Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
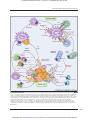
Published OnlineFirst December 7, 2012; DOI: 10.1158/0008-5472.CAN-12-2027 Cancer Research Review Role of Chemokines and Chemokine Receptors in Shaping the Effector Phase of the Antitumor Immune Response re2, and Katarzyna Franciszkiewicz1, Alexandre Boissonnas2, Marie Boutet1, Christophe Combadie Fathia Mami-Chouaib1 Abstract Immune system–mediated eradication of neoplastic cells requires induction of a strong long-lasting antitumor T-cell response. However, generation of tumor-specific effector T cells does not necessarily result in tumor clearance. CTL must first be able to migrate to the tumor site, infiltrate the tumor tissue, and interact with the target to finally trigger effector functions indispensable for tumor destruction. Chemokines are involved in circulation, homing, retention, and activation of immunocompetent cells. Although some of them are known to contribute to tumor growth and metastasis, others are responsible for changes in the tumor microenvironment that lead to extensive infiltration of lymphocytes, resulting in tumor eradication. Given their chemoattractive and activating properties, a role for chemokines in the development of the effector phase of the antitumor immune response has been suggested. Here, we emphasize the role of the chemokine–chemokine receptor network at multiple levels of the T-cell–mediated antitumor immune response. The identification of chemokine-dependent molecular mechanisms implicated in tumor-specific CTL trafficking, retention, and regulation of their in situ effector functions may offer new perspectives for development of innovative immunotherapeutic approaches to cancer treatment. Cancer Res; 72(24); 6325–32. 2012 AACR. Introduction The identification of tumor-associated antigens (TAA) and the isolation of tumor-specific cytotoxic T cells have led to great efforts in developing immunotherapeutic approaches to overcoming tumor invasion. Immunotherapy represents a promising approach to cancer treatment, with less severe side effects than conventional strategies. Major strategies have focused on the induction of T-cell–mediated antitumor responses. However, the generation of antigen-specific tumor-reactive T cells has rarely been translated into therapeutic success. One of the reasons for the failure of the immune system to eradicate cancer cells is a defect in T-cell migration to the tumor site (1). To destroy established tumors, CTL must traffic to and infiltrate the tumor tissue before specific activation and triggering of target cell death. The dissection of cellular and molecular processes that enhance T-cell recruitment and ultimately lead to tumor elimination is therefore a et de la Recherche Authors' Affiliations: 1Institut National de la Sante Medicale (INSERM) U753, Team 1: Tumor Antigens and T-cell Reactivity, Integrated Research Cancer Institute in Villejuif (IRCIV), Institut de rologie Gustave Roussy (IGR), Villejuif; and 2INSERM UMR-S 945 Cance Pierre et Marie Curie (UPMC University Paris 06), Laboratory and Universite ^ pital, Paris, France of Immunity and Infection, Boulevard de l'Ho Corresponding Author: Fathia Mami-Chouaib, Institut National de la et de la Recherche Medicale (INSERM) U753, IGR, 39 rue Sante Camille-Desmoulins, F-94805 Villejuif, France. Phone: 33-1-42-11-4965; Fax: 33-1-42-11-52-88; E-mail: [email protected] doi: 10.1158/0008-5472.CAN-12-2027 2012 American Association for Cancer Research. critical step in optimization of current cancer immunotherapy protocols. Chemokines coordinate circulation, homing, and retention of immune cells. Originally characterized for their ability to induce leukocyte chemoattraction, they are now recognized to orchestrate a wide array of leukocyte functions during inflammation and immunity (2). Indeed, in addition to their chemotactic properties, chemokines can directly regulate T-cell development, priming, and effector functions (3). In the context of cancer immunosurveillance, chemokines orchestrate the spatiotemporal distribution of immunocompetent cells crucial for induction of antitumor immune response and optimal effector function (4, 5). Chemokines constitute a large family of small, mostly secreted proteins comprising more than 50 members, which interact with 20 chemokine receptors. Chemokine receptors are G-protein–coupled 7-transmembrane–domain receptors responsible not only for triggering intracellular signals resulting in cell polarization, migration, and adhesion, but also for contributing to gene expression, cell proliferation, and survival (2). Most chemokines bind to more than 1 receptor. On the other hand, chemokine receptors display overlapping ligand specificities with variable affinity and functional activities (3). There is now overwhelming evidence that the chemokine– chemokine receptor system is directly or indirectly involved in tumor development (6). On the basis of their role in cell migration, chemokines contribute to tumor dissemination and metastasis. Moreover, chemokine-triggered signaling pathways can facilitate tumor cell proliferation and contribute to neovascularization. Tumor-derived chemokines are also www.aacrjournals.org Downloaded from cancerres.aacrjournals.org on June 18, 2017. © 2012 American Association for Cancer Research. 