Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
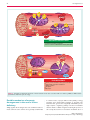
REVIEW European Journal of Heart Failure (2016) doi:10.1002/ejhf.467 The influence of iron deficiency on the functioning of skeletal muscles: experimental evidence and clinical implications Magdalena Stugiewicz1, Michał Tkaczyszyn2,3, Monika Kasztura2, Waldemar Banasiak3, Piotr Ponikowski3,4, and Ewa A. Jankowska2,3,* 1 Students’ Scientific Association, Laboratory for Applied Research on Cardiovascular System, Department of Heart Diseases, Wroclaw Medical University, Wroclaw, Poland; for Applied Research on Cardiovascular System, Department of Heart Diseases, Wroclaw Medical University, Wroclaw, Poland; 3 Cardiology Department, Centre for Heart Diseases, Military Hospital, Wroclaw, Poland; and 4 Department of Heart Diseases, Wroclaw Medical University, Wroclaw, Poland 2 Laboratory Received 30 June 2015; revised 16 October 2015; accepted 22 October 2015 Skeletal and respiratory myopathy not only constitutes an important pathophysiological feature of heart failure and chronic obstructive pulmonary disease, but also contributes to debilitating symptomatology and predicts worse outcomes in these patients. Accumulated evidence from laboratory experiments, animal models, and interventional studies in sports medicine suggests that undisturbed systemic iron homeostasis significantly contributes to the effective functioning of skeletal muscles. In this review, we discuss the role of iron status for the functioning of skeletal muscle tissue, and highlight iron deficiency as an emerging therapeutic target in chronic diseases accompanied by a marked muscle dysfunction. .......................................................................................................... Iron deficiency • Skeletal muscles • Exercise intolerance Heart failure • Introduction Skeletal and respiratory muscle dysfunction constitutes an important pathophysiological feature of heart failure (HF) and chronic obstructive pulmonary disease (COPD), highly prevalent chronic diseases, which account for a significant health and socio-economic burden in developed countries.1 – 6 As skeletal and respiratory myopathy worsens symptoms and outcomes in patients with HF and COPD,3 – 6 it is plausible to consider muscle tissue abnormalities as a co-target in the therapeutic process. Functional abnormalities of skeletal and respiratory muscle tissue in HF and COPD correlate with structural derangements, which occur at both histological and molecular levels, and have all been described.7 – 10 Since the vast majority of abnormalities impair muscle energetics, it is presumed that there may be a common denominator for those derangements. ................................................ Keywords Chronic obstructive pulmonary disease • There is evidence that iron plays a critical role in the optimal functioning of skeletal muscle tissue. It is an essential micronutrient for oxidative energy metabolism as well as numerous cellular processes.11 The particular importance of optimal iron status is well reflected by the fact that both iron deficiency (ID) and iron overload are detrimental to the cellular machinery involved in energy generation.12 Iron overload represents one arm of the U-shaped curve describing the relationship between cellular vitality, energy generation capacity, and iron status, and its potential for excessive generation of reactive oxygen species (ROS) has already been discussed.13,14 In this review, we aimed to summarize the evidence on the relationship between ID and skeletal muscle dysfunction, from the subcellular to macroscopic levels. Further, we discuss the possible pathophysiological links between ID and myopathy in HF and COPD, along with potential clinical implications. *Corresponding author. Laboratory for Applied Research on Cardiovascular System, Department of Heart Diseases, Wroclaw Medical University, Centre for Heart Diseases, Military Hospital, ul. Weigla 5, 50-981 Wroclaw, Poland. Tel:/Fax: +48 261 660 661, Email: [email protected] © 2016 The Authors European Journal of Heart Failure © 2016 European Society of Cardiology 2 Skeletal muscle plasticity for ATP utilization The efficient functioning of skeletal muscle is directly related to its intact energy metabolism because of the extraordinarily high energy demand in comparison with other body tissues, such as smooth muscle, liver, or even brain.15 Therefore, skeletal muscle must be highly specialized in