Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
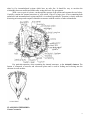
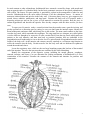
DEUTEROSTOME LABORATORY Phylum Echinodermata Class Crinoidea 1. Comatulid crinoid – observe the loss of a stalk, free-living crinoids Antedon bifida – thin, pinnate arms, 25 short cirri arising from a central disk 2. Stalked crinoid – permanently attached Class Ophiuroidea 1. Gorgoncephalus eucnemis – branching of arms repeatedly dichotomous, color varies 2. Ophioderma brevispina 3. Live ophiuroid to observe movement and behavior Class Asteroidea 1. Asterias rubens – whole preserved specimens for dissection 2. Asterias rubens – model 3. Asterias rubens arm, cross-section 4. Asterias rubens – development, bipinnaria, brachiolaria larva (completed in embryology lab) 5. Oreaster reticulates – Carribean cushion star 6. Pisaster ochraceous – Pacific sea star, arm number varies between 4-7 7. Patira miniata – bat star, very broad disk, arms practicall indistinct, 49 arms, aboral surface, crescent shaped ossicles Class Echinoidea 1. Arbacia punctulata – preserved specimens for dissection 2. Lytechinus variegatus – variegated sea urchin (Carribean), live specimens for embryology, observe pluteus 3. Strongylocentrotus droebachiensis – green sea urchin, test only 4. Encope emarginata – bowed petals, test discoidal with flat base 5. Melitta quinquiesperforata – key hole urchin, tropical 6. Spatangoid sea urchin heart urchin, observe ambulacra, peristome and periproct, lantern absent Class Holothuroidea 1. Cucumaria – preserved specimens for dissection 2. Cucumaria – preserved specimens, observe external structure, particularly tentacles Phylum Urchordata 1. preserved adult specimens to examine adult features 2. tadpole larval slides to find deuterostome characteristics Phylum Cephalochordata 1. prepared slide 1 PHYLUM ECHINODERMATA The phylum Echinodermata is clearly separated from all other phyla. Its members are coelomate, and the coelom is of great importance not only in carrying and distributing nutrients, respiratory gases, excretory and storage products but also in functioning as a water-vascular system. The water-vascular system operates the hydraulic, extensible tube-feet which are diagnostic of the phylum and which play an essential role in the life processes of these animals and their interaction with the environment. Echinoderms comprise a large phylum of specialized marine organisms that are interesting for both their striking body form, their deuterostome characteristics and phylogenetic position as precursors to the vertebrate line. Though adults of most echinoderms show radial symmetry, this does not ally them with the Cnidaria. The radial symmetry of echinoderms is a secondary characteristic associated with their sessile or creeping lifestyle. The larval stages of echinoderms are actively swimming bilaterally symmetrical animals. Most echinoderms have a calcite skeleton. In some, such as ophiuroids (brittle stars), the skeleton is in the form of rods supporting arms and other projections; in others, such as asteroids (starfish) and echinoids (sea urchins), it forms a more-or-less rigid capsule; finally, in others, such as holothuroids (sea cucumbers), it is highly reduced and represented only by isolated spicules in the integument. The skeleton is generally not solid, but is built as a crystalline calcite meshwork, with living protein material permeating the crystalline framework. The better-known members, such as sea urchins, brittlestars, and starfish, have skeletal spines to which the name Echinodermata (spiny skinned) refers. But possession of spines is not diagnostic of the phylum. The earliest echinoderms were thought to be cup-shaped animals with a five-part symmetry (pentamerous), and a sessile lifestyle with the mouth uppermost and the arms held out radially to catch food. The crinoids (sea lilies) are the only extant group to retain this primitive form. From this form, two different patterns evolved: one (Asterozoa) showed radial growth patterns as evident in starfish and brittle stars, while the other (Echinozoa) showed fusion of the original radial appendage plan as seen in urchins and sea cucumbers. While the Asterozoa universally retain radial symmetry, there is a tendency within the Echinozoa to assume a tertiary bilateral symmetry. This is reflected in the worm-like body of holothuroids (sea cucumbers) and in the heart shape of some irregular echinoids (urchins). Asterozoa and Echinozoa can be considered subphyla of Echinodermata. I. SUBPHYLUM CRINOZOA Mouth uppermost, arms well-developed and often branching; sealilies and feather stars. Crinoids are the most primitive echinoderms. Early fossil crinoids had stems, by which they were attached permanently to the substrate, but subsequently the stem was lost so that the animal could swim or crawl. The mouth is always directed upward and feeding is accomplished by filtering with the arms. The arms are also used for swimming. You have some specimens i.e. Antedon sp. free-living comatulids in lab for observation. II. SUBPHYLUM ASTEROZOA = Star-shaped; mouth on undersurface. A. CLASS OPHIUROIDEA Arms cylindrical and sharply demarcated from the central disc; brittle stars. Based on the coelom and water vascular system, brittlestars are thought to be the group next most primitive to the crinoids. They are also highly successful and probably outnumber all other classes of echinoderms in number of species. B. CLASS ASTEROIDEA Arms not sharply demarcated from the central disc, well-developed water vascular system with suction tube feet; starfish. Whereas the ophiuroids use their arms for propulsion over the sea floor, asteroids rely on movements of the tube-feet for locomotion, and while ophiuroid skeletons are fragile, asteroid skeletons are strong and capable of pulling apart bivalve shells. 