Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
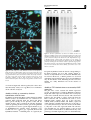
Gene Therapy (2005) 12, 1559–1572 & 2005 Nature Publishing Group All rights reserved 0969-7128/05 $30.00 www.nature.com/gt RESEARCH ARTICLE Bacterial transfer of large functional genomic DNA into human cells A Laner1,7, S Goussard2,7, AS Ramalho3, T Schwarz1, MD Amaral3,4, P Courvalin2, D Schindelhauer1,5,6 and C Grillot-Courvalin2 1 Department of Medical Genetics, Childrens Hospital, Ludwig Maximilians University, Munich, Germany; 2Unité des Agents Antibactériens, Institut Pasteur, Paris, France; 3Centre of Human Genetics, National Institute of Health Dr Ricardo Jorge, Lisboa, Portugal; 4Department of Chemistry and Biochemistry, Faculty of Sciences, University of Lisboa, Lisboa, Portugal; 5Institute of Human Genetics, Technical University, Munich, Germany; and 6Livestock Biotechnology, Life Sciences Center Weihenstephan, Freising, Germany Efficient transfer of chromosome-based vectors into mammalian cells is difficult, mostly due to their large size. Using a genetically engineered invasive Escherichia coli vector, alpha satellite DNA cloned in P1-based artificial chromosome was stably delivered into the HT1080 cell line and efficiently generated human artificial chromosomes de novo. Similarly, a large genomic cystic fibrosis transmembrane conductance regulator (CFTR) construct of 160 kb containing a portion of the CFTR gene was stably propagated in the bacterial vector and transferred into HT1080 cells where it was transcribed, and correctly spliced, indicating transfer of an intact and functional locus of at least 80 kb. These results demonstrate that bacteria allow the cloning, propagation and transfer of large intact and functional genomic DNA fragments and their subsequent direct delivery into cells for functional analysis. Such an approach opens new perspectives for gene therapy. Gene Therapy (2005) 12, 1559–1572. doi:10.1038/ sj.gt.3302576; published online 23 June 2005 Keywords: gene delivery; bacterial E. coli vector; human artificial chromosome; CFTR Introduction For functional analysis of genomic DNA of higher eukaryotes, stable cloning of large genomic fragments is well established in bacterial artificial chromosomes (BACs) and in P1-based artificial chromosomes (PACs) that are single-copy artificial chromosomes in Escherichia coli based on the F factor and P1 phage replicons, respectively.1,2 BACs and PACs can contain the majority of human genomic loci; most of them are comprised between 27 and 72 kb in size with just B4% larger than 140 kb.3 In addition, BACs and PACs allow the stable cloning of highly repetitive sequences, which have proven to be unstable in other cloning systems, as demonstrated for larger than 100 kb of highly homogeneous alpha satellite tandem repeat arrays of human centromeres.4 Successful gene therapy requires persistent tissuespecific expression of the transgene and could be optimally achieved by delivery of complete loci of genomic DNA of interest including native regulatory and promoter elements. To avoid random integration into the host chromosomes and allow stable inheritance, additional genetic elements are required. These are based Correspondence: C Grillot-Courvalin, Unité des Agents Antibactériens, Institut Pasteur, 25-28 rue du Docteur Roux, 75724 Paris cedex 15, France and D Schindelhauer, Livestock Biotechnology, Life Sciences Center, TUM, Hochfeldweg 1, 85354 Freising-Weihenstephan, Germany 7 These authors contributed equally to this work Received 10 February 2005; accepted 22 May 2005; published online 23 June 2005 either on viruses or, to circumvent viral protein expression, on natural host mechanisms. The most important of these elements is a functional centromere. Human artificial chromosomes (HACs) based on alpha satellite DNA as the only human component in a circular PAC or on linear, telomerized alpha satellite DNA, faithfully replicate and segregate during mitosis for many cell divisions even in the absence of selection.5 Specific alpha satellite DNA arrays from various human chromosomes, such as chromosomes 14, 17, 21, 22, X (and Y inefficiently), have been transferred as purified DNA in order to form centromeres on de novo HACS.5–13 The centromeres formed on the transferred DNA acquire chromatin proteins specific for functional centromeres such as CENP-A a histone H3 variant, only found within active centromere chromatin.13 Moreover, complete genes can be incorporated into HACs and expressed, as shown for human HPRT (42 kb),14,15 or GCH1 genes (61 kb).16 However, most gene delivery systems do not allow efficient transfer of large (4100 kb) DNA fragments into mammalian cells, thus limiting their functional analysis. The currently used viral derived vectors do not provide sufficient packaging capacity. High-capacity herpes simplex virus-based amplicon vectors are an exception but still cannot accommodate loci larger than 150 kb.17 Nonviral delivery systems based on lipofection, combined or not with polycations, have no size limits and have therefore been used with some success to transfer intact BAC DNA, both in vitro and in vivo.18 However, this technique requires production and purification of Bacterial transfer into human cells A Laner et al 1560 the DNA construct in significant amounts prior to transfection, a step that can in some cases impair its physical intactness. We have shown that invasive E. coli, which undergo lysis upon entry into mammalian cells because of impaired cell wall synthesis due to diaminopimelate (dap) auxotrophy, can efficiently deliver plasmid DNA to host mammalian cells.19 Plasmid pGB2Oinv-hly containing the inv gene from Yersinia pseudotuberculosis and the hly locus from Listeria monocytogenes has been introduced into the dap auxotroph E. coli BM2710. The inv gene confers to E. coli the ability to invade nonphagocytic cells provided they express b1-integrins. The hly gene product, listeriolysin O, is a pore-forming cytolysin that allows escape of the bacteria, or of its cytoplasmic content, from the vacuole of entry. Transfer of functional DNA into a variety of mammalian cell lines occurs after simple coincubation with the bacterial vector.20 Recombinant-deficient E. coli DH10B is widely used for the propagation of BAC clones. Recently, a dap DH10B derivative, similarly made invasive by transformation with pGB2Oinv DNA was able to mediate transfer into HeLa cells of a 200 kb BAC plasmid, as demonstrated by EGFP expression 48 h after transfection.21 We have used the recombination-deficient and dap auxotroph strain E. coli BM2710 expressing the invasin and listeriolysin O to deliver three large PAC-derived vectors into HT1080, a telomerase-proficient human lung sarcoma cell line often used to analyse HAC formation due to its relatively stable pseudotetraploid karyotype. These vectors included two PACs containing alphoid DNA, TTE122 and B2T8, of 150 and 200 kb, respectively, which could form HACs upon cell transfer, and an engineered 160 kb PAC construct, CGT21, which contains a large portion of the human locus containing the cystic fibrosis (CF) transmembrane conductance regulator (CFTR) gene fused to the EGFP gene as a marker.23 We demonstrate, for the first time, that bacterial gene delivery systems allow stable clone formation after transfer into cells of low copy number genetic elements of large size, either through the formation of free episomal HACs or through their integration into the host chromosome. Moreover, bacteria allow transfer of functional DNA of at least 80 kb, as demonstrated by expression and correct splicing in stable cell lines of the transferred construct. Results Invasive E. coli can stably propagate PAC-based constructs PAC vectors are usually stably maintained and propagated in recA1 E. coli DH10B after introduction by