Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
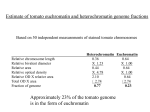
8th International Symposium on Cytochrome P450 Biodiversity and Biotechnology Dwarfism and cytochrome P450-mediated C-6 oxidation of plant steroid hormones G. Bishop*1 , T. Nomura†, T. Yokota‡, T. Montoya§, J. Castle§, K. Harrison*, T. Kushiro†, Y. Kamiya†, S. Yamaguchi†, S. Bancos, A.-M. Szatmári and M. Szekeres *Division of Biology, Imperial College London, Wye Campus, Kent TN25 5AH, U.K., †Plant Science Center, RIKEN, Institute of Physical and Chemical Research, Tsurumi-ku, Yokohama, Kanagawa 230-0045, Japan, ‡Department of Biosciences, Teikyo University, Utsunomiya 320-8551, Japan, §Institute of Biological Sciences, University of Wales, Aberystwyth SY23 3DD, U.K., and Institute of Plant Biology, Biological Research Centre of the Hungarian Academy of Sciences, P.O. Box 521, H-6701 Szeged, Hungary Abstract BRs (brassinosteroids) are plant steroid hormones that are essential for normal plant development. The dramatic dwarfism exhibited by mutants in the CYP (cytochrome P450) enzymes involved in BR biosynthesis indicates a role for these hormones in plant growth and development. Since the mid-1990s, collaborative research has been geared towards developing a better understanding of the CYP85 class of CYPs involved in BR biosynthesis in both Arabidopsis and tomato. Some of the most recent observations include the fact that certain CYP85 CYPs catalyse the synthesis of the most bioactive BR, BL (brassinolide). Current evidence suggests that evolution of this function may have occurred independently in different dicotyledonous species. Interestingly, BL accumulates in tomato fruits, highlighting a key role for this hormone in fruit development. At the same time as developing a better understanding of the enzymatic function of these CYPs, we have also carried out experiments towards characterizing where and when these genes are expressed and mechanisms of their regulation. As expected for a hormone involved in growth and development, biosynthetic gene promoter activity is associated with young rapidly growing cells and with fruit development. Background CYPs (cytochrome P450s) are crucial enzymes required for hydroxylation/oxidation reactions in both mammalian and plant steroid hormone synthesis. Since the mid-1990s, rapid advances have been made in our knowledge and understanding of the biosynthesis and signalling of BRs (brassinosteroids; plant steroid hormones). The key discoveries that stimulated the BR research were the identification of mutants involved in BR synthesis and signalling. These mutants provided a role for the most bioactive BR BL (brassinolide) that had been isolated from rape pollen [1]. The structure of BL is similar to the mammalian steroid hormones [2]; however, bioactive BRs are polyhydroxylated on both the A ring and the side chain (Figure 1). The importance of the role of BRs in plant growth and development is easily observed in the striking phenotype of the dwarf mutants that lack BR synthesis and response. These mutants are generally dark green and dwarfed in all stages of plant growth and development. The dwarfism indicates BR’s involvement in the regulation of growth, photomorphogenesis and fertility. This micro-review will focus on a brief description of the tomato and Arabidopsis CYP85 CYPs involved in C-6 oxidation of BRs; please refer to Fujioka Key words: Arabidopsis, brassinolide, brassinosteroid, cytochrome P450, dwarfism, plant steroid hormone. Abbreviations used: BL, brassinolide; BR, brassinosteroid; BR1, brassinosteroid-insensitive 1; CS, castasterone; CYP, cytochrome P450; 6-deoxoCS, 6-deoxocastasterone. 1 To whom correspondence should be addressed (email [email protected]). and Yokota [3] or Szekeres and Bishop [4] for more extensive reviews of BR biosynthesis and metabolism. CYP85A genes and function The tomato DWARF gene was isolated via targeted transposon mutagenesis and represented the first member of a new class of CYPs (CYP85) [5]. The extreme dwarf (dx ) and transposon-tagged alleles exhibit severe dwarfism having dark green and crinkled leaves [5]. Yeast that heterologously expresses CYP85A was fed various BR intermediates as substrates and shown to convert 6-deoxoCS (6-deoxocastasterone-) into CS (castasterone) [6]. This conversion was