2 III. SUBPHYLUM ECHINOZOA = Globose or worm-like, without radiating arms; some tertiary bilateral symmetry. A. CLASS ECHINOIDEA Skeleton consists of heavy plates that enclose the body in a rigid test. Sea urchins and sand dollars. B. CLASS HOLOTHUROIDEA Bilaterally symmetrical with an elongated oral-anal axis; skeleton reduced to ossicles in the integument; filter and/or deposit feeding with modified oral tube-feet called buccal tentacles. CLASS OPHIUROIDEA Genus Ophioderma Obtain an Ophioderma specimen and place it in a fingerbowl of seawater for examination under the dissecting microscope. Brittle stars are superficially similar to asteroids, but their long sinuous arms are more distinctly set off from the central disc and they locomote and feed in an entirely different manner. The mouth is located in the center of the oral surface of the central disc and is surrounded by five triangular jaws. The arms have reduced tube feet called tentacles that issue from between the skeletal plates or shields of the oral surface of the arms. There is no ambulacral groove and the skeletal plates are arranged so that the arms are flexible. On the edge of the central disc are located ten pairs of genital bursae that store gametes. Note the rough nature of the spines that are used for gripping the substrate, and the absence of suctorial discs on the tube feet. How do brittle stars move? Examine the aboral surface of Ophioderma. Can you locate a madreporite? pedicellariae? dermal papulae? The skeletal plates of the ophiuroid skeleton form a complete, rigid covering over the central disc. The skeleton of the arms consists of two parts: an outer skeleton composed of aboral, oral, and lateral shields, and an internal series of vertebral ossicles. This arrangement is designed to allow the arms to move freely and enables the brittlestar to crawl rapidly and swim. To examine the skeletal structure of Ophioderma, remove the distal part of an arm and place it in sodium hypochlorite to clean. Examine and describe the skeletal plates. How are they designed to allow movement? Try to observe both crawling and swimming movements. Some brittlestars have feathery spines that allow them to swim quite effectively. Feeding: Ophiuroids are microphagous ciliary-mucoid feeders and may feed macrophagously as well. Mucous glands in the integument produce a network of mucus on the surface of the arms, and the mucus and food entrapped by it are moved by cilia, tube feet, and arms to the mouth. They also sweep their arms over the substrate to capture larger food particles, which may be brought to the mouth by flexion of the arms or by passage from podia to podia. Place your Ophioderma in a fingerbowl and offer it pieces of chopped mussel in the following sizes to detect different feeding modes: 1) 10 x 5 mm; 2) <2 mm squares, and 3) finely-ground mussel. In hopes of observing a mucous net, introduce into the dish of seawater a weak solution of toluidine blue so that the water is colored a faint blue. CLASS ASTEROIDEA Genus Asterias Obtain a small specimen of Asterias rubens and place it under the dissecting microscope for external observation. Carefully examine the oral and aboral surfaces and identify the structures. The mouth is located on the oral surface of the central disc, and radiating outward from the disc are five arms or rays. Running down the center of each ray is the ambulacral groove with four rows of tube feet. The roof of the ambulacral groove contains a radial nerve, which you may be able to distinguish from the other tissue. Closely examine the structure of the tube feet which originate lateral to the radial nerve. Try to watch a tube foot attach to a probe. Modified spines border each ambulacral groove and the mouth. The 3 ambulacral spines serve to protect the tube feet, while the oral spines help move food into the mouth. Locate the tentacle at the end of each ray. These tentacles are modified tube feet and have a sensory function. At the base of each tentacle is located an eyespot which is light sensitive. On the aboral side of your Asterias locate the conspicuous dome-shaped madreporite (sieve plate) which opens into the stone canal of the water-vascular system. The function of this perforated plate is uncertain. It appears to function either as a device for ensuring equal pressure within the water-vascular system and the external environment, or as a pressure receptor for providing information to the tube feet about the changing head of water above the animal as the tide rises and falls. A small anus is located