electrotransformation. We tested if recA1 strains BM2710 and BM4573, both derivatives of E. coli MM294,24 were also able to propagate stably PAC-based constructs. The three P1-derived plasmids used, TTE1, B2T8 and CGT21 (Figure 1), are compatible and can therefore coexist with pGB2Oinv-hly in E. coli BM2710. Figure 1 Maps of the plasmids used. (a) Plasmid TTE1 carries ca. 116 kb of a-satellite DNA from the centromeric region of human chromosome 5. The array was cloned as a NotI fragment from P1 artificial chromosome (PAC) E1 into the Bsp120 I site of the tetratelomeric PAC vector pTT. The pTT vector was derived from pTAT-BS by insertional duplication of vector portions, including telomeres, that resulted in tandem repeated vector sequences. An EGFPcDNA cassette (CMV/EGFP), two blasticidin S resistance genes (BS) and 2 pairs of telomeres (TTAGGG)n ¼ 135 (arrows) are present along with the unit copy replicon of phage P1 (P1). The location of primer pairs used to analyse the constructs by PCR are shown as black dots (5IF/5IR is present in multiple copies (ca. 340-fold) in the 0.34 kb higher order repeats throughout the array). (b) Plasmid B2T8, a ditelomeric PAC containing approximately 190 kb of asatellite DNA from human chromosome 17, was constructed by subcloning the NotI cut PAC B2 in the Bsp120 I site of pTAT-BS. The location of primer pairs is indicated by black dots (17-1A/17-2A is present, ca. 70-fold, in the 2.6 kb higher order repeats). (c) Plasmid CGT21 was engineered by cloning from a PAC containing the human CFTR locus from approximately 60 kb 50 to the middle of intron 9. To this CFTR region, a synthetic CFTR-EGFP fusion, composed of CFTR intron 9/exon 10 sequences, the EGFP cDNA CFTR exon 24 and 30 UTR and an additional 2 kb 30 region of the CFTR gene, was fused in frame. The construct was then cloned as a NotI fragment into the Bsp120 I site of the ditelomeric PAC vector pTAT-BS containing a blasticidin S resistance gene (BS). Gene Therapy Bacterial transfer into human cells A Laner et al Circular DNA from the three PAC plasmids, TTE1, B2T8 and CGT21 isolated from agarose plugs prepared from strain DH10B, was introduced by electrotransformation into E. coli BM2710/pGB2Oinv-hly and BM4573. Structural integrity and stable propagation of the plasmids in the new hosts were assessed, after 150 generations, by pulsed-field gel electrophoresis. Plasmid I-SceI fragments of the expected size were obtained from some of the clones studied, 124 kb for TTE1 (one out of two, clone F), 212 kb for B2T8 (three out of eight, clones U4, U7 and U8) and 151 kb for CGT21 (all clones studied, clones 20, 24, 34 and 38) (Figure 2). These data indicate efficient transfer by electrotransformation of DNA isolated from E. coli DH10B to E. coli BM2710/pGB2Oinv-hly and BM4573 and stable propagation of PAC-based constructs in these two bacterial vectors in the absence of major DNA rearrangements. Of note, the alphoid arrays tend to rearrange more than the CFTR region. Optimal conditions for invasion of HT1080 by invasive E. coli The first requirement for gene transfer from bacteria to mammalian cells to occur is efficient cell internalization of the bacteria, which varies depending on the cell line. The percentage of initially infected HT1080 cells was determined by flow cytometry analysis of fluorescent cells after invasion with bacteria expressing GFP under the control of a prokaryotic promoter. Invasive E. coli harbouring pAT505 (pUC18Ogfpmut1), a plasmid that directs synthesis of GFP in bacteria, was used at multiplicities of infection (MOI) (bacteria/mammalian cells ratio) of 5 or 25 to infect HT1080. After a 2 h coincubation, the cells were washed and incubated in complete medium containing 20 mg/ml of gentamicin to kill extracellular bacteria. After 45 min of incubation, the cells were trypsinized and analysed by flow cytometry. At an MOI of 5, 74.5% of the cells were fluorescent (Figure 3, panel a), and at an MOI of 25, 97.6% of the cells were strongly fluorescent (Figure 3, panel b). Viable intracellular bacteria were enumerated after gentle cell lysis; the number of viable bacteria per cell is calculated from the number of bacteria recovered per well. The number of viable intracellular bacteria recovered was 0.25 per cell at an MOI of 5 and 1.2 and at an MOI of 25. The discrepancy between the number of viable bacteria recovered from cells and the percentage of GFP-positive cells can be explained by the fact that dead or dying bacteria can still be GFP positive. Altogether, the results indicate that, at an MOI of 25, nearly all cells had internalized at least one GFP-producing bacterium. Cell viability, 48 h postinfection, ranged from 90 to 50% for an MOI of 5 and 25, respectively (data not shown). 1561 Bacterial vectors can stably deliver PAC-derived constructs into HT1080 cells Plasmid delivery into HT1080 cells was performed by simple coincubation for 2 h of HT1080 cells with E. coli BM2710/pGB2Oinv-hly harbouring plasmids B2T8 or CGT21, or E. coli BM4573 harbouring plasmid TTE1 at an MOI of 5 or 25. After 48 h in complete culture medium Figure 2 PFGE of constructs in bacteria. PAC DNA preparations in agarose plugs were digested with I-SceI, run on a pulsed field gel and stained with ethidium bromide. Clones with apparently intact inserts of approximately 124 kb (TTE1, clone F), 212 kb (B2T8, clones U4, U7, U8) and 151 kb (CGT21, clones 20, 24, 34, 38) indicated stability of the constructs. Plugs from strain DH10B are shown for comparison. The sizes of the inserts (in kb) are indicated on the side. (M, PFG mid range marker II, New England BioLabs). The plasmids are indicated at the bottom, and the host strains at the top. Gene Therapy Bacterial transfer into human cells A Laner et al 1562 with an average of 1.8 clones per 1 105 cells, B2T8 transfer by BM2710/pGB2Oinv-hly was less efficient with an average of 1.3 clone per 1 105 cells, whereas CGT21 delivery by the same bacterial vector was the least efficient with 0.25 clone per 1 105. The reasons for these differences could reside in the nature of the genetic element, HAC versus integration of a large DNA fragment for CGT21, or of the bacterial vector, BM4573 being more efficient than BM2710/pGB2Oinv-hly. Of note, bacterial delivery of pEGFP-C1 into HT1080 in similar conditions resulted, 48 h after transfer, in 0.3 and 1%, at an MOI of 5, and 1.2 and 2.9%, at an MOI of 25, EGFP-positive cells for BM2710/pGB2Oinv-hly and BM4573, respectively. A single internalized bacteria delivers ca. 500 copies of plasmid pEGFP-C1. Characterization by PCR of BS-resistant HT1080 clones The cell clones obtained were further characterized by PCR using the primer pairs described in Table 4 and Figure 1: BemF/BemR, LPF/LPR and KF/KR amplified, respectively, 159, 109 and 381 bp fragments present in the PAC vectors pTAT-BS and pTT. Primers 17-2A/T7 amplified a 150 bp fragment from B2T8 containing the junction between a-satellite DNA and the pTAT-BS vector, and EGF/EGR primer pair amplified a 281 bp fragment present in plasmids TTE1 and CGT21. Table 3 summarizes the results of the PCR experiments performed on a fraction of isolated BS-resistant clones. As expected, nearly all clones harboured apparently complete cloning vectors. Most importantly, in the majority of the clones harbouring plasmids B2T8 (11/12) and CGT21 (23/40), fragments from the insert (generated with primer pairs 17-2A/T7 and EGF/EGR, respectively) could also be amplified. Rare deletant derivatives were observed with B2T8 (1/12) and more frequently with TTE1 (5/14) and CGT21 (17/40). The cell clones were further investigated by FISH for detection of extrachromosomal elements for B2T8- and TTE1-derived clones and for the expression of genes present in TTE1 and CGT21. Figure 3 Efficiency of bacterial internalization by HT1080 cells. Cells infected with