6325 Published OnlineFirst December 7, 2012; DOI: 10.1158/0008-5472.CAN-12-2027 Franciszkiewicz et al. responsible for shaping the tumor microenvironment into an immunosuppressive site, determining the qualitative and quantitative composition of tumor-infiltrating immune cells and affecting their maturation and activation status (7). In consequence, tumors seem to use chemokines to evade immunosurveillance and actively progress. Nevertheless, expression of some chemokines within the tumor bed has been associated with an effective antitumor immune response, an angiostatic effect, a low recurrence rate and increased patient survival. Indeed, some chemokines are responsible for changes in chemoattractive properties of the tumor microenvironment that allow extensive infiltration of leukocytes (7). Thus, chemokines and chemokine receptors represent valuable targets for optimizing antitumor immune responses. In this context, the major concern of tumor immunologists is to better understand chemokine-mediated pathways involved in T-cell recruitment at the tumor site and in regulating their intratumoral effector functions. In this review, we present findings implicating chemokines in regulation of the CTL-mediated effector phase of the antitumor immune response and we provide insights into their therapeutic applications. Role of Chemokines in Priming the T-Cell– Mediated Immune Response Development of an effective antitumor immune response relies on the coordinated interactions of immunocompetent cells, the spatiotemporal distribution of which is in part orchestrated by chemokines (Fig. 1). Acting through their cognate receptors, chemokines regulate trafficking between the tumor site and lymph nodes. To become competent killer cells, CTL require efficient priming by professional antigenpresenting cells (APC) and cognate licensing of dendritic cells by CD4þ T cells (8). For this purpose, na€ve CD8þ T cells continuously traffic through secondary lymphoid organs in which they systematically scan the surface of dendritic cells searching for TAA. Na€ve T cells express CCR7, which recognizes constitutively expressed CCL19 and CCL21. CCL21, produced by lymph nodes, Peyer's patch–associated high endothelial venules, and afferent lymphatic vessels, triggers a multistep process for recruitment of na€ve lymphocytes (9). Similarly, CCL21-CCR7 signaling is involved in trafficking of antigen-presenting dendritic cells. Indeed, their maturation into potent APC implies downregulation of tissue-specific chemokine receptors, such as CCR1, CCR5, and CCR6, and upregulation of CCR7, which guides dendritic cells from sites of antigen exposure to the local lymph nodes via draining afferent lymphatic vessels (10). The pivotal role of CCR7 in these processes was shown in CCR7-deficient mice, which displayed reduced numbers of na€ve T cells in secondary lymphoid organs (11). Once in the lymph nodes, T cells display high basal motility, enabling them to scan up to several thousand APC per hour. These steady-state movements are dependent on Gai-coupled chemokine receptor signaling triggered by CCL19 and CCL21 present in the lymph node T-cell zone (12). The major biologic relevance of na€ve T-cell motility within lymph nodes is to ensure recognition of a few antigen-bearing dendritic cells by rare specific T cells. However, random migration of T cells does not seem to be efficient enough to 6326 Cancer Res; 72(24) December 15, 2012 provide CD4 T-cell help for CD8 T-cell priming, where 2 lymphocyte types have to encounter the same APC (13). It has been shown that CCL19 secreted by mature dendritic cells increases na€ve CD4 T-cell scanning behavior and their response to rare cognate antigens (14). Moreover, engagement of na€ve CD4 T cells with APC triggers secretion of CCL3 and CCL4, which favor CCR5-dependent guidance of na€ve CD8 T cells toward dendritic cells (DC)-CD4 T-cell conjugates (15). Interestingly, CCR5-dependent recruitment of CD8 T cells to dendritic cells engaging TAA-specific T cells increases the efficiency of alternative TAA-specific na€ve CD8 T-cell priming (16). Following the antigen encounter and T-cell expansion, a modification in the general chemokine receptor expression profile is required to enable appropriate redistribution of activated T cells. This modification consists of downregulation of receptors that mediate entry and the encounter with APC in the lymph node, and of upregulation of other receptors to first egress from the lymph node and then sensitize T cells to infiltrate the tumor site. Pertussis toxin has been shown to affect lymphocyte egress from the lymph node, suggesting an implication of chemokine receptors. Indeed, one of the initial events in effector T-cell differentiation is downregulation of CCR7 and upregulation of receptors specific to chemokines expressed in target tissues, such as CCR1, CCR2, CCR3, CCR5, and CXCR3 (17). Acquisition of an appropriate migration program is thus crucial for targeting the right cell to the right place. Regulation of T-Cell Recruitment at the Tumor Site by Chemokines To exert their functions, recently primed T cells leave the lymph node and migrate to the tumor site, in which they physically engage cognate T-cell receptor (TCR) ligand-expressing