generating ATP as an immediate source of energy and must also provide adequate flexibility to meet the challenge of dynamic changes of ATP utilization. Indeed, the substantial increase (up to 300-fold) in energy turnover from the resting to the fully activated state occurs in a matter of milliseconds.16 In contrast, muscles have to deal with long periods of moderately increased energy consumption. These large fluctuations in energy demand require the complex network of interacting pathways of fuel metabolism which result in ATP production.17 – 19 Fibre types and their energetics The quintessential attribute of skeletal muscles that significantly contributes to their multitasking is the co-existence of different fibre types which allows the same muscle to be used for various tasks from posture maintenance to explosive movements in response to an unexpected threat. The presently dominating classification system for human skeletal muscle is based on the myosin heavy chain (MHC) isoforms and determines three major fibre types: I, IIa, and IIx.6,19 – 21 The slow type I fibres have highly oxidative properties, such as high mitochondrial content and high activity of iron-dependent enzymes, and thus they are adapted to long-lasting repetitive activities such as locomotion or respiration.6,20 – 24 The fast glycolytic type IIx fibres exhibit a high glycolytic capacity (generate ATP mostly via glycolysis), and thus are required for rapid and generally powerful actions.6,20 – 24 The fast oxidative type IIa fibres take advantage of both oxidative metabolism and the glycolytic pathway, and therefore their susceptibility to fatigue is intermediate.6,20 – 24 Energy store in skeletal muscle The immediate energy source during muscle contraction is a phosphate–phosphate bond present in ATP.16 Skeletal muscle demonstrates a relatively poor reserve (∼80 g) of this energy currency, which is sufficient only for 1–2 s of mechanical effort and must be continuously re-synthesized at the same rate as its consumption.25,26 Hence, there is a need for efficient ATP restoration, which occurs via three main mechanisms: short-term phosphocreatine (PCr) hydrolysis, medium-term anaerobic glycolysis, and long-term aerobic oxidative phosphorylation.16 ........................................................................................................................................................................ Skeletal muscle: highly specialized tissue with an extraordinarily high energy demand M. Stugiewicz et al. Energy metabolism during strenuous activity Upon the stimulus of exercise, the initial pathway of ATP restoration relies on the hydrolysis of a high-phosphate substrate, a reaction catalyzed by creatine kinase (CK) which provides 16% of energy currency generated via anaerobic pathways.27 In turn, PCr re-synthesis carried out by the mitochondrial isoform of CK occurs when ATP is in abundance under resting conditions.16,28 The fuel for an increased turnover of ATP during strenuous exercise is provided by carbohydrate catabolism, namely by the conversion of intramuscular glycogen and blood glucose.29 After an initial consumption of the energetically more efficient glycogen store,30 the contribution of blood glucose becomes more appreciable and constitutes up to 35% of oxidative metabolism and nearly 100% of muscle carbohydrate metabolism.29 Energy metabolism during long-term activity Carbohydrate substrates can be metabolized via two pathways, namely anaerobic glycolysis and aerobic oxidative phosphorylation.16 The first process occurs in the cytoplasm and permits the high performance of skeletal muscle when oxidative metabolism alone is not sufficient. As soon as the oxygen supply is plentiful, the catabolism of the same substrates results in more efficient ATP synthesis which is then performed within the mitochondrial respiratory chain in the oxidative pathway.16 Besides carbohydrate substrates, the second main fuel for muscle oxidative metabolism is fat.31 Being released from their stores in adipose tissue and muscle, and metabolized in mitochondria, fatty acids are the preferred fuel used for a sustained contractile function in oxidative skeletal muscle.32 Critical role of optimal iron availability for effective cellular energy metabolism in skeletal muscle Systemic vs. local iron metabolism in skeletal muscle Recently, there has been great interest in skeletal muscle in the context of iron metabolism, as this tissue contains 10–15% of iron in the