consistent with the analysis of BR intermediate concentrations in vegetative tissue of the dx mutant [6]. However, more recent BR quantification of fruits from the dx mutant and wild-type indicates the production of BL in both samples [7,8]. A new CYP85A family gene, named CYP85A3, that is expressed preferentially in tomato fruits was isolated [7]. Functional expression of CYP85A3 in yeast indicated that this enzyme catalyses the Baeyer–Villiger oxidation responsible for conversion of CS into BL, in addition to C-6 oxidation of 6deoxoCS to CS [7]. In Arabidopsis, two CYP85A genes, CYP85A1 and CYP85A2, are present in the genome. The T-DNA (transfer DNA) insertion lines in CYP85A1 exhibit a normal phenotype but those of CYP85A2 show a weak dwarfism [7,9,10]. The loss-of-function cyp85a1/cyp85a2 double mutant causes severe dwarfism and sterility, indicating that, unlike tomato, C 2006 Biochemical Society 1199 1200 Biochemical Society Transactions (2006) Volume 34, part 6 Figure 1 CYP85-mediated synthesis of BL Schematic highlighting the conversion of 6-deoxoCS into BL. Figure 2 Phylogenetic tree of CYP85 genes Phylogenetic tree of selected CYP85 full-length protein sequences R present in GenBank Nucleotide Sequence Database and rooted using the CYP90A1 gene from Arabidopsis. The tree was made using MEGA 3.1 program and CLUSTAL W sequence alignment. The tree used the neighbour-joining method (Bootstrap value, 500) and Poisson correction. to verify these biological activities using purified enzymes, the results suggest that either the BL synthase function has independently evolved several times or that this function has been lost several times. Discerning the function of the rice CYP85 that is present in the genome as a single copy will hopefully clarify the ancestral function of these CYPs. CYP85A expression the roles of the two Arabidopsis CYP85A genes are overlapping in growth and development. A functional assay of the Arabidopsis CYP85A1 in yeast indicated that it catalyses BR C-6 oxidation in the conversion of 6-deoxoCS into CS [11]. The Arabidopsis CYP85A2 has recently been shown to be a BL synthase, similar to the tomato CYP85A3 protein [7,9]. Both CYP85A1 genes of Arabidopsis and tomato when expressed in yeast also catalyse the C-6 oxidation of intermediates in the upstream pathway of the BR biosynthesis [11]. Lines that are defective in both CYP85A1 and CYP85A2 function do not exhibit substantial changes in 6-oxocampestanol levels [10], which indicates that C-6 oxidation of campestanol is unlikely to be catalysed by a CYP85A enzyme. CYP85 evolution There has been differential expansion and loss of CYP families in plants with the CYP85 family being no exception. Using selected CYP85 sequences, a phylogeny tree was generated (Figure 2). This tree indicates a highly novel finding in that the CYP85s of tomato, pea and Arabidopsis (dicotyledonous plants) seem to have duplicated independently, whereas in monocotyledonous plants there is only one gene. This independent duplication also results in the generation of a novel function, with the CYP85A3 and CYP85A2 genes from tomato and Arabidopsis encoding the Baeyer–Villiger BL synthase function. Although further investigation is required C 2006 Biochemical Society The transcriptional regulation of CYP85 genes plays a crucial role in determining BR levels. Bioactive BRs facilitate the down-regulation of transcript levels of BR-biosynthesis genes thereby maintaining an optimal level of bioactive BR production. Such transcriptional feedback regulation of CYP85 CYPs is the major mechanism involved in altering BR synthesis. In Arabidopsis, CYP85A1 and CYP85A2 transcripts are down-regulated by bioactive BRs within 2 h of BL treatment [12–14]. A similar regulation has been seen for the tomato CYP85A1 gene [8]. The feedback regulation requires a functional BRI1 (brassinosteroid-insensitive 1) receptor [8,14–16] because mutants defective in the BRI1 receptor lack feedback regulation that results in very high CYP transcript level and BR accumulation [8,17,18]. In addition to the feedback control, organ-specific variation in transcript levels is observed [7,8,12,14]. The spatial regulation of gene expression provides a mechanism of altering the BR levels in different organs. The CYP85A2 transcripts are at higher levels in shoots than in roots