targets. Chemokines play a major role in the recruitment of effector T cells within the tumor microenvironment. Antitumor CTL respond to numerous inflammatory chemokines, mainly CCL3 (18), CCL5 (19), CCL20 (20), and CXCL10 (21), which can be produced at the tumor site (22). Therefore, the intratumoral production of chemokines, which determine optimal T-cell recruitment, is one of the key factors in an efficient antitumor immune response. In this context, CCL5 was one of the first chemokines implicated in regulating antitumor immunity (19). A role for CCR5 in T-cell recruitment to the tumor site has been documented and local production of CCL5 or CCL3 induces selective recruitment of CD8 T cells and CTL-dependent tumor suppression in mouse models (23). CCR5 ligands are detected in many human tumors, including non–small-cell lung carcinoma (NSCLC), and can induce T-cell infiltration. However, the role of CCL5 in the antitumor immune response remains controversial. In NSCLC, the production of CCL5 has been associated with an active lymphocyte-mediated response and represents a positive predictive factor for patient survival (24). In contrast, high levels of CCL5 have been reported to correlate with poor prognosis in breast and cervical carcinomas (25). Although the cellular infiltrates were not investigated in these studies, it is conceivable that disease progression was related to tumor escape mechanisms Cancer Research Downloaded from cancerres.aacrjournals.org on June 18, 2017. © 2012 American Association for Cancer Research. Published OnlineFirst December 7, 2012; DOI: 10.1158/0008-5472.CAN-12-2027 Chemokines and the Antitumor Immune Response Lymph node MHC-II-TCR Migration to LN Maturation CD4ⴙ T cell CD4 CCR7 CD40–CD40L CCL19, 21 IL-2, IFNγ… DC Ag presentation MHC-I-TCR IDC CD8 CD8ⴙ T cell CD80/86-CD28 CCR1, 5, 6, 9 CXCR4 Ag uptake Innate immunity Adaptive immunity CCR2, 5 CXCR3 CCR2, 5, 9 CXC3R1 M NK CCR1, 2, 4, 5, 7, 8 CXCR1, 2, 3, 4 (HSP, Calreticulin…) XCR1 NKT Lysis CCL2, 3, 5, 16 Dying tumor cells CXCL8, 9, 10, 12, 21 CCR5, 6 CXCR3, 4, 6 Danger signals CCL3 CCL4 CCL3, 4, 5 Migration to the tumor CXCL9, 10, 11 CXCL10, 12, 16 PRR CD4ⴙ Th1 cell CCR5 CXCR3 XCL1 IFNα DC precursor CD8ⴙ CTL Specific lysis PAMP CCR3, 4, 8 Treg CD4ⴙ Th2 cell CCR2, 4, 5, 6, 8 CCR2, 4, 5, 6, 8 MDSC Tumor site Figure 1. Chemokine network in the antitumor immune response. Malignant cells express pathogen-associated molecular patterns (PAMP) that can be recognized by pattern recognition receptors (PRR) on dendritic cells (DC) and macrophages (M), triggering release of chemokines. This results in recruitment and activation of M, NK, and NKT cells, which are able to lyse tumor cells. DC phagocytose apoptotic tumor cells and HSP-complexed tumor-derived peptides. Upon maturation, DC change their homing proprieties by downregulating tissue-specific chemokine receptors and upregulating CCR7 that guides them to CCL19/CCL21-rich lymph nodes (LN), where they present processed tumor þ þ peptides to CD4 and CD8 T cells. Activated T cells upregulate expression of chemokine receptors including CCR5 and CXCR3, and in response to intratumoral chemokines, circulating CTL infiltrate the tumor to destroy malignant cells. Tumor-derived chemokines are also responsible for recruitment of Treg cells and MDSC, which participate in establishment of a protumoral microenvironment. Ag, antigen; IDC, immature DC. www.aacrjournals.org Cancer Res; 72(24) December 15, 2012 Downloaded from cancerres.aacrjournals.org on June 18, 2017. © 2012 American Association for Cancer Research. 6327 Published OnlineFirst December 7, 2012; DOI: 10.1158/0008-5472.CAN-12-2027 Franciszkiewicz et al. 6328 through concurrent recruitment of immunosuppressive cell populations, such as tumor-associated macrophages and regulatory T (Treg) cells, or triggering of tumor-infiltrating lymphocyte (TIL) apoptosis (26). Conversely, other groups provided evidence for CCL3 and CCL5-induced T-cell proliferation and activation (27, 28). Among other chemokines implicated in tumor infiltration by immune cells, CX3CL1 and CXCL16 have been associated with high numbers of CD8þ and CD4þ TIL and with good prognosis in colorectal cancer (29). CCL20 has been detected in breast cancer, in which it may be responsible for recruitment of CCR6þ memory T-cell subsets (30). The main chemokines attracting effector cells seem to be the CXCR3 ligands CXCL9 and CXCL10. In addition to their angiostatic activity, these chemokines participate in the antitumor immune response through recruitment of T and natural killer (NK) cells (23). In renal cell carcinoma, intratumoral expression of CXCL9 and CXCL10 coincided with a high degree of CD8 T-cell infiltration and elicited an inverse correlation with tumor growth and recurrence after curative surgery (31). High levels of CXCL9 have also been associated with strong infiltration of malignant melanoma by CD8 T cells and improvement in patient survival (32). Thus, it seems evident that chemokines contribute to the establishment of the immune response by orchestrating the distribution of its key cellular components and delivering the generated effectors to the tumor site. Whether intratumoral chemokines promote immune surveillance or tumor escape depends on the composition of cell infiltrates, which