial cells and is extrude by process of cellular exocytosis into the space between the cells. From the site of the epithelial cells, chylomicrons find their ways into the central lactus of the villi, and from here they are propelled along with the lymph by the lymphatic pump upward through the thoracic duct to be emptied into the great vein of the neck. Between 80 and 90% of all fat absorbed from the gut get into the interstitium in this manner. Absorption of proteins Amino acids (and some small peptides) are absorbed mainly in the duodenum and upper jejunum into the portal blood. D- amino acids are absorbed passively, whereas L-amino acids are absorbed actively by a Na-linked carrier mechanism. There are at least four specific mechanisms by which amino acids are absorbed: one for neutral amino acids, one for basic amino acids, one for acidic amino acids and one for the imino acids (proline, sarcosine). Dipeptides and tripeptides may be absorbed from the lumen and are later hydrolyzed within the epithelial cells. In neonates, antibodies and other proteins contained in colostrum may be absorbed in their intact form by pinocytosis. Absorption of Water Only a small amount of water move across the gastric mucosa but water moves in both directions across the mucosa of the small and large intestines in response to osmotic gradient. Absorption of ions/electrolytes Na+ is actively absorbed throughout the small and large intestines. Active transport of Na+ is important in the secondary transport of glucose and some amino acid. 66 NSC 203 HUMAN PHYSIOLOGY (II) Chloride ions are rapidly absorbed mainly by passive diffusion in the upper part of the small intestines. They move along with the absorbed Na+ to balance the electrical gradient caused by Na+ ion absorption. K+ ions are absorbed across the gastrointestinal mucosa by diffusion. Absorption of Vitamins Water soluble vitamins are rapidly absorbed while the absorption of fat soluble vitamins is dependent of fat absorption. Most vitamins are absorbed in the upper small intestine but vitamin B12 is absorbed in the ileum. 4.0 Conclusion Six different hormones secreted in different parts of the gastrointestinal tract are discussed with information of different functions performed to prepare the different organs to support digestion and absorption in different parts of the tract. 5.0 Summary In this unit, you have leant about the following hormones, pancreozymin (CCK – P2), Secretin, Gastric inhibitory peptide, (VIP) and Motilin. You have also learnt about digestion of the different areas of the gastro-intestinal tract. Absorption through diffusion in different areas of the gastro-intestinal tract. 6.0 6.1 6.2 Gastrin, Ccholecystokinin – Vasoactive intestinal peptide various smaller molecules in active, passive transport and Tutor Marked Assignments Activity: As directed by the facilitator. Answer the following questions: 1. Describe the forms and actions of the following hormones: a) Gastrin b) CCK – P2 (cholecystokinin – pancreozymin) c) Secretin d) Gastric inhibitory peptide e) Vasoactive intestinal peptide (VIP) f) Motilin 2. 7.0 References and other resources Fox, S.I. (2012). Human Physiology. 12th edition, Mc Graw Hill, New York. Ganong W.F. (2010). Review of Medical Physiology. 23rd edition, Mc Graw Hill, New York. Guyton, A.C& Hall J.E. (2001). Textbook of Medical Physiology. Harcourt International Edition, 10th edition, W.B. Saunders, Philadelphia. Oyebola, D.O. (2002). Essential Physiology, Vol 1, Nihort Press 67