of Arabidopsis seedlings [14,19]. This is consistent with the fact that 6deoxoCS and CS are at relatively higher concentrations in Arabidopsis and tomato shoots compared with those in roots, which have increased levels of 6-deoxotyphasterol and BR intermediates upstream in the pathway [14,20]. As discussed above, tomato fruits have been shown to exhibit preferential expression of CYP85A3 and accumulate BL, highlighting a key function of BL in fruit development. These localized gene expression profiles coupled with the lack of BR transport, as indicated by sectored plants and grafting experiments [5,21,22], suggest that the sites of expression coincide with the sites of active BR synthesis [7,22,23]. More recent work has shown that Arabidopsis CYP85A2 and CYP90A1 transcript levels exhibit a circadian rhythm of regulation [24]. This regulation leads to increased levels of BL during the day 8th International Symposium on Cytochrome P450 Biodiversity and Biotechnology and decreased levels at night, with both the light signalling pathway and the circadian clock influencing the regulation of the transcript levels. G.B. thanks the funding agencies, Biotechnology and Biological Sciences Research Council, Royal Society, Human Frontier Research Program and the British Council, for support. Thanks also go to all co-authors and collaborators who have helped make this research possible. References 1 Mitchell, J.W., Mandava, B.N., Worley, J.F., Plimmer, J.R. and Smith, M.V. (1970) Nature 225, 1065–1066 2 Grove, M.D., Spencer, G.F., Rohwedder, W.K., Mandava, N., Worley, J.F., Warthen, J.D., Steffens, G.L., Flippenanderson, J.L. and Cook, J.C. (1979) Nature 281, 216–217 3 Fujioka, S. and Yokota, T. (2003) Annu. Rev. Plant Biol. 54, 137–164 4 Szekeres, M. and Bishop, G.J. (2006) Annu. Plant Rev. 24, 67–92 5 Bishop, G.J., Harrison, K. and Jones, J.D.G. (1996) Plant Cell 8, 959–969 6 Bishop, G.J., Nomura, T., Yokota, T., Harrison, K., Noguchi, T., Fujioka, S., Takatsuto, S., Jones, J.D.G. and Kamiya, Y. (1999) Proc. Natl. Acad. Sci. U.S.A. 96, 1761–1766 7 Nomura, T., Kushiro, T., Yokota, T., Kamiya, Y., Bishop, G.J. and Yamaguchi, S. (2005) J. Biol. Chem. 280, 17873–17879 8 Montoya, T., Nomura, T., Farrar, K., Kaneta, T., Yokota, T. and Bishop, G.J. (2002) Plant Cell 14, 3163–3176 9 Kim, T.W., Hwang, J.Y., Kim, Y.S., Joo, S.H., Chang, S.C., Lee, J.S., Takatsuto, S. and Kim, S.K. (2005) Plant Cell 17, 2397–2412 10 Kwon, M., Fujioka, S., Jeon, J.H., Kim, H.B., Takatsuto, S., Yoshida, S., An, C.S. and Choe, S. (2005) J. Plant Biol. 48, 237–244 11 Shimada, Y., Fujioka, S., Miyauchi, N., Kushiro, M., Takatsuto, S., Nomura, T., Yokota, T., Kamiya, Y., Bishop, G.J. and Yoshida, S. (2001) Plant Physiol. 126, 770–779 12 Tanaka, K., Asami, T., Yoshida, S., Nakamura, Y., Matsuo, T. and Okamoto, S. (2005) Plant Cell Physiol. 46, S216–S216 13 Goda, H., Shimada, Y., Asami, T., Fujioka, S. and Yoshida, S. (2002) Plant Physiol. 130, 1319–1334 14 Bancos, S., Nomura, T., Sato, T., Molnar, G., Bishop, G.J., Koncz, C., Yokota, T., Nagy, F. and Szekeres, M. (2002) Plant Physiol. 130, 504–513 15 Choe, S., Fujioka, S., Noguchi, T., Takatsuto, S., Yoshida, S. and Feldmann, K.A. (2001) Plant J. 26, 573–582 16 Nomura, T., Bishop, G.J., Kaneta, T., Reid, J.B., Chory, J. and Yokota, T. (2003) Plant J. 36, 291–300 17 Nomura, T., Nakayama, M., Reid, J.B., Takeuchi, Y. and Yokota, T. (1997) Plant Physiol. 113, 31–37 18 Noguchi, T., Fujioka, S., Choe, S., Takatsuto, S., Yoshida, S., Yuan, H., Feldmann, K.A. and Tax, F.E. (1999) Plant Physiol. 121, 743–752 19 Shimada, Y., Goda, H., Nakamura, A., Takatsuto, S., Fujioka, S. and Yoshida, S. (2003) Plant Cell Physiol. 44, S69–S69 20 Yokota, T., Sato, T., Takeuchi, Y., Nomura, T., Uno, K., Watanabe, T. and Takatsuto, S. (2001) Phytochemistry 58, 233–238 21 Symons, G.M. and Reid, J.B. (2004) Plant Physiol. 135, 2196–2206 22 Montoya, T., Nomura, T., Yokota, T., Farrar, K., Harrison, K., Jones, J.G.D., Kaneta, T., Kamiya, Y., Szekeres, M. and Bishop, G.J. (2005) Plant J. 42, 262–269 23 Castle, J., Szekeres, M., Jenkins, G. and Bishop, G.J. (2005) Plant Mol. Biol. 57, 129–140 24 Bancos, S., Szatmari, A.M., Castle, J., Kozma-Bognar, L., Shibata, K., Yokota, T., Bishop, G.J., Nagy, F. and Szekeres, M. (2006) Plant Physiol. 141, 299–309 Received 21 June 2006 C 2006 Biochemical Society 1201