shape the environmental context through an immunoediting process. Interestingly, nonspecific tumor T cells can deeply infiltrate tumor tissues only upon tumor destruction by antigen-specific CTL (35). This suggests that changes in the tumor microenvironment, such as secretion of chemokines induced by destruction in tumor architecture, determine deep lymphocyte infiltration. Therefore, delivery of appropriate chemokines into the tumor, reminiscent of what takes place during an effective T-cell response, would induce extensive infiltration of effector cells, and thereby tumor destruction. The contribution of chemokine receptors in T-cell retention at the tumor site is not well documented. We recently reported that recruitment of CCR5 at the immune synapse formed between TIL and tumor cells, resulted in inhibition of T-cell responsiveness to a CCL5 chemotactic gradient. This CCR5 clustering is dependent on the interaction of the aE(CD103)b7 integrin on T cells with its ligand, E-cadherin, on NSCLC cells (36). An alternative mechanism of CTL retention in epithelial tumors could implicate CCR6. CCR6þ T cells were found to be more frequent in CD8 subpopulations isolated from TIL than from PBL (36). This observation, together with efficient homing of CCR6 CTL to the tumor site, suggests that CCR6 is not involved in T-cell recruitment to NSCLC but may play a local role. Apart from its function in cell trafficking, CCR6 was reported to be crucial for cell activation and conformational changes in integrins (37). Furthermore, it has been shown that the interaction of CCR6 with its unique ligand, CCL20, is a critical event in the arrest of effector/memory T cells on endothelial cells (38). It is therefore conceivable that intratumoral induction of CCR6, together with CD103, play a role in Tcell retention at the tumor site. Intratumoral T-Cell Location, Motility, and Retention Costimulatory Role of Chemokines–Chemokine Receptors in T-Cell Activation Positioning of CTL within tumor tissues is critical for an efficient antitumor immune response. After extravasation, CTL must migrate through the interstitial space of the tumor to recognize and kill target cells. Thus far, little is known about the intratumoral migration of infiltrating CTL. In experimental studies, CD8 T cells were mostly observed at the tumor periphery, with limited infiltration into the tumor mass. A similar pattern was identified in human tissue sections from metastatic melanoma. In NSCLC, T cells accumulate in stromal regions in which chemokines likely participate in controlling their motility and their entry into tumor islets (33). Before establishing stable contact with target cells, TIL migrate randomly within the tumor microenvironment, arguing against the early concept of T-cell guidance by a long-range chemokine gradient. Two-photon microscopy studies provided insight into the dynamics of infiltration and elimination of solid tumors by immune cells (34, 35). After diapedesis, effector T cells initially display random migration and start engaging a transient antigen-independent interaction with target cells. CTL that encounter antigen-expressing tumor cells, arrest their migration to establish a stable contact with the target. This leads to release of cytokines and cytotoxic granules resulting in tumor cell death. Once cancer cells are cleared, CTL resume motility to further search for new target cells. Although there is a considerable evidence implicating chemokines in antigen-experienced T-cell recruitment to the tumor site, their role in T-cell effector function is begging for intensified investigation. It has been widely reported that engagement of chemokine receptors triggers a "go" signal that competes with TCR-mediated "stop" signals, and thus negatively influences the stability and duration of the immune synapse (39). Indeed, the gradient of some chemokines, including CXCL10 and CCL19 or CCL21, renders T cells ignorant of agonist peptide-MHC (pMHC), thus suppressing their activation. However, CCL19 secreted by antigen-activated dendritic cells induces T-cell polarization and motility, resulting in improved scanning of APC, thus increasing cognate pMHC encounters (14). Chemokines such as CXCL12 were also reported to enhance adhesion of T cells to dendritic cells by regulating the avidity/ affinity of key integrins, including leukocyte function-associated antigen (LFA)-1 (37). Other chemokines, namely CCR7 ligands bound on dendritic cells surface, may promote T-cell activation by improving immune synapse formation (40). CCR5 and CXCR4 can be recruited to the immune synapse during Tcell stimulation by APC, leading to a reduction in T-cell sensitivity to other chemokine sources and enhanced T-cell responses (27). It has also been proposed that APC-derived chemokines can act as costimulatory molecules for engaged T Cancer Res; 72(24) December 15, 2012 Cancer Research Downloaded from cancerres.aacrjournals.org on June 18, 2017. © 2012 American Association for Cancer Research. www.aacrjournals.org Prostate cancer (M), breast cancer (M), melanoma (M) Mammary carcinoma (M) T cells Lung cancer (M) Mammary carcinoma (M) Breast carcinoma (M) Colon adenocarcinoma (M) Melanoma (M and H phase I trial) Lung cancer (M) Colon carcinoma (M) Melanoma (M) Breast cancer (M) Melanoma (M) Neuroblastoma (H) Ovarian carcinoma (M) Colorectal carcinoma (M) Melanoma (H) CCL2, CCL3 (rCCL2 and/or rCCL3) CCL5 (rCCL5, Ad-RANTES-E1A) CCL19 (rCCL19) CCL20 (rCCL20) CCL21 (rCCL21, nanoparticles, Ad-CCL21–transduced DC) CXCL8 (rCXCL8) CX3CL1 (rCX3CL1) CXCR1 (adoptive immunotherapy) TIL Macrophages, neutrophils T cells, NK cells CD4þ and CD8þ T cells, NK cells CD8þ T cells, NK cells T cells DC CD4, NK cells CD8þ T cells, DC Macrophage Macrophage, T cells Immune effector cells Infiltrating cells Lymphocyte-dependent response Unknown Increase of immune cell infiltration into tumors Increase of tumor infiltration by CD4þ and CD8þ T cells, and neutrophils Tumor-infiltrating T cells Increase of immune cell infiltration into tumors T-cell proliferation, cytokine secretion APC maturation and T-cell priming Increase of tumor infiltration by immune cells Unknown Lymphocyte-dependent response Lymphocyte-dependent response Immune response Increase of TIL recruitment into CXCL1 and CXCL8expressing tumors Tumor regression Tumor regression Tumor regression Tumor growth inhibition Complete tumor eradication Tumor rejection Tumor growth inhibition Tumor regression Tumor rejection Tumor regression Tumor regression Treatment outcome NOTE: Respective human chemokines, DC, or adenoviral vectors encoding human chemokines were injected directly at the tumor site. Abbreviations: Ad, adenovirus; DC, dendritic cells; (H), human; (M), mouse; r: recombinant. XCL1 (Ad-XCL1 þ IL-2 or IL-12 or XCL1 þ gp100pulsed DC) CCL22 (Ad-CCL22) CCL16 (rCCL16, CCL16 fusion protein: LEC-chTNT3) Mouse (M) or Human (H) tumor model Human chemokines (therapeutic approach) Table 1. Experimental and clinical trials based on intratumoral delivery of human chemokines (55) (46) (47) (53) (54) (5) (29) (23) (52) (5) (5) (27) (52) (52) (5) References Published OnlineFirst December 7, 2012; DOI: 10.1158/0008-5472.CAN-12-2027 Chemokines and the Antitumor Immune Response Cancer Res; 72(24) December 15, 2012 Downloaded from cancerres.aacrjournals.org on June 18, 2017. © 2012 American Association for Cancer Research. 6329 Published OnlineFirst December 7, 2012; DOI: 10.1158/0008-5472.CAN-12-2027 Franciszkiewicz et al. cells through chemokine receptors relocalized at the immune synapse (27). Role of Chemokines in Shaping the Tumor Microenvironment The tumor microenvironment is characterized by chronic inflammation with the presence of cytokines and chemokines, the profile of which dictates neoplastic outcome (7). Indeed, chemokines mediate accumulation of immunocompetent cells and participate in shaping a tumor-promoting or -suppressive microenvironment. CCR5 and CXCR3 predominate on Th1 cells, whereas Th2 cells preferentially express CCR3, CCR4, and CCR8. At the tumor site, Th1 cells colocalize with macrophages and neutrophils, enhancing the cell-mediated immune response. In contrast, Th2-type inflammation is considered to be protumoral and is associated with a poor prognosis (41). Chemokine receptor patterns that guide CD8 T cells to target tissues have not been studied as extensively as CD4 T cells but seem to be similar. Upon antigen-reencounter, CD8 T cells secrete inflammatory chemokines, such as CCL3 and CCL5, which increase infiltration of neutrophils, monocytes, and Th1 lymphocytes, and thus contribute to so-called autorecruitment of CTL, resulting in amplification of effector responses. However, the protective activity of TIL is often compromised by immunosuppressive tumor microenvironment components, including Treg cells. Treg can be actively attracted to the tumor mostly via CCR4. Infiltration by Treg of ovarian tumors producing high levels of CCL17 and CCL22 correlated with unfavorable prognosis (42). Treg cells, recruited through CCL22-CCR4, can be activated in lymphoid infiltrates surrounding breast tumors, leading to an adverse clinical outcome (43). Other immunosuppressive cells participating in shaping the tumor microenvironment are myeloidderived suppressor cells (MDSC; Fig. 1). Once recruited within tumors in a CCR2, CXCR4, or CXCR2-dependent manner, MDSC have a significant effect on tumor progression, mainly through suppression of antitumor effectors (44). Thus, by determining the composition of cellular infiltrates, intratumoral chemokines continuously shape the tumor microenvironment and regulate the extent of antitumor immune responses. Chemokines in Cancer Immunotherapy Strategies of chemokine–chemokine receptor–based tumor immunotherapy are aimed at eradicating tumors by inhibiting survival and metastasis of malignant cells. The overexpression of chemokines at the tumor site usually resulted in infiltration by host leukocytes; however, disease outcome was more divergent and related to the nature of the injected chemokines, the subset of recruited immune cells and the tumor model (Table 1). In mice, intratumoral delivery of chemokines can induce tumor suppression and immunity to subsequent tumor challenge through recruitment of dendritic cells, NK, and T cells (5). However, the efficacy of such approaches seems limited in humans (23). As chemokines are also responsible for attraction of immunosuppressive cells, such as Treg cells, blocking the activity of the host CCR5 has been proposed for improving the potency of dendritic cell–based vaccines against melanoma 