GFP-producing bacteria were analyzed by flow cytometry. E. coli BM2710 containing invasion pGB2Oinv-hly and reporter pUC18Ogfpmut1 plasmids were added at an MOI of (a) 5 or (b) 25. FL1-H fluorescence was monitored on cells trypsinized 45 min after cell invasion. with gentamicin to kill residual extracellular bacteria, the cells were cultured in the presence of blasticidin S (BS) for up to 3 weeks. Resistant clones were detected as early as day 14 and were not isolated after day 19, to avoid reseeding from existing clones. Clones were obtained, albeit at a low frequency in every experiment and with each PAC-derived vector (Table 2). The MOI of 25 yielded more clones for the same number of plated cells than that of 5. Of note, an MOI of 25 led to the internalization of at least one bacterium per cell and therefore of at least one PAC-derived vector molecule per cell. Efficiency of transfer was variable depending on both the plasmid and the bacterial vector: the highest efficiency was obtained for delivery of TTE1 by BM4573, Gene Therapy Centromere formation Dual colour FISH analysis using the Rsf vector probe and cen 17 probe in B2T8 lines U10-3 and U10-6 and cen 5 probe in TTE1 lines F100-1 and F100-4 was carried out after 30 days of growth with BS selection, which corresponds to approximately 30 generations. Free episomal HACs hybridizing with both probes (vector and cen) were found in most of metaphases in the four lines analysed (U10-3, n ¼ 13; U10-6, n ¼ 23; F100-1, n ¼ 12; F100-4, n ¼ 15). None of the lines showed integration into the host chromosomes. In the U10-3 and U10-6 lines grown for 30 additional days in the absence of BS, HACs that hybridized with the Rsf and the cen 17 probes were still detected (number of metaphases studied: U10-3, n ¼ 30 and U10-6, n ¼ 42) (Figure 4a); this demonstrates that transfer of B2T8 leads to the formation of stably segregating HACs. Similarly, dual colour FISH analysis with the Rsf and cen 5 probes, of the two stable F100-1 and F100-4 lines, after 30 generations without BS, showed one, two or rarely three HACs in the metaphases studied (n ¼ 19 and 23, respectively), indicating mitotic stability in the absence Bacterial transfer into human cells A Laner et al 1563 Figure 5 Southern hybridization of U10-3 and U10-6 cell lines. To analyse the stability of HAC DNA, chromosomal DNA of the U10-3 and U10-6 lines (indicated at the top) was prepared 30 days after growth in the presence (BS) and 30 days in the absence (D) of BS. Equal amounts (3 mg) of HindIII digested genomic DNA were loaded on a 1% agarose gel, electrophoresed (left panel), blotted and hybridized with the 32P-labelled probe Rsf including the BS-resistance marker of pTAT-BS (right panel). The similar intensities of the characteristic HindIII bands specific to B2T8 (3.4, 2.81 and 1.97 kb) show mitotic stability of the HACs even in the absence of selection. Figure 4 FISH analysis of stable HT1080 derived lines. Metaphases of B2T8 U10-6 (a) and TTE1 F100-4 (b) lines were prepared from cells grown 30 days in the presence of BS and 30 days in the absence of BS. Free low copy episomal HACs (white arrowheads) could be detected by colocalization of the vector probe rsf (red) and the centromere probe (green). In all metaphases analysed (n450 for both cell lines), no integration was observed with both probes. HACs at higher magnification are shown in the inserts. of selection (Figure 4b). Taken together, these data show that mitotically stable, low copy HACs have formed in all the cell lines analysed. Stability of HACs as assessed by Southern hybridization of B2T8 clones In addition to monitoring HAC stability directly by FISH, indirect analysis by Southern blot was carried out on genomic DNA prepared from the U10-3 and U10-6 clones grown 30 days in the presence of BS and for another 30 days with or without BS. The Rsf fragment (Table 4) spanning the BS resistance gene in the pTAT-BS vector portion was used as a probe. Equal amounts (3 mg) of HindIII-digested genomic DNA fragments were loaded onto a 1% agarose gel, separated, blotted and hybridized with the 32P-labelled probe. As expected, the Rsf probe hybridized with the bands corresponding to the B2T8 construct (3.4, 2.81 and 1.97 kb) (Figure 5), whereas no hybridization was detected in HT1080 (data not shown). Moreover, a similar pattern was observed with DNA prepared from clones grown for 30 additional days in the absence of BS, indicating that the resistance marker was stably inherited even in the absence of selection. Stability of TTE1-derived clones as assessed by EGFP expression The pTT PAC vector contains the EGFP expression cassette and lipofection of TTE1 in HT1080 resulted in bright and stable expression of EGFP in the vast majority of the clones obtained (data not shown). This observation indicates that the EGFP is stably expressed from the HAC constructs and can therefore be used to assess indirectly HAC stability. Four out of five cell lines studied, F100-1, F100-4, F300-3 and F300-6, expressed visible amounts of EGFP in more than 95% of the cells as assessed by FACS analysis (data not shown); in line F3005, only 50% of the cells expressed EGFP. All cell lines were divided into two parallel cultures at day 35 and grown with or without BS for an additional 30 days. The percentage of EGFP-expressing cells was determined by Gene Therapy Bacterial transfer into human cells A Laner et al 1564 Figure 6 Stability of EGFP expression from HACs in HT1080 lines during growth with (continuous lines) and without BS selection (dotted lines). Five cell lines (F100-1, F100-4, F300-3, F300-5 and F300-6) were divided into two cultures on day 35, and were passaged in the presence or in the absence of BS. The percentage of cells expressing EGFP was determined by flow cytometry at days 45, 55 and 65. flow cytometry at 10, 20 and 30 days after the split into two cultures (ie 45, 55 and 65 days after the initial transfection). As can be seen in Figure 6, the percentage of EGFP-expressing cells remained stable while selection was maintained for four of the five cell lines. The expression of EGFP by line F300-5 was stable until day 55, but started to decrease at day 65. When the cells were grown without selection, the percentage of EGFPpositive cells decreased slowly over time in three cell lines. There was a 50% drop after 30 days in a single line (F300-3), indicating an overall good mitotic stability in the absence of selection. Stable EGFP expression correlates with HAC formation in lines F100-1 and F100-4. Study of gene expression in CGT21-derived cell clones To evaluate the expression of the CGT21 construct and further assess its integrity after transfer, HT1080 cells were infected with E. coli BM2710/pGB2inv-hly harbouring this genomic expression construct and individual clones were obtained after BS selection. The CGT21 PAC construct results from the fusion of a portion of the human CFTR locus (exons 1–9 including 60 kb of upstream sequences) and of a synthetic exon made of CFTR exon 10, EGFP-cDNA, 30 UTR of CFTR exon 24, followed by the endogenous 30 CFTR genomic sequences as described.23 The resulting transcript encodes an artificial fusion protein without CFTR Cl channel function. This transcript, however, is easily distinguishable from endogenous CFTR transcripts by reverse transcription (RT)-PCR using a reverse primer complementary to the EGFP sequence (see Materials and methods). Gene Therapy From two independent bacterial transfer experiments, a total of 40 individual cell clones were isolated and expanded. Although PCR with EGF/EGR primers amplified an EGFP-specific fragment in 23 lines (Table 3), EGFP expression was never detected in those clones by FACS analysis (data not shown). For transcript analysis, RNA was extracted from 14 out of these 23 lines, subjected to long RT-PCR (see