Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
“The Essence of Life!”
Photosynthesis 6 CO2 + 6 H2O + light
C6H12O6 (as triose-P) + 6 O2
ATP
NADPH, Carbon Skeletons?
{
Reduction of CO2/oxidation of H2O are coupled to formation of ATP and
NADPH which are used in the chloroplast, mainly to make Triose-phosphate.
Respiration C12H22O11 + 12 O2
(Biological
combustion)
Fictitious sugar, for
equation balance
only!
12 CO2 + 11 H2O + heat
{
ATP
NADH
Carbon skeletons
NADPH (Pentose Phosphate)
Oxidation of sugar and reduction of O2 is linked to the formation of ATP,
NADH (for OxPhos) and NADPH (mainly for biosynthesis). An often
ignored but key product of respiration is carbon skeletons to make
biomolecules. Respiration mainly consists of: glycolysis, the pentose
phosphate shunt, and oxidative phosphorylation.
Combustion ( /kəmˈbʌs.tʃən/) or burning is the sequence of exothermic chemical reactions between a fuel and an oxidant accompanied
by the production of heat and conversion of chemical species. The release of heat can result in the production of light in the form of either
glowing or a flame. Fuels of interest often include organic compounds (especially hydrocarbons) in the gas, liquid or solid phase.
1
Required readings, 2013
1) Front Plant Sci. 2012;3:210. doi: 10.3389/fpls.2012.00210. Epub 2012 Sep 6.
Functional genomics tools applied to plant metabolism: a survey on plant
respiration, its connections and the annotation of complex gene functions.
Araújo WL, Nunes-Nesi A, Williams TC
2) Front Plant Sci. 2011; 2: 89. Published online 2011 November 28. Prepublished
online 2011 October 4. doi: 10.3389/fpls.2011.00089 PMCID: PMC3355598
Mining the Active Proteome of Arabidopsis thaliana
Renier A. L. van der Hoorn,1,* Tom Colby,2 Sabrina Nickel,3 Kerstin H. Richau,1 Jürgen
Schmidt,2 and Markus Kaiser3
3
Logan, D.C., 2006.The
Mitochondrial Compartment, J.
Exp. Bot. 57: 1225-12243.
4
biologically-
useful
5
For muscle and
cytoskeletal motors
Oxidized
NAD
Reduced NAD
Source of protons
For Ox Phos in the
mitochondria
6
Respiration is
more than just
ATP production!!
Key Metabolites
7
(pentose phosphate also)
8
The main “fuel” in plants is sucrose or starch
Glycolysis
Alcoholic fermentation (common in
plants and yeasts) is less
damaging than lactic acid
fermentation.
Much regulation in
glycolysis, sucrose,
starch biosynthesis is
apparently designed to
accommodate Pi
deprivation.
9
NOTE: The PFK block
(irreversibility) is
circumvented in plants
by plant-unique PFP
PFP
PFK
In plants there are several
additional, important variations of
glycolysis. Note the carbon feeds
from chloroplasts versus
amyloplasts and the shunts from
PEP (to produce pyruvate) and
PEP to malate. Pyruvate and
malate can be utilized in the
mitochondria for ox phos to
produce ATP.
10
Gluconeogenesis
Looks easy, but there is a critical irreversible block here
(irreversible PFK). There are several ways to get around
this in plants and animals that we don’t have time to talk
about. We only have time for the plant-unique one that
is tied to phosphate nutrition/deprivation (i.e., PFP).
Why is gluconeogenesis important to plant leaves that
can make their own food via PS?? I’ll answer with a
question: “What about a cloudy day? What if starch
made during the day is not adequate to provide the
needed surcrose for the next day or during the night?
Gluconeogenesis can then be used to tap lipids by
recycling through glycerol-P to dihydroxyacetone-P,
thru to sucrose, and acetylCoA to Pyr thru to yield
sucrose and Pi. But, lipid utilization thru
gluconeogenesis is not normally as prominent in
plants as in animals, because of photosynthesis. But,
gluconeogenesis from storage lipids in seeds is very
important to plant, so much so that it requires the
special organelle, the glyoxysome that animals do not
have. In animals oxidation of fatty acids is done in the
mitochondria.
Note: Not to be tested on in
2012
11
Plant metabolism must satisfy Pi requirements/supply
PFP- induced by
Pi starvation.
12
Complex regulation of glycolysis.
Note the multitude of regulatory
steps surrounding F2,6BP. In
plants reversible PFP makes this
regulation even more complex
than in animals.
Extensive
regulation
glycolysis and
gluconeogenesi
s in plants.
PFK
We know this much from in vitro
(mostly) studies, but these may not be
correct in some instances and
undoubtedly there is more to learn
about this regulation.