6330 Cancer Res; 72(24) December 15, 2012 (45). Combination of chemokines with cytokines for cancer immunotherapy seems to deserve consideration. A combination of XCL1 and interleukin (IL)-2 has been reported to provide enhanced and long-term antitumor immunity (46). Other approaches to chemokine-based tumor immunotherapy include vaccination strategies with TAA. Despite a great deal of effort, the rate of objective cancer regression in vaccinated patients has remained weak. Among the limitations of such vaccines are the insufficient numbers of recruited effector cells and their inappropriate activation in an immunosuppressive tumor microenvironment (1). In this context, manipulating the chemokine network may represent an attractive adjuvant strategy (Table 1). A vaccine based on XCL1- and IL-2– secreting neuroblastoma cells induced an increase in T-cell infiltration and resulted in complete or partial tumor remission in vaccinated patients (47). Adoptive immunotherapy is currently one of the most promising approaches, with significant positive results in preclinical and clinical trials (48, 49). The success of adoptive therapy depends on the optimal selection and/or genetic engineering of antigen-specific cells, induction of their proliferation while preserving effector functions, engraftment ability, and efficient homing to the tumor (50, 51). Because recruitment of transfused lymphocytes at the tumor site is one of the critical steps, intratumoral expression of adequate chemokines represents one of the strategies for improving adoptive immunotherapy. Moreover, redirecting the migratory properties of adoptively transferred T cells toward chemokine-secreting tumors can be achieved by genetic expression of appropriate chemokine receptors. Thus, the intratumoral production of chemokines, which determine the optimal CTL recruitment, is one of the key factors for efficient antitumor responses. Any alteration can result in tumor evasion and represent one of the conceivable reasons for the failure of adoptive transfer or vaccination-based immunotherapies (4). Concluding Remarks The success of cancer immunotherapeutic strategies relies on the generation of efficacious effector mechanisms associated with the presence of high-avidity tumor-specific CTL. Chemokines contribute to induction of an effective immune response, orchestrating distribution of its key cellular components, and delivering the generated effectors to the target. It should, however, be noted that the chemokine– chemokine receptor network is also implicated in immune system homeostasis, thus limiting its manipulation in a clinical setting. Despite these obstacles, intensified investigation of the role of chemokines in tumor immunosurveillance and immunosuppression may provide a gateway to development of innovative and effective strategies for cancer immunotherapy. Disclosure of Potential Conflicts of Interest No potential conflicts of interest were disclosed. Authors' Contributions Conception and design: K. Franciszkiewicz, M. Boutet, C. Combadiere, F. Mami-Chouaib Development of methodology: F. Mami-Chouaib Cancer Research Downloaded from cancerres.aacrjournals.org on June 18, 2017. © 2012 American Association for Cancer Research. Published OnlineFirst December 7, 2012; DOI: 10.1158/0008-5472.CAN-12-2027 Chemokines and the Antitumor Immune Response Acquisition of data (provided animals, acquired and managed patients, provided facilities, etc.): F. Mami-Chouaib Analysis and interpretation of data (e.g., statistical analysis, biostatistics, computational analysis): F. Mami-Chouaib Writing, review, and/or revision of the manuscript: K. Franciszkiewicz, A. Boissonnas, M. Boutet, C. Combadiere, F. Mami-Chouaib Administrative, technical, or material support (i.e., reporting or organizing data, constructing databases): K. Franciszkiewicz, F. Mami-Chouaib Study supervision: F. Mami-Chouaib Grant Support This work was supported by grants from Institut National de la Sante et de la Recherche Medicale (INSERM), the Association pour la Recherche sur le Cancer (ARC), the Ligue contre le Cancer, and Institut National du Cancer. K. Franciszkiewicz was supported by a fellowship from the ARC and INCa. Received May 21, 2012; revised September 13, 2012; accepted October 1, 2012; published OnlineFirst December 7, 2012. References 1. 2. 3. 4. 5. 6. 7. 8. 9. 10. 11. 12. 13. 14. 15. 16. 17. 18. 19. Gajewski TF, Fuertes M, Spaapen R, Zheng Y, Kline J. Molecular profiling to identify relevant immune resistance mechanisms in the tumor microenvironment. Curr Opin Immunol 2011;23:286–92. Zlotnik A, Yoshie O. The chemokine superfamily revisited. Immunity 2012;36:705–16. Viola A, Sarukhan A, Bronte V, Molon B. The pros and cons of chemokines in tumor immunology. Trends Immunol 2012;33:496–504. Dell'Agnola C, Biragyn A. Clinical utilization of chemokines to combat cancer: the double-edged sword. Expert Rev Vaccines 2007;6: 267–83. Homey B, Muller A, Zlotnik A. Chemokines: agents for the immunotherapy of cancer? Nat Rev Immunol 2002;2:175–84. Balkwill FR. The chemokine system and cancer. J Pathol 2012;226: 148–57. Coussens LM, Werb Z. Inflammation and cancer. Nature 2002;420: 860–7. Smith CM, Wilson NS, Waithman J, Villadangos JA, Carbone FR, Heath WR, et al. Cognate CD4(þ) T cell licensing of