Materials and methods) and nine out of the 14 clones expressed transcripts from the CGT21 construct (Figure 7a). Clones showing CGT21 expression, albeit at various levels, were: 24-2, 24-4, and 38-1 (faint amplification was confirmed by sequencing of the product), 20B2, 34A2, 34A3, 34A4, 34B1 and 38A1 (see Figure 7a). Parallel amplification of b-actin (Figure 7b) showing equivalent levels of RT-PCR amplification in samples from all clones demonstrated similar efficiency in RNA extraction. Although these RT-PCR results cannot be fully interpreted in a quantitative manner, results in Figure 7a also suggest varying levels of expression of the CGT21 construct in different clones. The CGT21 RT-PCR product was sequenced in the nine expressing lines, and the sequences obtained confirmed that it corresponded to the correctly spliced CGT21 transcript spanning all exons between CFTR exon 1 and synthetic exon 10-EGFP (data not shown). Detection of the CGT21 transcripts as well as the fact they were correctly spliced in all nine cell lines indicates intact transfer of the CGT21 construct. Moreover, these data demonstrate maintenance of a genomic region of at least 80 kb spanning a minimal CFTR promoter. Two additional RT-PCR were performed to assess the relative levels of transcripts from the CGT21 construct and from the endogenous CFTR gene of the parental cell line. A semiquantitative approach was used consisting in two amplifications yielding products of similar size, but each being specific for one of the two transcripts. The results obtained for the 34A2 and 34A3 clones are shown in Figure 7c and indicate that the endogenous CFTR gene is also transcriptionally active in the parental cell line. Discussion Several laboratories have recently demonstrated that bacteria can transfer functional genes to a very broad range of mammalian cells.25,26 Attenuated strains of Shigella,27 invasive E. coli,19,20,28 Salmonella29 and of Listeria30 are able to transfer plasmid DNA to mammalian cells. The bacteria used for transfer to phagocytic and nonphagocytic cells are facultative intracellular pathogens that have been engineered to lyse after cell invasion. Until now, this property has been mainly exploited to develop DNA vaccines based on the ability of Shigella and Salmonella to target dendritic cells.27,29,31 More recently, invasive bacteria have also been used as DNA vectors for gene therapy. Administration to genetically immuno-deficient mice of avirulent Salmonella bearing the murine plasmid-borne IFN-g gene resulted in the expression of the transgene within macrophages and dendritic cells and correction of the genetic defect.32 Taking advantage of the proximity of bowel mucosa to luminal bacteria, we have successfully delivered a therapeutic gene to the colonic mucosa of mice. E. coli BM2710/pGB2Oinv-hly (Table 1) was able to deliver the Bacterial transfer into human cells A Laner et al 1565 Figure 7 RT-PCR analysis of CGT21 clones. (a) Results of long RT-PCR of samples from 14 cell lines transfected with the CGT21 genomic construct. Analysis was performed using primers A1R (CFTR exon 1) and GFP1-AL (synthetic exon 10-EGFP, Table 4). NC, negative control. (b) b-actin amplification of the same samples as a control for RNA extraction and the relative RNA levels among the samples. (c) Relative expression of the CGT21 construct and of endogenous CFTR in two cell lines, as analysed by two RT-PCR; one amplification specific for short CGT21 transcripts using the primers B3F (CFTR exon 8) and GFP1-AL (synthetic exon 10-EGFP), spans exon 8 of the CFTR gene to the EGFP sequence in the engineered fusion exon 10 of CGT21, yielding the 474 bp product (right arrow). The other amplification using the primers B3F and CI6D (CFTR exon 10) specific for endogenous CFTR transcripts, produces a 391 bp fragment (left arrow). The size of the inserts (in bp) is indicated on the sides (M1, 1 kb ladder; M2, 100 bp ladder). plasmid-carried TGF-b1 gene into the colonic epithelial cell line CMT-93 and oral administration to mice of the bacteria significantly reduced the severity of experimental DNBS-induced colitis.33 However, chromosome-based vectors are considered to be potentially useful to introduce exogenous genes into mammalian cells, since they replicate autonomously and have unlimited cloning capacity allowing insertion of entire genes with their proper endogenous and tissuespecific regulatory elements.34,35 Thus, simple and efficient techniques to transfer these very large molecules into mammalian cells need to be developed. We have demonstrated that BAC/PAC-based constructs can be transferred into mammalian cells by an invasive E. coli (Table 1) in which they can be introduced and propagated (Figure 2) resulting in de novo formation of HACs (Figure 4) or stable gene expression (Figures 6 and 7). As shown previously, using an inducible homologous recombination system adapted for use in E. coli DH10B,36 a dap derivative of this strain was generated and human genomic BAC clones were modified into suitable vectors for mammalian cells by the introduction of an EGFP neocassette.21 After transformation with plasmid pGB2Oinv DNA, the resulting invasive bacteria were able to deliver a 200 kb modified BAC to HeLa cells, resulting in 2.8% of cells expressing EGFP 48 h after bacterial invasion at high MOI (1500–4000); however, intactness of the transferred BAC and long-term BAC stability in the transfected cells were not studied. We demonstrate here that E. coli strain BM2710 can be transformed by large (from 140 to 220 kb) genomic constructs and that the bacterial vector allows stable cloning and propagation of the highly homogeneous tandem repeat arrays known to support de novo centromere formation (Figure 2). Bacterial transfer led to formation of stable HACs, which faithfully segregated in the absence of selection (Figure 4) and thus supported centromere formation by alpha satellite DNA. Until now, de novo formation was observed only after transfection of purified DNA from constructs by lipofection or microinjection. Since transfer occurs by direct cytoplasmic delivery upon lysis of intracellular dap bacteria28 without the need for DNA purification from E. coli, a possible negative effect of prokaryotic proteins bound to the PAC-DNA on the initial steps of HAC formation seems unlikely. Efficiency of stable clone recovery (Table 2) was in the same range as that observed with other transfer methods (ie 105–106, Magin-Lachmann et al18), although these figures are usually not rigorously documented; most studies consider early gene, such as egfp expression as an estimation of transfer efficiency. The optimal conditions for bacterial invasion defined in this study lead to more than one bacterium per cell (Figure 3) and therefore to more than one PAC-derived vector delivered intracellularly in each cell. Access to the cell cytosol of bacteria and plasmid DNA is an important requirement for successful gene transfer,37 but transfer to the nucleus is another limiting step as is the case for all other methods. Detection of free episomal HACs after bacterial transfer of TTE1 or B2T8 (Figure 4) do not prove per se transfer of intact centromere constructs. Owing to the highly homogeneous nature of alpha satellite arrays, Gene Therapy Bacterial transfer into human cells A Laner et al 1566 Table 1 Plasmids and bacterial strains used Designation Relevant characteristicsa Source or reference Plasmids pAT504 pAT505 pTAT-BS pTT B2T8 pGB2Oinv-hly, inv from Y. pseudotuberculosis and hly from L. monocytogenes, SmR, SpR pUC18Ogfpmut1, ApR ditelomeric PAC vector, PSV40-BSR; ApR, KmR tetratelomeric PAC vector, PSV40-BSR; PCMV-egfp, ApR, KmR pTAT-BSO190 kb a-satellite DNA array from human chromosome 17, PSV40-BSR; ApR, KmR 20 20 5 22 4, this work and unpublished data 22 23 TTE1 CGT21 E. coli DH10B