in the inter radius clockwise from the madreporite. The rays on either side of the madreporite are termed bivium arms and the remaining three arms are called trivium arms. The aboral surface is covered by cilia and calcareous tubercles, spines, and beak-like pedicellariae. To examine the skeleton and ossicles of Asterias, remove a small piece of skin and boil it in sodium hypochlorite. Examine the ossicles under the compound microscope and sketch them so you can compare them with ossicles from the other echinoderm classes. The intact skeleton of Asterias consists of articulating ossicles (plates) of calcite (calcium carbonate) held together by leathery connective tissue. Be sure you closely examine the surface skeletal structure of Asterias. Carefully cut into the aboral surface tracing the central disk outline. Trace out the path of the water vascular system. The madreporite opens into the stone canal which leads to the ring canal around the esophagus. Tease around the digestive tract to see these structures. On the edge of the radial canal are nine swollen areas referred to as Tiedemann's bodies. Their function is uncertain, but they are thought to filter the water vascular system of debris with the aid of the amoebocytes which are found concentrated there. They are then much like the lymph nodes of vertebrates. Leading from the ring canal are five radial vessels each running the length of a single ray. Along the radial vessels are located the tube feet and ampullae. How do the tube feet work? What is the function of the ampullae? Examine a prepared slide of a cross section of a starfish ray to get a better look at the functional organization of the water vascular system. Now locate the structures of the digestive system. A short esophagus leads from the mouth and connects to the stomach. The stomach is divided into a globular protruding cardiac portion and an aboral pyloric region. Running from the pyloric portion into each arm are a pair of hepatic caeca that function in digestion and food storage. The rectum runs to the aboral surface from the pyloric stomach and exits via the aboral anus. Cardiac retractor muscles control the protrusion of the cardiac stomach. 4 The testes or ovaries of Asterias are paired organs located in each ray. Asteroids are dioecious. Each gonad is attached by a gonoduct to the exterior via a gonopore located at the junction of the rays. Determine the sex of your animal by making a gonadal smear. CLASS ECHINOIDEA Genus Strongylocentrotus Obtain a specimen and examine the oral surface and locate the mouth which bears five protruding teeth used for scraping and chewing algae. A soft, membranous area called the peristome surrounds the mouth and has modified buccal tube feet which have a chemosensory function. At the edge of the peristome are five pairs of gills that are invaginations of the body wall that communicate with the coelom and are thought to have a respiratory function. On the aboral surface locate the periproct, madreporite and genital plates. The genital plates are perforated and provide exit for the gametes. Between each genital plate is an ocular plate which has a single pore through which a modified light-sensitive tube foot/tentacle emerge. Examine the surface of your urchin under the dissecting scope. Note that the entire external surface is ciliated. Examine the spines of Arbacia. Are they all similar? Why is this so? Examine how the spines are attached to the surface of the test (skeleton. Locate the tube feet on the urchin and note that they are restricted to five bands on the test known as the ambulacra. The ambulacra areas are separated from each 5 other by five interambulacral regions which have no tube feet. It should be easy to envision the relationship between starfish and urchin after seeing this basic five-part pattern. With a saw or pair of scissors, cut through the equator of the urchin and separate the two halves. Carefully examine the internal structures of your urchin. Chip off a large piece of test containing both ambulacral and interambulacral regions and boil it in sodium hypochlorite. Examine it closely under the dissecting microscope and compare echinoid test structure with the ossicles of other echinoderms. Pay particular attention, when examining the internal structures, to the Aristotle's lantern. The lantern is composed of muscles and calcareous plates and is used in feeding and in boring into the substrate in some urchins CLASS HOLOTHUROIDEA Genus Cucumaria 6 In stark contrast to other echinoderms, holothuroids have assumed a worm-like shape, with mouth and anus at opposite ends of a cylindrical body; but the basic pentameric structure of the typical echinoderm is still evident. The former five-part symmetry is particularly evident in the arrangement of tube feet and muscles. Holothuroids have the most reduced skeleton of all the echinoderm classes. It consists of numerous isolated plates and spicules in the body wall. Calcareous plates, however, give support to the mouth, cloaca, tentacles, madreporite, and ring canal. Examine the body wall of Cucumaria under a dissecting microscope and cut out a piece of wall material to examine the spicules. Boil the piece in sodium