dendritic cells in CD8(þ) T cell immunity. Nat Immunol 2004;5:1143–8. Gunn MD, Tangemann K, Tam C, Cyster JG, Rosen SD, Williams LT. A chemokine expressed in lymphoid high endothelial venules promotes the adhesion and chemotaxis of naive T lymphocytes. Proc Natl Acad Sci U S A 1998;95:258–63. Sallusto F, Lanzavecchia A. Understanding dendritic cell and T-lymphocyte traffic through the analysis of chemokine receptor expression. Immunol Rev 2000;177:134–40. Forster R, Schubel A, Breitfeld D, Kremmer E, Renner-Muller I, Wolf E, et al. CCR7 coordinates the primary immune response by establishing functional microenvironments in secondary lymphoid organs. Cell 1999;99:23–33. Asperti-Boursin F, Real E, Bismuth G, Trautmann A, Donnadieu E. CCR7 ligands control basal T cell motility within lymph node slices in a phosphoinositide 3-kinase-independent manner. J Exp Med 2007; 204:1167–79. Castellino F, Huang AY, Altan-Bonnet G, Stoll S, Scheinecker C, Germain RN. Chemokines enhance immunity by guiding naive CD8þ T cells to sites of CD4þ T cell-dendritic cell interaction. Nature 2006;440:890–5. Kaiser A, Donnadieu E, Abastado JP, Trautmann A, Nardin A. CC chemokine ligand 19 secreted by mature dendritic cells increases naive T cell scanning behavior and their response to rare cognate antigen. J Immunol 2005;175:2349–56. Castellino F, Germain RN. Chemokine-guided CD4þ T cell help enhances generation of IL-6RalphahighIL-7Ralpha high prememory CD8þ T cells. J Immunol 2007;178:778–87. re C, Hugues S, Scholer A, Boissonnas A, Nussbaum A, Combadie Amigorena S, et al. Dynamic imaging of chemokine-dependent CD8þ T cell help for CD8þ T cell responses. Nat Immunol 2007;8:921–30. Moser B, Loetscher P. Lymphocyte traffic control by chemokines. Nat Immunol 2001;2:123–8. Maric M, Liu Y. Strong cytotoxic T lymphocyte responses to a macrophage inflammatory protein 1alpha-expressing tumor: linkage between inflammation and specific immunity. Cancer Res 1999;59: 5549–53. Mule JJ, Custer M, Averbook B, Yang JC, Weber JS, Goeddel DV, et al. RANTES secretion by gene-modified tumor cells results in loss of tumorigenicity in vivo: role of immune cell subpopulations. Hum Gene Ther 1996;7:1545–53. www.aacrjournals.org 20. Fushimi T, Kojima A, Moore MA, Crystal RG. Macrophage inflammatory protein 3alpha transgene attracts dendritic cells to established murine tumors and suppresses tumor growth. J Clin Invest 2000;105:1383–93. 21. Luster AD, Leder P. IP-10, a -C-X-C- chemokine, elicits a potent thymus-dependent antitumor response in vivo. J Exp Med 1993; 178:1057–65. 22. Vicari AP, Caux C. Chemokines in cancer. Cytokine Growth Factor Rev 2002;13:143–54. 23. Johrer K, Pleyer L, Olivier A, Maizner E, Zelle-Rieser C, Greil R. Tumourimmune cell interactions modulated by chemokines. Expert Opin Biol Ther 2008;8:269–90. 24. Moran CJ, Arenberg DA, Huang CC, Giordano TJ, Thomas DG, Misek DE, et al. RANTES expression is a predictor of survival in stage I lung adenocarcinoma. Clin Cancer Res 2002;8:3803–12. 25. Niwa Y, Akamatsu H, Niwa H, Sumi H, Ozaki Y, Abe A. Correlation of tissue and plasma RANTES levels with disease course in patients with breast or cervical cancer. Clin Cancer Res 2001;7: 285–9. 26. Mellado M, de Ana AM, Moreno MC, Martinez C, Rodriguez-Frade JM. A potential immune escape mechanism by melanoma cells through the activation of chemokine-induced T cell death. Curr Biol 2001; 11:691–6. 27. Molon B, Gri G, Bettella M, Gomez-Mouton C, Lanzavecchia A, Martinez AC, et al. T cell costimulation by chemokine receptors. Nat Immunol 2005;6:465–71. 28. Wong M, Fish EN. RANTES and MIP-1alpha activate stats in T cells. J Biol Chem 1998;273:309–14. 29. Ohta M, Tanaka F, Yamaguchi H, Sadanaga N, Inoue H, Mori M. The high expression of Fractalkine results in a better prognosis for colorectal cancer patients. Int J Oncol 2005;26:41–7. 30. Bell D, Chomarat P, Broyles D, Netto G, Harb GM, Lebecque S, et al. In breast carcinoma tissue, immature dendritic cells reside within the tumor, whereas mature dendritic cells are located in peritumoral areas. J Exp Med 1999;190:1417–26. 31. Kondo T, Ito F, Nakazawa H, Horita S, Osaka Y, Toma H. High expression of chemokine gene as a favorable prognostic factor in renal cell carcinoma. J Urol 2004;171:2171–5. 32. Mullins IM, Slingluff CL, Lee JK, Garbee CF, Shu J, Anderson SG, et al. CXC chemokine receptor 3 expression by activated CD8þ T cells is associated with survival in melanoma patients with stage III disease. Cancer Res 2004;64:7697–701. 33. Salmon H, Franciszkiewicz K, Damotte D, Dieu-Nosjean MC, Validire P, Trautmann A, et al. Matrix architecture defines the preferential localization and migration of T cells into the stroma of human lung tumors. J Clin Invest 2012;122:899–910. 34. Mrass P, Takano H, Ng LG, Daxini S, Lasaro MO, Iparraguirre A, et al. Random migration precedes stable target cell interactions of tumorinfiltrating T cells. J Exp Med 2006;203:2749–61. 35. Boissonnas A, Fetler L, Zeelenberg IS, Hugues S, Amigorena S. In vivo imaging of cytotoxic T cell infiltration and elimination of a solid tumor. J Exp Med 2007;204:345–56. 36. Franciszkiewicz K, Le Floc'h A, Jalil A, Vigant F, Robert T, Vergnon I, et al. Intratumoral induction of CD103 triggers tumor-specific CTL function and CCR5-dependent T-cell retention. Cancer Res 2009; 69:6249–55. 