BM2710 BM4573 pTTO116 kb a-satellite DNA array from human chromosome 5, PSV40-BSR, PCMV-egfp; ApR, KmR pTAT-BSOhuman CFTR locus with EGFP fusion, PSV40-BSR; ApR, KmR, TcR F mcrA D(mrr-hsdRMS-mcrBC) (f80dlacZDM15) DlacX74 deoR recA1 araD139 D(ara-leu)7697 galU galK rpsL (SmR) endA1 l nupG + R thi-1 endA1 hsdR17 (r K m K) supE44 DlacX74 recA1 DdapAOcat (Cm ) BM2710 DmsbB DdapAOPtac-inv DdapBOPtet-hly 43 19 Unpublished data a Abbreviations used for antibiotic resistance (R): Ap ¼ ampicillin; BS ¼ blasticidin S; Cm ¼ chloramphenicol; Km ¼ kanamycin; Sm ¼ streptomycin; Sp ¼ spectinomycin; Tc ¼ tetracycline. Table 2 HT1080 clones obtained at days 17–19 with selection on BS Experiment Plasmid transferred 1 B2T8, clone U7 2 3 B2T8, clone U7 B2T8, clone U4 clone U7 clone U8 1 TTE1, clone F 2 TTE1, clone F 3 TTE1, clone F 1 CGT21, clone clone clone clone CGT21, clone clone clone clone 2 20 24 34 38 20 24 34 38 MOI Number of clones per total number of plated cells 5 25 25 25 25 25 3/3 106 B60/3 106 13/1.5 106 11/1.5 106 12/1.5 106 19/1.5 106 5 25 5 25 25 3/5 105 17/5 105 0/5 105 19/5 105 8/1.5 106 25 25 25 25 25 25 25 25 0/3 106 11/3 106 3/3 106 1/3 106 17/3 106 5/3 106 17/3 106 7/3 106 transfer of a deleted construct could result, by subsequent homologous recombination, in a final functional centromere; in fact, de novo formed HACs have been estimated to be larger than single input centromere sequences implying DNA concatemerization7,6,14,38 and arrays as small as 70 kb of contiguous sequences have proven to be sufficient to form centromeres.5 To test if bacteria allow transfer of large intact and functional genomic loci, we used an engineered genomic construct CGT21,23 which expresses a large transcriptional unit of the CFTR gene and is easily distinguishable from the endogenous CFTR transcripts due to an EGFP fusion. Results indicated that the CGT21 construct after bacterial transfer into mammalian cells is expressed from the CFTR promoter lying at a distance of 80 kb from the last of the 12 exons (exon 10 according to the CFTR Gene Therapy nomenclature),39 indicating successful transfer of a large functional unit of at least 80 kb. A long RT-PCR spanning all CGT21 exons (from the CFTR start codon to the EGFP coding region fused to CFTR exon 10) showed that this construct is stably expressed from the CFTR promoter in the mammalian HT1080 cell line and that all exons are correctly spliced. Assuming complete transfer of a given PAC construct, integration requires opening (ie linearization) of the circular molecules. If this process occurs randomly, any region of the construct, including the functional unit (ie the gene), could be interrupted during insertion. Within the 159 kb sized CGT21 construct, the functional unit covers at least 80 kb. Therefore, one would expect that less than 50% of the integrants should have an intact functional CGT21 unit. Among the 40 clones resistant to BS obtained after transfer of CGT21, 23 harboured vector and insert portions as assessed by PCR (Table 3). In all, 14 candidate clones were analysed and nine (64%) were shown to express the correctly spliced transgene, as assessed by long RT-PCR and sequencing (Figure 7, and data not shown). Since stable cell lines were isolated with selection for the BS marker spanning only 2 kb, there was no bias towards transfer of the intact 80 kb unit. Taken together, these data suggest that transfer and maintenance of an unaltered large construct is not a rare event. Gene delivery systems play a central role in the development of gene therapy strategies for monogenetic hereditary diseases, like CF, as they offer the potential of correcting the underlying cause when the responsible gene is known.35 Since delivery of a large gene, like CFTR, to the relevant cells, at the proper expression level seems difficult to achieve with available vector systems, alternative approaches should be considered. Engineered bacteria have the potential to circumvent some of the problems associated with construct/vector systems currently in use, such as the inefficiency of intact gene delivery and lack of stable expression. In particular, this system further extends the ability of introducing very large DNA sequences into phenotypically inert and nonintegrative autonomously replicating vectors such as artificial chromosomes. This is achieved by combining their inherent advantages (ie stability and large carrying Bacterial transfer into human cells A Laner et al 1567 Table 3 PCR screening of BS-resistant HT1080 clones Primers used for PCR Plasmid transferred 17-2A/T7 EGF/EGR LPF/LPR BemF/BemR KF/KR Number of clones B2T8 + NAa NA + + + + + 11/12 1/12 TTE1 NA NA NA + + + + + + + + + 9/14 4/14 1/14 CGT21 NA NA NA NA + + + + + + + + + 23/40 10/40 4/40 3/40 a NA ¼ not applicable. capacity) with the ease of purification and transferability into mammalian cells. The potential of this system is demonstrated by the design of CGT21 that could be modified by replacing EGFP by the remainder of CFTR to encode a functional CFTR or by insertion of the other elements required for HAC formation, opening new perspectives for gene therapy of CF or for any other human genomic-based cell therapy. A number of genetic techniques can be used to modify in bacteria the genomic inserts cloned in BACs and direct transfer of the engineered BAC from the host strain into the target cell can be performed without DNA purification. This approach allows functional analysis of large structural and regulatory genomic regions in cell culture assays and has been successfully adapted for the analysis and rapid phenotypic screening of the murine cytomegalovirus genome.40 Combined with the efficient transfer of low copy intact large functional DNA demonstrated here for the human CFTR locus, these data suggest that, in principle, bacteria can be engineered to transfer artificial chromosomes that express human genes. Materials and methods PAC-based plasmids The maps of the PAC vectors are shown in Figure 1. Plasmid B2T8 contains a 190 kb a-satellite DNA array of human chromosome 17 composed of 2.6 kb sized, 16meric EcoRI higher-order repeats belonging to the pentameric family of alpha satellite sequences also found on human chromosomes X and 11. It was constructed by inserting NotI cut PAC B2 (isolation described in Schindelhauer and Schwarz4) in the Bsp120I site of the ditelomeric 17 kb PAC vector pTAT-BS, which contains a BS resistance gene. End-sequence pairs of 10 randomly subcloned EcoRI higher-order repeats showed a high similarity (497%). The sequences are provided in the EMBL/Genbank database under accession numbers AJ563631–AJ563650. Partial restriction and pulsed field mapping showed the absence throughout the array of fragments other than the 2.6 kb EcoRI fragments (data not shown). De novo centromere proficiency of the plasmid was shown by intranuclear microinjection of approximately 1–10 linearized copies and by lipofection (data not shown), and similar alpha satellite arrays of chromosome 17.6,12 Plasmid TTE1 contains a 116 kb a-satellite array of chromosome 5, belonging to a subtype of the dimeric alpha satellite family, which is present on human chromosomes 1, 5 and 19.41 The homogeneous array present in TTE1 contains 0.34 kb EcoRI higher-order repeats as revealed by restriction analysis (not shown). It was isolated from PAC library RPCI 704 by array-specific PCR using primers 5IF and 5IR. The primer sequences were derived from that of the 0.7 kb insert in plasmid pZ5.1 (Hulsebos et al42; http://www.biologia.uniba.it/ rmc/5-alfoidi/alfoidiplasmids.html). The NotI insert (116 kb) of PAC E1 was cloned into the Bsp120I site of pTT (26 kb), a tetratelomeric PAC vector derived from pTAT-BS by insertional duplication of the portion between the telomeres, insertion of a prokaryotic white/blue selectable marker derived from pUC19 (Gibco