body.33,34 Iron is fundamental to oxidative metabolism in skeletal muscle, both for efficient oxygen storage in myoglobin and for an optimal activity of mitochondrial enzymes. Therefore, iron is present in a larger amount in slow, ‘red’ fibres, which are common in, for example, dorsal muscles, lower extremity extensors, the diaphragm, and intercostal muscles.35 In general, one should differentiate systemic iron metabolism from local iron metabolism. On the whole, systemic iron homeostasis is maintained by the co-ordination of its absorption in the © 2016 The Authors European Journal of Heart Failure © 2016 European Society of Cardiology 3 duodenum, recycling of senescent erythrocytes, and a mobilization of iron stored in the liver. The role of skeletal muscle as a tissue utilizing and storing iron, and possibly in secreting molecules modulating systemic iron metabolism (haemojuvelin; HJV) may be hypothesized (see below),36,37 but has not been proven yet (Figure 1). All major processes within systemic iron metabolism are controlled by hepatic hepcidin (HAMP), the key iron regulatory hormone.38,39 HAMP down-regulates a membrane iron exporter, ferroportin, thus inhibiting iron efflux from iron-storing and iron-distributing cells into the circulation, where iron travels bound to transferrin (Tf). Hepatic HAMP expression is regulated by: (i) body iron load [through bone morphogenetic proteins (BMPs) together with HJV);39,40 (ii) inflammation, through interleukin-6 (IL-6) and IL-22;39 and (iii) erythropoietic activity.39 Although HAMP is produced predominantly by hepatocytes, it can be detected in other tissues, including skeletal muscle, where it participates in local iron regulation, which is different from systemic regulation and not clearly understood. Molecules involved in skeletal muscle iron metabolism and their potential participation in systemic iron regulation There is a substantial gap in the understanding of muscle-specific iron regulation. Only recently has local iron metabolism in skeletal muscle started to be explored. For example, Polonifi et al. examined skeletal muscle iron metabolism and confirmed the expression of several genes implicated in four pathways: iron import, export, storage, and regulation (for a detailed review of iron metabolism genes in skeletal muscle, see Polonifi et al.41 and Sekyere et al.42 ) As mentioned before, little has as yet been unravelled regarding the mechanisms that control local iron regulation in skeletal muscle. Since the expression of two main regulatory peptides, namely HAMP and HJV, has been confirmed in skeletal muscle,37,41 the existence of tissue-specific translational iron regulation can be assumed. Although the production of HAMP in skeletal muscle is negligible (in comparison with hepatic production), some preliminary results indicate its potential contribution to local iron regulation and immune response.43 HJV, on the other hand, is greatly expressed in skeletal muscle. Although a comprehensive study on its biogenesis has been performed, the role of muscle HJV in skeletal muscle remains undefined.36 Since muscle-derived HJV accumulates in extracellular fluid, it was proposed to influence hepatic hepcidin expression,37 but there is a need for in-depth research to support this hypothesis. Involvement of iron in cellular energetics in skeletal muscle Iron in skeletal muscle is of a particular importance for oxygen reducing systems in order to provide efficient ATP production. Oxidative energy generation takes place in mitochondria via the mitochondrial respiratory chain composed of four complex transmembrane iron-containing enzymes [haem proteins ........................................................................................................................................................................ Iron deficiency and skeletal muscles © 2016 The Authors European Journal of Heart Failure © 2016 European Society of Cardiology (cytochromes) and iron–sulfur cluster (ISC) proteins].12,28 Haem is a part of the cytochromes in complexes III and IV, whereas ISCs are incorporated into respiratory complexes I, II, and III.12,28 Mechanisms aiming to preserve normal intracellular iron status Since iron-deficient conditions result in impairment of many cellular processes and iron-containing enzymes, a single cell should be in possession of mechanisms that minimize detrimental changes. Mammalian cells employ iron-regulatory proteins 1/2 (IRP1/2) that constitute central regulators