Note: Not to be tested on
in 2012
13
Note: Not to be tested
on in 2012
14
PPi
Pi
PFP
A
F-6-P
PFK B
F-1,6-P
So the PFK block to
gluconeogenesis is more
highly regulated in plants
than in animals and is tied
to Pi and PPi. Also,
remember that plants can
only ultimately eat by
photosynthesis and
gluconeogenesis.
Note: Not to be tested on
in 2012
15
16
Sucrose and
sugar
derivatives
17
NEW!! Signaling from sucrose to growth and development (including stress sensing)??
There is promise too that knowledge of T6P biology in
plants will soon deliver into tackling global concerns
about food security. Indeed, links between T6P and
plant abscisic acid stress hormone signaling as well as
the increased photosynthetic capacity upon
overexpression of TPS may allow engineering plants
that can overcome periods of water shortage and/or
result in higher yields under non-optimal conditions.
Front Plant Sci. 2011; 2: 90. How Do Sugars Regulate
Plant Growth? Matthew Paul and Patrick Van Dijck
Trehalose biosynthesis and its role in carbohydrate metabolism. Trehalose-6-phosphate (T6P) is
synthesized from UDP-glucose (UDPG) and glucose-6-P (G6P) by the activity of trehalose-6-phosphate
synthase (TPS) and subsequently converted to trehalose by trehalose-6-phosphate phosphatase (TPP).
trehalase1 (TRE1) hydrolyzes trehalose into two molecules of glucose. T6P plays a central role in regulating
sugar metabolism in plants. The precursors of T6P are derived from the sucrose metabolism. It has been
suggested that T6P is transported by an unknown mechanism into plastids where it induces starch synthesis
via thioredoxin-mediated activation of AGPase. T6P might be converted into trehalose, which has been
shown to regulate starch breakdown in plastids. Several TPPs (marked with an asterisk) have been predicted
to localize to plastids, but this still needs to be confirmed experimentally. SnRK1, which represses plant
growth, is inhibited by T6P. A regulatory loop, which involves T6P, SnRK1, and bZIP11, and that is thought to
control sucrose availability and utilization, has been proposed.
Front Plant Sci. 2011; 2: 70. Trehalose-6-Phosphate: Connecting Plant Metabolism and Development
Jathish Ponnu,1 Vanessa Wahl,2 and Markus Schmid1,*
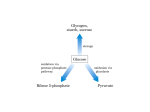
Pentose Phosphate
Metabolism
plants
19
Some internediates
of the pentose
phosphate pathway
are also in the Calvin
Cycle.
Erythrose-4-P plus PEP gives 3deoxy-D-arabino-heptulosonate-7-P,
the precursor of most naturally
occuring aromatic compounds (vis
the shikimate pathway).
Theoritically these products
could be cycled back into
glycolysis or the MEP pathway
20
Carbon skeletons
to plant
secondary
products
Phytoalexins:
Isoflavanoids, tannins,
lignin precursors from
the phenylpropanoid
path
MEP pathway
MVA pathway
21
Two pathways for Terpenoid Biosynthesis
1)
Chloroplastic MEP or GAP/G3P path
ubiquinones (mitochondrion?), dolichols, chlorophyll,
carotenoids, monoterpenes, diterpenes, tetraterpenes, GA, abscisic acid, some prenylated
proteins
2)
Cytosolic Mevalonate pathway
proteins
sesquiterpenes, sterols, triterpenes, some prenylated
Note: Not to
be tested on
in 2012
Biochemistry and Molecular Biology of Plants
Buchannan, Gruissem, Jones, 2000
22
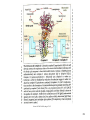
Into the Plant
Mitochondrion
Remember, plants can only eat by PS!!
Speculation with some
evidence: glycolytic
multienzyme complex with
pentose phosphate
multienzyme complex
23
Also, NADPH produced by
pentose phosphate pathway.
Remember the possibility for
NADPH production between
G-3-P and 3-PG (on pg 11,
under Pi deprivation).
24
25
26
Part of the
electron transport
chain
27
Biochemistry and Molecular Biology of Plants
Buchannan, Gruissem, Jones, 2000
28
Science 299:700-704 2003, and pp 671-672
29
The Respiratory
Chain
30
31
32
Pumped plus chemical
protons
I
III
II
IV
33
34
New data suggests that AO may be important for maintaining growth rate homeostasis
by strictly controling phosphorylation potential. TIPS 7:478-481, 2002
35
Components of respiration working in concert
germ tube
36
NADH & NADPH
dehydrogenases
37
38
MEP
pathway
Phytoalexins:
Isoflavanoids, tannins,
lignin precursors from
the phenylpropanoid
path
So, when Ca increases the pentose phosphate path
allows carbon flow through PEP to the shikimate path,
lignin, terpenes despite a decrease in glycolysis and TCA
NAD Kinase
NAD+ATP
NADP+ADP
NADH & NADPH
dehydrogenases
Also, increase in
dehydrogenase allows
use of NADPH from
pentose phosphate for
ox phos
Possible control points for
calcium in cytoplasmic
metabolic pathways
39
40
41
42
Beyond photosynthesis and photorespiration, plants are unique in that:
-More of a need for different carbon structures (secondary metabolites).