BRL, Gaithersburg, MD, USA) and insertion of a eukaryotic CMV/EGFP expression cassette derived by PCR from plasmid pEGFP-N1 (Clontech, Palo Alto, CA, USA).22 Construction of the genomic CFTR expression plasmid CGT21 (EMBL/Genbank accession number BN000167) is described elsewhere.23 In brief, an unaltered PAC containing the 50 portion of the human CFTR locus (60 kb to CFTR intron 9) was joined by cloning with a synthetic exon fused from sequences of CFTR intron 9, exon 10, the EGFP coding region, CFTR exon 24 plus 30 UTR and further CFTR 30 downstream region. Sequences of the CF-PAC exons are available online from the VR of the European working group on CFTR expression (http://central.igc.gulbenkian.pt/cftr/vr/) and under Genbank/EMBL accession numbers AJ574939–AJ575055. The resulting NotI fragment of 142 kb was cloned in the Bsp120I site of pTAT-BS (17 kb) to include the BS selectable marker and for future incorporation into HACs. This PAC plasmid designated CGT21 (159 kb) was functionally analysed in stable HT1080 cell lines. RT-PCR spanning all exons revealed correct splicing of all exons, including the synthetic one (the sequence of the resulting cDNA is deposited in the EMBL/Genbank database under accession number AY299332).23 Gene Therapy 1568 Gene Therapy Table 4 Oligodeoxynucleotides used in this study Sequence (50 –30 ) Locationa 17-2A T7 ATAACTGCACCTAACTAAACG TAATACGACTCACTATAGGG EGF EGR LPF LPR BemF BemR KF KR RsF AGGGCGAGGAGCTGTTCAC GTGCGCTCCTGGACGTAGC GAAACGGCCTTAACGACGTAGTCG ATGATAAGCTGTCAAACATGAGAATTG CATGCTCACGGCAATGCCGG TGGCACTTTGCGTATCGTCCA GGGAAAACAGCATTCCAGGTATTAG CCATGAGTGACGACTGAATCCGGT AGCGGTCGGACCGTGCTC B2T8, chr. 17 a-sat DNA, present in the 2.6 kb higher-order repeats Cloning boundary of the NotI insert of chr.17 a-sat DNA derived from PAC B2 TTE1, CGT21, EGFP cDNA fragment 5IF GTGAGGAAACAGTCTGTTTGTC 5IR 17-1A Size of the PCR product (bp) Reference Ca. 150 45 This work, 4 PCR 281 This work, 23 PCR B2T8, TTE1, CGT21, vector fragment, adjacent to the loxP site 109 This work, 23 PCR B2T8, TTE1, CGT21, vector fragment, close to Bsp120I 159 This work, 23 PCR 381 This work PCR pTAT-BS vector fragment, includes the loxP site and the BS selectable marker 3160 This work, 23 TTE1; chr. 5 a-sat DNA type I, present in the 0.34 kb higher-order repeats 275 41,22 GAATCATTCTGTCTAGTTTTTATAC TGTTTAGTCAGCTGAAATT B2T8; chr. 17 a-sat DNA, present in the 2.6 kb higher-order repeats 45 A1R CGAGAGACCATGCAGAGGTC CFTR exon 1 (forward) C16D GFP1-AL B3F ActS ActAS AC1L CFex5.F B2R GTTGGCATGCTTTGATGACGCTTC GAACTTCAGGGTCAGCTTG AATGTAACAGCCTTCTGGGAG GCACTCTTCCAGCCTTCC GCGCTCAGGAGGAGCAAT GAAACCAAGTCCACAGAAGGC CTCCTTTCCAACAACCTGAAC GGAAGGCAGCCTATGTGAGA Endogenous CFTR exon 10 (reverse) Synthetic exon 10 in CGT21, EGFP portion (reverse) CFTR exon 8 (forward) b-Actin (forward) b-Actin (reverse) CFTR exon 6a CFTR exon 5 CFTR exon 7 Ca. 1900 with 17-2A 1668 with GFP1-AL 391 with B3F 474 with B3F a KmR ¼ resistance to kanamycin; BSR ¼ resistance to blasticidin S. B2T8, TTE1, CGT21, KmR fragment R 228 — — — 49 47 23 47 48 48 This work This work 49 Used for Probe for Southern and FISH Probe for FISH Probe for FISH RT-PCR RT-PCR RT-PCR RT-PCR RT-PCR RT-PCR Sequencing Sequencing Sequencing Bacterial transfer into human cells A Laner et al Primer Bacterial transfer into human cells A Laner et al Bacterial strains, plasmids and growth conditions The bacterial strains and plasmids used in this study are listed in Table 1. Bacteria were grown in brain heart infusion (BHI, Difco Laboratories, Detroit, MI, USA) broth or agar. The dap auxotroph strains E. coli BM2710 and BM4573 (a BM2710 derivative in which the inv and hly genes have been inserted in the chromosome, manuscript in preparation) were grown with 0.5 mM of diaminopimelic acid (dap, Sigma, St Louis, MO, USA). Bacteria harbouring pGB2Oinv-hly were selected with 25 mg/ml of spectinomycin. Bacteria harbouring B2T8 and TTE1 were selected with both 25 mg/ml of kanamycin, 50 mg/ml of ampicillin; bacteria harbouring CGT21 were selected with kanamycin, ampicillin and 2 mg/ml of tetracycline. Ampicillin (50 mg/ml) was used to select strains containing pAT505. Electroporation Overnight cultures of E. coli BM2710/pGB2Oinv-hly or E. coli BM4573 were diluted 1:50 in 40 ml of BHI broth containing 0.5 mM dap. Bacteria were grown at 371C with shaking to an OD600 of 0.5–0.6, chilled 15 min on ice, harvested by centrifugation for 5 min at 6000 r.p.m. at 41C, washed twice in 10 ml of cold sterile water and collected by centrifugation for 5 min at 6000 r.p.m. at 41C. The bacteria were resuspended in 10 ml of 10% cold glycerol, centrifuged 10 min at 6000 r.p.m. and the pellet was resuspended in 0.2 ml of 10% glycerol. Circular DNA of PAC plasmids B2T8, TTE1 and CGT21 was isolated from E. coli DH10B43 in agarose plugs containing approximately 1 mg/100 ml of DNA. From 30 to 50 mg of low melting point agarose containing plasmid DNA was melted by incubation at 651C for 10 min with 1/10 volume of 10 b-Agarase I buffer (New England BioLabs, Beverly, MA, USA). The melted agarose was cooled to 401C and incubated from 1 to 1 h 30 with bAgarase (one unit for 100 ml). Electrocompetent bacteria (25 ml) and 3 ml of plasmid DNA were added to microcentrifuge tubes placed on ice, mixed and transferred to cuvettes with a 0.1 cm gap (Bio-Rad Laboratories, Richmond, CA, USA) chilled on ice. Electroporation (Bio-Rad gene Pulser) conditions were 100 O, 1.6 kV and 25 mF. A measure of 1 ml of BHI medium with 0.5 mM dap was immediately added to the cuvette and the content was then transferred to sterile glass cultured tubes for growth for 1 h with moderate shaking at 371C. Bacteria (0.25 ml) were spread on BHI agar containing 0.5 mM dap and appropriate antibiotics, and the plates were incubated for a minimum of 24 h at 371C. Analysis of plasmid DNA by pulsed field gel electrophoresis (PFGE) To verify integrity of the PAC plasmids after transfer in the bacteria, agarose plugs were prepared from E. coli BM2710 harbouring pGB2Oinv-hly+B2T8 or CGT21, E. coli BM4573 harbouring TTE1 and from parental E. coli DH10B strain. Circular DNA was purified after fragmentation of E. coli chromosome by restriction digestion and PFGE purification as described.44 The size of the plasmids was determined by I-SceI cleavage, removing vector sequences outside of the telomeric repeats, and PFGE with a CHEF DRII apparatus (Bio-Rad Laboratories, Richmond, CA, USA) with the following settings: 6 V/cm, ramped switch time 1–30 s, 16 h run in 0.5 TAE buffer at 121C. The gels were stained with ethidium bromide and the bands visualized under UV light. 1569 Bacterial invasion of cells The HT1080 (human lung sarcoma) cell line was maintained in complete culture Dulbecco’s modified Eagle’s medium (DMEM; ICN Biomedicals, Aurora, OH, USA) supplemented with 2 mM L-glutamine and 10% fetal calf serum (FCS, Gibco-BRL, Gaithersburg, MD, USA). The cells were incubated overnight in six-well plates (2.5 105 cells/well) or in 100 mm Petri dishes (0.65 105–1.5 106 cells/dish). Bacteria were grown overnight at 301C in BHI broth supplemented with 0.5 mM dap and the appropriate antibiotics, harvested by centrifugation and resuspended in DMEM containing 0.5 mM dap at 2.5 108 bacteria/ml. Bacteria were added to HT1080 cells in 2 ml (six-well plates) or 6 ml (100 mm Petri dishes) of DMEM with dap to obtain an MOI of ca. 5 or 25 and the plates or dishes were incubated for 2 h at 371C. The cells were then washed three times with DMEM and incubated in complete medium containing 20 mg/ml of gentamicin to remove extracellular bacteria for 2 days. Generation of stable BS-resistant HT1080 clones The infected cells were then incubated in complete medium containing 4 mg/ml of BS (ICN Biochemicals, Aurora, OH, USA) for at least 2 weeks and individual resistant HT1080 clones (containing 250–1000 