37. Laudanna C, Kim JY, Constantin G, Butcher E. Rapid leukocyte integrin activation by chemokines. Immunol Rev 2002;186:37–46. Cancer Res; 72(24) December 15, 2012 Downloaded from cancerres.aacrjournals.org on June 18, 2017. © 2012 American Association for Cancer Research. 6331 Published OnlineFirst December 7, 2012; DOI: 10.1158/0008-5472.CAN-12-2027 Franciszkiewicz et al. 38. Fitzhugh DJ, Naik S, Gonzalez E, Caughman SW, Hwang ST. CC chemokine receptor 6 (CCR6) is a marker for memory T cells that arrest on activated human dermal microvascular endothelium under shear stress. J Invest Dermatol 2000;115:332. 39. Dustin ML. Stop and go traffic to tune T cell responses. Immunity 2004;21:305–14. 40. Friedman RS, Jacobelli J, Krummel MF. Surface-bound chemokines capture and prime T cells for synapse formation. Nat Immunol 2006; 7:1101–8. 41. De Monte L, Reni M, Tassi E, Clavenna D, Papa I, Recalde H, et al. Intratumor T helper type 2 cell infiltrate correlates with cancer-associated fibroblast thymic stromal lymphopoietin production and reduced survival in pancreatic cancer. J Exp Med 2011;208:469–78. 42. Curiel TJ, Coukos G, Zou L, Alvarez X, Cheng P, Mottram P, et al. Specific recruitment of regulatory T cells in ovarian carcinoma fosters immune privilege and predicts reduced survival. Nat Med 2004; 10:942–9. 43. Gobert M, Treilleux I, Bendriss-Vermare N, Bachelot T, Goddard-Leon S, Arfi V, et al. Regulatory T cells recruited through CCL22/CCR4 are selectively activated in lymphoid infiltrates surrounding primary breast tumors and lead to an adverse clinical outcome. Cancer Res 2009; 69:2000–9. 44. Nagaraj S, Gabrilovich DI. Myeloid-derived suppressor cells. Adv Exp Med Biol 2007;601:213–23. 45. Ng-Cashin J, Kuhns JJ, Burkett SE, Powderly JD, Craven RR, van Deventer HW, et al. Host absence of CCR5 potentiates dendritic cell vaccination. J Immunol 2003;170:4201–8. 46. Dilloo D, Bacon K, Holden W, Zhong W, Burdach S, Zlotnik A, et al. Combined chemokine and cytokine gene transfer enhances antitumor immunity. Nat Med 1996;2:1090–5. 47. Rousseau RF, Haight AE, Hirschmann-Jax C, Yvon ES, Rill DR, Mei Z, et al. Local and systemic effects of an allogeneic tumor cell vaccine 6332 Cancer Res; 72(24) December 15, 2012 48. 49. 50. 51. 52. 53. 54. 55. combining transgenic human lymphotactin with interleukin-2 in patients with advanced or refractory neuroblastoma. Blood 2003; 101:1718–26. Dudley ME, Wunderlich JR, Robbins PF, Yang JC, Hwu P, Schwartzentruber DJ, et al. Cancer regression and autoimmunity in patients after clonal repopulation with antitumor lymphocytes. Science 2002; 298:850–4. Overwijk WW, Theoret MR, Finkelstein SE, Surman DR, de Jong LA, Vyth-Dreese FA, et al. Tumor regression and autoimmunity after reversal of a functionally tolerant state of self-reactive CD8þ T cells. J Exp Med 2003;198:569–80. June CH. Principles of adoptive T cell cancer therapy. J Clin Invest 2007;117:1204–12. Rosenberg SA, Restifo NP, Yang JC, Morgan RA, Dudley ME. Adoptive cell transfer: a clinical path to effective cancer immunotherapy. Nat Rev Cancer 2008;8:299–308. Lechner MG, Russell SM, Bass RS, Epstein AL. Chemokines, costimulatory molecules and fusion proteins for the immunotherapy of solid tumors. Immunotherapy 2011;3:1317–40. Emtage PC, Wan Y, Hitt M, Graham FL, Muller WJ, Zlotnik A, et al. Adenoviral vectors expressing lymphotactin and interleukin 2 or lymphotactin and interleukin 12 synergize to facilitate tumor regression in murine breast cancer models. Hum Gene Ther 1999;10: 697–709. Xia DJ, Zhang WP, Zheng S, Wang J, Pan JP, Wang Q, et al. Lymphotactin cotransfection enhances the therapeutic efficacy of dendritic cells genetically modified with melanoma antigen gp100. Gene Ther 2002;9:592–601. Sapoznik S, Ortenberg R, Galore-Haskel G, Kozlovski S, Levy D, Avivi C, et al. CXCR1 as a novel target for directing reactive T cells toward melanoma: implications for adoptive cell transfer immunotherapy. Cancer Immunol Immunother 2012;61:1833–47. Cancer Research Downloaded from cancerres.aacrjournals.org on June 18, 2017. © 2012 American Association for Cancer Research. Published OnlineFirst December 7, 2012; DOI: 10.1158/0008-5472.CAN-12-2027 Role of Chemokines and Chemokine Receptors in Shaping the Effector Phase of the Antitumor Immune Response Katarzyna Franciszkiewicz, Alexandre Boissonnas, Marie Boutet, et al. Cancer Res 2012;72:6325-6332. Published OnlineFirst December 7, 2012. Updated version Cited articles Citing articles E-mail alerts Reprints and Subscriptions Permissions Access the most recent version of this article at: doi:10.1158/0008-5472.CAN-12-2027 This article cites 55 articles, 20 of which you can access for free at: http://cancerres.aacrjournals.org/content/72/24/6325.full#ref-list-1 This article has been cited by 14 HighWire-hosted articles. Access the articles at: http://cancerres.aacrjournals.org/content/72/24/6325.full#related-urls Sign up to receive free email-alerts related to this article or journal. To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at [email protected]. To request permission to re-use all or part of this article, contact the AACR Publications Department at [email protected]. Downloaded from cancerres.aacrjournals.org on June 18, 2017. © 2012 American Association for Cancer Research.