hypochlorite and describe the spicules. How do they compare with the other ossicles you have seen. To open your Cucumaria make a ventral incision from the mouth to anus, open the incision, and pin the specimen to a dissecting pan. In cutting, be sure you don't harm the internal organs. Identify the dorsal madreporite and stone canal, which hang free in the coelom. The stone canal connects to the water vascular ring canal, which surrounds the esophagus. The ring canal has two elongate sacs called polian vesicles which are thought to be reservoirs of water vascular fluid. From the ring canal, radial canals run anterior to the oral tentacles, and then turn back as posterior branches that are embedded in the longitudinal muscle bands and give rise to the tube feet. Five paired bands of longitudinal muscles run the length of the body wall. At the anterior end retractor muscles, attached to a calcareous ring function to retract the tentacles into the body. Circular muscles line the body wall and are thickened as sphincters around the mouth and cloaca. Locate the respiratory trees, which are the two large branching organs that lead out of the terminal end of the intestine. These are the structures that are ventilated by the cloacal breathing. Identify the components of the digestive system, including the mouth, pharynx, esophagus, stomach, intestine, and cloaca. Can you determine the feeding mechanism of holothuroids by a close examination of the tentacles? Examine the contents of the stomach and intestine for parasites and commensals. 7 Phylum Urochordata Tunicates are highly derived and less like the ancestral chordates than are cephalochordates or vertebrates. At some time in the life cycle all possess a notochord, dorsal hollow nerve cord, pharyngeal gill slits, postanal tail, and endostyle but only the gill slits and endostyle are present in adults. Tunicates use the pharyngeal gill slits for suspension feeding. The larva is much more chordate-like than the adult and resembles a tadpole or fish, has all the chordate apomorphies, and is known as the tadpole larva. Metanephridia are absent and coelom is reduced to a pericardial cavity and gonads. As in cephalochordates the gut is dominated by an enormous pharynx surrounded by a water-filled atrium but unlike cephalochordates, it is U-shaped with the mouth and anus anterior. Tunicates may be benthic or planktonic and solitary or colonial. All are marine. Tunicata is traditionally divided into Ascidiacea (the benthic sea squirts in three taxa; Aplousobranchia, Phlebobranchia, and Stolidobranchia), Thaliacea (the pelagic salps), and Appendicularia (the pelagic larvaceans). Recent molecular evidence and reevaluation of morphological evidence, however, suggests that Ascidiacea is paraphyletic and Tunicata should be reorganized into three different higher taxa. In this reorganization Stolidobranchia would be one higher taxon. Phlebobranchia plus Thaliacea would be the second taxon. Aplousobranchia plus Appendicularia is the final tunicate taxon. For now, however, the traditional classification will be followed. Ascidiacea = Ascidiacea is usually taken as representative of Tunicata, at least for the purposes of introductory laboratory exercises. Ascidians, or sea squirts, are sessile filter feeders that, as adults, bear little resemblance to their chordate relatives. Ascidians have a living, external, cellular exoskeleton, or tunic, underlain by epidermis. The tunic resembles connective tissue, except it is outside the epidermis, and consists of cells, a secreted extracellular matrix, and ground substance. Much of it is a cellulose-like polysaccharide. In many ascidians blood vessels cross the epidermis to enter the tunic, a feature found in no other animal. The gut is U-shaped and both openings are anterior, with the anus dorsal to the mouth. The gut is dominated by an enormous pharynx whose wall is perforated by numerous tiny gill slits. The pharynx is surrounded by a water-filled atrium into which the gill slits open and which itself opens to the sea. It is both respiratory organ and filter-feeding device. Water and food particles enter the pharynx and the water passes through the gill slits to the atrium and then out the siphon. Food, entangled in mucus secreted by the endostyle, remains in the gut and passes posteriorly to be digested. The hemal system includes a heart, vessels, and blood spaces in the connective tissue. The heart is enclosed in a pericardial cavity derived from the ancestral coelom. The pattern of blood flow resembles that of the cephalochordates and early vertebrates except that the heart reverses direction periodically and the blood thus flows in both directions through the system. Ascidians have no structure recognizable as a kidney. Ascidians are simultaneous hermaphrodites and the gonoducts open into the atrium. Some ascidians are solitary and may be relatively large. Others are colonial with tiny individual zooids in a common tunic. Pleurogona = The gonads are on the inner surface of the body wall. The epicardium is absent or represented by a renal sac. Stolidobranchia = Stolidobranchia are the higher ascidian taxa. In stolidobranchs the gonads are on the inside surface of the body wall beside the pharynx and not in the gut loop. No epicardium is present but Molgulidae has renal sacs, which are derived from the epicardium. The neural gland is dorsal to the cerebral ganglion. The stolidobranch pharyngeal lining is strongly pleated and has transverse and longitudinal blood vessels. Molgulidae has spiral gill slits. Stolidobranchs can be solitary or colonial. The plane of the tadpole tail is vertical. Molgula Examine preserved specimens of Molgula to find the buccal and atrial siphons, pharyngeal gills slits, intestine and general location of gonads. Describe the water flow within the organism and determine feeding! 