cells) were isolated using cloning cylinders (Sigma, St Louis, MO, USA). Colonies were trypsinized and transferred into six-well plates and subsequently expanded in 25 and 75 cm2 flasks for analysis. Counts of internalized bacteria After invasion, cells were incubated 30 min at 371C in complete medium containing 20 mg/ml of gentamicin and washed three times with DMEM. The bacteria were released from the cells with 0.25% deoxycholate, and viable counts were determined on BHI agar plates containing 0.5 mM dap and antibiotics. Flow cytometric analysis HT1080 cells were trypsinized, washed once with 2% FCS in phosphate-buffered saline (PBS), resuspended in the same medium at 106 cells/ml and 3 104 cells were analysed by flow cytometry using a FACScan flow cytometer with CellQuest software (Becton-Dickinson, Mountain View, CA, USA). PCR screening of resistant HT1080 clones Cell clones were screened by PCR using primer pairs described in Table 4. Cells (1–2 106) were trypsinized, and washed once with 5 ml of PBS; the pellet was resuspended in 50–100 ml of TTE buffer (0.01% Triton X-100, 20 mM Tris-HCl (pH 8), 2 mM EDTA, pH 8), incubated 10 min at 1001C and centrifuged for 3 min at 10 000 r.p.m. A measure of 2 ml of supernatant was used in a final volume of 50 ml for a PCR of 35 cycles with annealing temperatures of 601C (LPF/LPR; BemF/BemR; KF/KR; EGF/EGR) or 531C (17-2A/T7). Untransfected HT1080 cells were used as a negative control. Gene Therapy Bacterial transfer into human cells A Laner et al 1570 Southern hybridization Genomic DNA of stable cell lines was prepared using pronase A, salt precipitation of proteins and ethanol precipitation. Approximately 3 mg of genomic DNA was digested with HindIII and separated on a 1% agarose gel at 30 V for 16 h in TBE buffer and probed with the 3.15 kb PCR fragment obtained with Rsf primer (Table 4) spanning the BS resistance marker. The PCR product was labelled using random primed polymerization with Klenow enzyme (Roche, Mannheim, Germany) and 32PdCTP (Amersham, UK). Hybridization was carried out at 651C overnight in the presence of dextransulphate and salmon sperm DNA. FISH analysis Cells enriched for metaphases using 0.4 mg/ml colcemid (Roche, Mannheim, Germany) for 4 h were treated with hypotonic solution at 371C for 40 min in 0.8% (w/v) sodium citrate. Nuclei were fixed in methanol/acetic acid (3:1) at 201C for standard chromosomal spreads. RNase A- (Roche, Mannheim, Germany) and pepsin(Boehringer, Mannheim, Germany) treated chromosome preparations were dehydrated in ethanol. Chromosomal DNA and probes were simultaneously denaturated at 721C for 5 min in 50% (v/v) formamide, 2 SSC, 20% (w/v) dextran sulphate and incubated in a humid chamber at 371C for 72 h. FISH hybridization was carried out in the presence of 1 mg/ml salmon testes DNA (Sigma, St Louis, MO, USA). Posthybridization washes were carried out as described.23 After blocking with 3% (w/v) BSA, Cy3.5-conjugated Avidin (Rockland, Gilbertsville, PA, USA) or FITC-conjugated anti-dig antibody (Roche, Mannheim, Germany) was bound in 1% (w/v) BSA, 4 SSC and 0.2% (v/v) Tween-20. Slides were washed (4 SSC, 0.2% Tween-20, 421C) and counterstained with DAPI (40 ,6-diamidino-2-phenylindole; Sigma, St Louis, MO, USA), mounted with antifade (PPD) and analysed on a Leica DM RXA with a CCD camera controlled by Q-FISH software (Leica Microsystems, Switzerland). The vector probe was obtained by amplifying a 3.15 kb fragment with primer Rsf (Table 4) from pTAT-BS. The product was labelled by PCR incorporation of biotin-16-dUTP (Roche, Mannheim, Germany). The centromeric probes cen 5I and cen 17 were amplified using primers 5IF/5IR and 17-1A/17-2A (Table 4), respectively, from the corresponding PACs. 5IF/5IR amplified a 275 bp region including the CENP-B box of the 0.34 kb higher-order repeat of alpha satellite array type I of chromosome 5 (Z5.1, Archidiacano et al41), and 17-1A/17-2A amplified a 1.9 kb segment of the 2.6 kb higher-order repeats of the alpha satellite DNA array of chromosome 17.45 RT-PCR analysis Total RNA was isolated from BS-resistant cell lines using the RNeasy extraction kit (Qiagen, Hilden, Germany) according to the manufacturer’s instructions. RT was carried out using random hexamers and Superscript II RnaseH-reverse transcriptase (Invitrogen, Paisley, UK) as described.46 The cDNA obtained was used as a template in a long RT-PCR to assess expression and correct splicing of the CGT21 construct. The reaction was performed between CFTR exon 1 and CGT21 synthetic fusion of CFTR exon 10-EGFP using primers A1R (CFTR Gene Therapy exon 1) and GFP1-AL (EGFP sequence at the CGT21 fusion) (Table 4). The expected size of the correctly spliced RT-PCR product was 1668 bp. Each reaction contained 10 ml of cDNA and 1 U of AmpliTaq DNA Polymerase (Applied Biosystems, Foster city, CA, USA); 1 buffer I (Applied Biosystems); 200 mM of each dNTP (Amersham Bioscience, Uppsala, Sweden); and 0.2 mM of each primer in a final volume of 50 ml. In all, 40 amplification cycles were used as follows: denaturation at 941C for 1 min, annealing at 581C for 1 min and extension at 721C for 3 min. A final extension at 721C was carried out for 12 min. Efficiency of RNA extraction and the RNA levels in the various samples were controlled by RT-PCR with 2 ml of each cDNA sample to amplify a 228 bp b-actin fragment using primers ActS (forward) and ActAS (reverse) (Table 4) at an annealing temperature of 581C. To assess the relative levels of transcripts from the CGT21 construct and from endogenous CFTR, two additional RT-PCR reactions were performed. The first of these reactions used primers B3F (CFTR exon 8) and GFP1-AL (described in Table 4) amplifying a fragment of 471 bp, specific of CGT21 transcripts. For the second reaction, amplifying a 391 bp fragment specific of endogenous CFTR, we used primers B3F (CFTR exon 8) and C16D (in region of CFTR exon 10 that is absent in CGT21, see also Table 4). A measure of 5 ml of each cDNA sample was added to the PCR reaction mixture, which was heated at 941C for 5 min, and then subjected to 35 cycles of denaturation at 941C for 1 min, annealing at 601C for 1 min and extension at 721C for 2 min, followed by a final extension at 721C for 12 min. The RT-PCR products were analysed by 2% (w/v) agarose gel electrophoresis. The gel images were registered on an UV DBT-08 digital analyser (UVItec Ltd, Cambridge, UK). Products from the long RT-PCR amplification were purified using the Jet Quick PCR product purification spin kit from Genomed (Löhne, Germany) and sequenced with the ABI PRISMt Dye Terminator Cycle sequencing system V1.1 (Applied Biosystems), according to the manufacturer’s instructions. Three primers (see Table 4) were used in different sequencing reactions to obtain the complete sequence of the long RT-PCR products, namely: AC1L (reverse primer located in CFTR exon 6a), CFex5.F (forward primer located in CFTR exon 5) and B2R (forward primer located in CFTR exon 7). Sequences of these three products were obtained on an ABI Prismt 3100 capillary electrophoresis automatic sequencer (Applied Biosystems). Acknowledgements This work was supported in part by two grants from l’association Vaincre la Mucoviscidose in 2002–2003 and 2003–2004, the latter together with the German Association Mukoviszidose e.v. D Schindelhauer was supported by the Deutsche Forschung Gemeinschaft and MD Amaral by research Grant POCTI/1999/MGI/35737 from FCT Portugal. AS Ramalho was a recipient of PhD fellowship SFRH/BD/3085/2000 from FCT, Portugal. We thank C Klein and M Speicher for FISH equipment. Bacterial transfer into human cells A Laner et al 1571 References 1 Shizuya H et al. Cloning and stable maintenance of 300-kilobasepair fragments of human DNA in Escherichia coli using an F-factor-based vector. Biochemistry 1992; 89: 8794–8797. 