- More need to shift carbon flow to produce “fight” molecules: so more regulation of glycolysis, etc.
- Need to coordinate PS and respiration (also perhaps photorespiration that takes 25% of PS fixed carbon).
- Plants must be concerned with Pi deprivation so there is added regulation to insure basic Pi supply:
-PEP to Pi
-PPI to Pi via PFP
-PWEP to OAA to malate (releases Pi
-Pi and PPi regulate PFK2 to regulate F2,6BP
-Gluconeogenesis is less important in plants than in animals because plants center on carbohydrates and
animals on fats for energy.
Pentose phosphate pathway is more important in plants to supply shikimate pathway in which Trp, Tyr, Phe
are formed (Phe is a precursor to Alkaloids, Flavanoids, lignin precursors).
43
Relevance to global warming!!
Global warming due to climate disruption (alteration of climate chiefly due to
burning of fossil fuels since the beginning of the industrial revolution) is predicted
(by experts) to have impacts on plant growth, development, and crop productivity.
As nights grow warmer, root respiration is predicted to increase, leading to
utilization of more photosynthate for respiration in the root at night. This is
predicted to reduce yield.
Also, regarding photorespiration, which disipates ~25% of the energy the plant
captures in photosynthesis, it may be difficult to reduce this process without
negative impacts on growth. Recent research suggests that inhibition of
photorespiration would reduce the ability of many plants to photoassimilate nitrate.
This may explain why many plants are unable to sustain rapid growth when there is
a significant increase in atmospheric CO2. So, this is another example of how
metabolic processes are interconnected. But, a paper in 2012 suggests that
transgenic manipulations introducing several enzymes may be useful to get around
this difficulty. Front Plant Sci. 2012;3:38,2012.00038
44
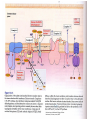
The photorespiratory carbon and nitrogen cycle (black) of a C3-plant
short-circuited by the novel glycolate catabolic pathway (red). The
transgenic enzymes introduced into A. thaliana chloroplasts are highlighted in
green. DiT1, dicarboxylate translocator 1; DiT2, dicarboxylate translocator 2;
CAT, catalase; GDC, glycine decarboxylase; GGAT: glutamate–glyoxylate
aminotransferase; GLYK, glycerate kinase; GO, glycolate oxidase; GOGAT,
glutamate–oxoglutarate aminotransferase; GS, glutamine synthetase; HPR,
hydroxypyruvate reductase; ME, NADP-malic enzyme; MS, malate synthase;
PDH, pyruvate dehydrogenase; PGP, phosphoglycolate phosphatase; SGAT,
serine–glutamate aminotransferase; SHMT, serine hydroxymethyl transferase.
Front Plant Sci. 2012;3:38. doi: 10.3389/fpls.2012.00038. Epub 2012
Feb 28.
Transgenic Introduction of a Glycolate Oxidative Cycle into A. thaliana
Chloroplasts Leads to Growth Improvement.
Maier A, Fahnenstich H, von Caemmerer S, Engqvist MK, Weber AP,
Flügge UI, Maurino VG
Abstract
The photorespiratory pathway helps illuminated C(3)-plants
under conditions of limited CO(2) availability by effectively
exporting reducing equivalents in form of glycolate out of the
chloroplast and regenerating glycerate-3-P as substrate for
RubisCO. On the other hand, this pathway is considered as
probably futile because previously assimilated CO(2) is
released in mitochondria. Consequently, a lot of effort has
been made to reduce this CO(2) loss either by reducing fluxes
via engineering RubisCO or circumventing mitochondrial
CO(2) release by the introduction of new enzyme activities.
Here we present an approach following the latter route,
introducing a complete glycolate catabolic cycle in
chloroplasts of Arabidopsis thaliana comprising glycolate
oxidase (GO), malate synthase (MS), and catalase (CAT).
Results from plants bearing both GO and MS activities have
already been reported (Fahnenstich et al., 2008). This
previous work showed that the H(2)O(2) produced by GO had
strongly negative effects. These effects can be prevented by
introducing a plastidial catalase activity, as reported here.
Transgenic lines bearing all three transgenic enzyme activities
were identified and some with higher CAT activity showed
higher dry weight, higher photosynthetic rates, and changes in
glycine/serine ratio compared to the wild type. This indicates
that the fine-tuning of transgenic enzyme activities in the
chloroplasts seems crucial and strongly suggests that the
approach is valid and that it is possible to improve the growth
of A. thaliana by introducing a synthetic glycolate oxidative
cycle into chloroplasts.
So, interventions like this one can supply extra CO2 and
reducing equivalents to RubisCO to decrease
phytorespiration and increase photosynthetic efficiency. Will
this work under higher CO2 from climate change?