8 Examine a commercially prepared whole mount of a tadpole larva of unknown species. Very little internal structure is discernable in most commercial slides but you can see the general shape of the larva and a few structures. It consists of an ovoid anterior body with a long, slender, muscular, posterior tail. In some preparations a thick, transparent, almost invisible tunicsurrounds the body. The notochord is usually visible in the center of the tail. Within the body the cerebral vesicle (= sensory vesicle) should be apparent. It contains at least one (as in Molgula), but more often two, opaque spheres. One is a statocyst for gravity detection and the other is an eyespot for photoreception. The cerebral vesicle will become the cerebral ganglion of the adult. Two adhesive papillae are present at the extreme anterior end. These will be used to attach to the substratum when the larva leaves the plankton and begins its sessile adult life. The gut, with nonfunctional pharynx, endostyle, and intestine are usually an amorphous mass whose details cannot be recognized in these slides. Buccal and atrial siphons, although present, are neither. Phylum Cephalochordata Cephalochordates retain many of the features of the ancestral chordate including the dorsal hollow nerve cord, notochord, postanal tail, and pharyngeal gill slits used for filter feeding. The swimming and feeding modes are like those hypothesized for the ancestor. Introduction As the most vertebrate-like of invertebrates, cephalochordates are studied in either vertebrate or invertebrate zoology courses. Comparative vertebrate anatomy traditionally begins with a consideration of cephalochordates and other protochordates and invertebrate zoology often ends with them. Protochordates are the inver-tebrate members of Chordata and include urochordates (sea squirts), and cephalochordates (amphioxus, lancelets). These taxa are clearly related to vertebrates but also show unmistakable 9 relationships with other invertebrates, most notably echinoderms and hemichordates. These then are the taxa that establish the evolutionary connections between the vertebrates and rest of the deuterostomes and ultimately, the remainder of the animal kingdom. Amphioxus With about twenty-five species inhabiting shallow tropical and temperate oceans, the Cephalochordata are a very small branch of the animal kingdom. Known as lancelets or as amphioxus (from the Greek for "both [ends] pointed," in reference to their shape), cephalochordates are small, eel-like, unprepossessing animals that spend much of their time buried in sand. However, because of their remarkable morphology, they have proved crucial in understanding the morphology and evolution of chordates in general -including vertebrates. The anatomy of a cephalochordate is diagrammed at left. Note that cephalochordates have all the typical chordate features. The dorsal nerve cord is supported by a muscularized rod, or notochord. The pharynx is perforated by over 100 pharyngeal slits or "gill slits", which are used to strain food particles out of the water. The musculature of the body is divided up into V-shaped blocks, or myomeres, and there is a postanal tail. All of these features are shared with vertebrates. On the other hand, cephalochordates lack features found in most or all true vertebrates: the brain is very small and poorly developed, sense organs are also poorly developed, and there are no true vertebrae. Water is taken in through the mouth, drawn in by the beating of cilia located on the wheel organ, a set of ridges lying inside the mouth. The water is first filtered by the oral cirri, slender projections that surround the opening of the mouth, clearly visible on the photograph at the top of the page. It then passes through the gill slits. These gill slits are enclosed by folds of the body wall, the metapleural folds, to form a body cavity known as the atrium. Food particles in the water are trapped by mucus, while water passes through the slits and out of the atrium through the atriopore, located towards the posterior end. The rest of the digestive system is fairly simple: a pouch or hepatic caecum secretes digestive enzymes, and actual digestion takes place in a specialized part of the intestine known as the iliocolonic ring. Cephalochordates also have a well-developed circulatory system and a simple excretory system composed of paired nephridia. The sexes are separate, and both males and females have multiple paired gonads. Eggs are fertilized externally, and develop into free-swimming, fishlike larvae. 10 11