2 Ioannou PA et al. A new bacteriophage P1-derived vector for the propagation of large human DNA fragments. Nat Genet 1994; 6: 84–89. 3 International Human Genome Mapping Consortium. A physical map of the human genome. Nature 2001; 409: 934–941. 4 Schindelhauer D, Schwarz T. Evidence for a fast, intrachromosomal conversion mechanism from mapping of nucleotide variants within a homogeneous alpha-satellite DNA array. Genome Res 2002; 12: 1815–1826. 5 Ebersole TA et al. Mammalian artificial chromosome formation from circular alphoid input DNA does not require telomere repeats. Hum Mol Genet 2000; 9: 1623–1631. 6 Harrington JJ et al. Formation of de novo centromeres and construction of first-generation human artificial microchromosomes. Nat Genet 1997; 15: 345–355. 7 Ikeno M et al. Construction of YAC-based mammalian artificial chromosomes. Nat Biotechnol 1998; 16: 431–439. 8 Grimes BR, Rhoades AA, Willard HF. Alpha-satellite DNA and vector composition influence rates of human artificial chromosome formation. Mol Ther 2002; 5: 798–805. 9 Henning KA et al. Human artificial chromosomes generated by modification of a yeast artificial chromosome containing both human alpha satellite and single-copy DNA sequences. Proc Natl Acad Sci USA 1999; 96: 592–597. 10 Schueler MG et al. Genomic and genetic definition of a functional human centromere. Science 2001; 294: 109–115. 11 Kouprina N et al. Cloning of human centromeres by transformation associated recombination in yeast and generation of functional human artificial chromosomes. Nucleic Acids Res 2003; 31: 922–934. 12 Meija JE et al. Efficiency of de novo centromere formation in human artificial chromosomes. Genomics 2002; 79: 297–304. 13 Ohzeki J, Nakano M, Masumoto H. CENP-B box is required for de novo centromere chromatin assembly on human alphoid DNA. J Cell Biol 2002; 159: 765–775. 14 Meija JE et al. Functional complementation of a genetic deficiency with human artificial chromosomes. Am J Hum Genet 2001; 69: 315–326. 15 Grimes BR et al. Stable gene expression from a human artificial chromosome. EMBO Rep 2001; 21: 910–914. 16 Ikeno M et al. Generation of human artificial chromosomes expressing naturally controlled guanosine triphosphate cyclohydrolase I gene. Genes Cell 2002; 7: 1021–1032. 17 Wade-Martins R et al. An infectious transfer and expression system for genomic DNA loci in human and mouse cells. Nat Biotechnol 2001; 19: 1067–1070. 18 Magin-Lachmann C et al. In vitro and in vivo delivery of intact BAC DNA – comparison of different methods. J Gen Med 2004; 6: 195–209. 19 Courvalin P, Goussard S, Grillot-Courvalin C. Gene transfer from bacteria to mammalian cells. CR Acad Sci 1995; 318: 1207–1212. 20 Grillot-Courvalin C et al. Functional gene transfer from intracellular bacteria to mammalian cells. Nat Biotechnol 1998; 16: 862–866. 21 Narayanan K, Warburton PE. DNA modification and functional delivery into human cells using Escherichia coli DH10B. Nucleic Acids Res 2003; 31: e51. 22 Laner A, Schwarz T, Christan S, Schindelhauer D. Suitability of a CMV/EGFP cassette to monitor stable expression from human artificial chromosomes but not transient transfer in the cells forming viable clones. Cytogenet Genome Res 2004; 107: 9–13. 23 Schindelhauer D et al. An engineered genomic CFTR construct is expressed and correctly spliced in the human lung sarcoma cell line HT1080. (submitted). 24 Bachmann BJ. Derivations and genotypes of some mutant derivatives of Escherichia coli K-12. In: Brooks LK, Ingraham JL, Magasanik B, Neidhardt FC, Schaechter M, Umbarger HE (eds). Escherichia coli and Salmonella Typhimurium, Cellular and Molecular Biology. ASM: Washington, 1987, pp 1190–1219. 25 Grillot-Courvalin C, Goussard S, Courvalin P. Bacteria as gene delivery vectors for mammalian cells. Curr Opin Biotechnol 1999; 10: 477–481. 26 Weiss S, Chakraborty T. Transfer of eukaryotic expression plasmids to mammalian host cells by bacterial carriers. Curr Opin Biotechnol 2001; 12: 467–472. 27 Sizemore DR, Branstrom AA, Sadoff JC. Attenuated Shigella as a DNA delivery vehicle for DNA-mediated immunization. Science 1995; 270: 299–302. 28 Fajac I et al. Recombinant Escherichia coli as a gene delivery vector into airway epithelial cells. J Control Rel 2004; 97: 371–381. 29 Darji A et al. Oral somatic transgene vaccination using attenuated S. typhimurium. Cell 1997; 91: 765–775. 30 Dietrich G et al. Delivery of antigen-encoding plasmid DNA into the cytosol of macrophages by attenuated suicide Listeria monocytogenes. Nat Biotechnol 1998; 16: 181–185. 31 Paglia P et al. Gene transfer in dendritic cells, induced by oral DNA vaccination with Salmonella typhimurium, results in protective immunity against a murine fibrosarcoma. Blood 1998; 92: 3172–3176. 32 Paglia P et al. In vivo correction of genetic defects of monocyte/ macrophages using attenuated Salmonella as oral vectors for targeted gene delivery. Gene Therapy 2000; 7: 1725–1730. 33 Castagliuolo I et al. Engineered Escherichia coli deliver therapeutic genes to the colonic mucosa of mice. Gene Therapy 12: 1070–1078. 34 Lipps HJ et al. Chromosome-based vectors for gene therapy. Gene 2003; 304: 23–33. 35 Klink D et al. Gene delivery systems-gene therapy vectors for cystic fibrosis. J Cystic Fibrosis 2004; 3: 203–212. 36 Narayanan K et al. Efficient and precise engineering of a 200 kb b-globin human/bacterial artificial chromosome in E. coli DH10B using an inducible homologous recombinant system. Gene Therapy 1999; 6: 442–447. 37 Grillot-Courvalin C, Goussard S, Courvalin P. Wild-type intracellular bacteria deliver DNA into mammalian cells. Cell Microbiol 2002; 4: 177–186. 38 Rudd MK, Mays RW, Schwartz S, Willard HF. Human artificial chromosomes with alpha satellite-based de novo centromeres show increased frequency of nondisjunction and anaphase lag. Mol Cell Biol 2003; 23: 7689–7697. 39 Zielenski J et al. Genomic DNA sequence of the cystic fibrosis transmembrane conductance regulator (CFTR) gene. Genomics 1991; 10: 214–228. 40 Brune W, Ménard C, Heesemann J, Koszinowski UH. A ribonucleotide reductase homolog of cytomegalovirus and endothelial cell tropism. Science 2001; 291: 303–305. 41 Archidiacano N et al. Comparative mapping of human alphoid sequences in great apes using fluorescence in situ hybridization. Genomics 1995; 25: 477–484. 42 Hulsebos TJ et al. Assignment of the beta B1 crystallin gene (CRYBB1) to human chromosome 22 and mouse chromosome 5. Genomics 1995; 29: 712–718. 43 Grant SGN, Jessee J, Bloom FR, Hanahan D. Differential plasmid rescue from transgenic mouse DNAs into Escherichia coli methylation-restriction mutants. Genetics 1990; 87: 4645–4649. 44 Schindelhauer D, Cooke HJ. Efficient combination of large DNA in vitro: in gel site specific recombination (IGSSR) of PAC fragments containing alpha satellite DNA and the human HPRT gene locus. Nucleic Acids Res 1997; 25: 2241–2243. Gene Therapy Bacterial transfer into human cells A Laner et al 1572 45 Warburton PE, Willard HF. Genomic analysis of sequence variation in tandemly repeated DNA. Evidence for localized homogeneous sequence domains within arrays of a-satellite DNA. J Mol Biol 1990; 216: 3–16. 46 Ramalho AS et al. Methods for RNA extraction, cDNA preparation and analysis of CFTR transcripts. J Cystic Fibrosis 2004; 3: 11–15. 47 Ramalho AS et al. Five percent of normal cystic fibrosis transmembrane conductance regulator mRNA ameliorates the Gene Therapy severity of pulmonary disease in cystic fibrosis. Am J Respir Cell Mol Biol 2002; 27: 619–627. 48 Mickle JE et al. A mutation in the cystic fibrosis transmembrane conductance regulator gene associated with elevated sweat chloride concentrations in the absence of cystic fibrosis. Hum Mol Genet 1998; 7: 729–735. 49 Chalkley G, Harris AA. Lymphocyte mRNA as a resource for detection of mutations and polymorphisms in the CF gene. J Med Genet 1991; 28: 777–780.