of adaptive intracellular response to ID (for a detailed review of IRP1/2, see Andeerson et al.44 ). Importantly, the expression of IRP in rat skeletal muscle has been reported by Guo et al., and the levels of IRP2 in myocardial and skeletal muscle tissues were reported to be the highest of all examined tissues.45 Recent studies have revealed the existence of another iron-regulatory pathway in mammalian cells, which involves the protein called tristetraprolin (TTP). TTP is induced by ID within a cell, and its presence is critical for cell survival in low-iron states,13 presumably by suppressing iron-consuming cellular processes, such as haem synthesis, the Krebs cycle (KC), or the mitochondrial electron transport chain (ETC). The limitation of the aforementioned processes may not be beneficial in the context of energy generation.46 Although the expression of TTP in skeletal muscle has been neither investigated nor proven to date, this pathway could potentially contribute to deranged skeletal muscle cell energetics in the course of ID. Molecular effects of depleted iron on skeletal muscle energetics: evidence from in vitro and animal studies Multifaceted detrimental effects of ID for the organism in the context of skeletal muscles involve decreased productivity and exercise capacity, and several alterations within the muscle tissue (Figure 2). Experimental data from in vitro and animal studies reveal that skeletal muscle energetics suffer from iron depletion at different levels, from shifts in energy substrate preferences to subcellular and molecular derangements, which result mostly in a decrement of oxidative capacity (Figure 3).47 – 50 Changes in fuel selection Several studies have demonstrated that the intracellular iron status influences the fuel selection for muscle energy metabolism. For example, the activity of a lactate dehydrogenase isoenzyme in muscle of iron-deficient rats was increased as compared with healthy controls, which indicated a clear shift towards anaerobic metabolism upon ID in order to maximize muscle capacity.50 – 54 It is hypothesized that these biochemical changes may also result 4 M. Stugiewicz et al. Liver Skeletal muscle HEPATIC HEPCIDIN MUSCLE HEMOJUVELIN +? FPN – +/– ? – Fe3+ –Tf FPN Fe FPN Macrophage MUSCLE HEPCIDIN - possible regulator of local iron metabolism FPN Duodeum Myocyte in alterations in skeletal muscle fibre composition, namely an oxidative to glycolytic shift, which has been demonstrated in different muscle pathologies associated with diminished oxidative capacity.55 Derangements in mitochondrial morphology Intracellular iron depletion also affects the morphology of mitochondria. Cartier et al. provided electron micrographs of muscle from iron-deficient rats, demonstrating an apparent decrease in the density of cristae of the mitochondrial inner membrane.48 Since these structures contain specific binding sites for mitochondrial enzymes involved in oxidative energy metabolism (i.e. complexes III and IV) and in ISC biogenesis, such a change should not persist without an impact on the organelle efficacy.56 Impaired oxidative metabolism Iron defiency severely affects the performance of respiratory chain enzymes. The activities of mitochondrial complexes I, II, and IV, and the activity of the machinery responsible for ISC .................................................................... Figure 1 The contribution of skeletal muscles to systemic and local iron regulation. Fe, iron; FPN, ferroportin; Tf, transferrin. protein maturation as well as the concentration of mitochondrial cytochromes were reported to be dramatically decreased in skeletal muscle of non-anaemic iron-deficient rats.47,48,50,51,57 – 60 Importantly, ID is also accompanied by a diminished pool of myoglobin, an oxygen-binding protein found in muscle tissue.61 The concentration of the aforementioned globin was decreased in predominantly slow- and mixed-fibre skeletal muscle from iron-deficient rats.58 Activation of a key regulator of cellular energy homeostasis (5’-AMP-activated protein kinase) Since ID constitutes an energy challenge for the cell, it induces an activation of a major sensor of cellular energetic insults, a 5’-AMP-activated protein kinase (AMPK).59,62 Indeed, the aforementioned kinase was proven to be chronically activated in iron-deficient rats.59 Since the chronic activation of AMPK is known to increase the expression of a glycolytic enzyme, i.e. hexokinase II,63,64 the described mechanism is suggested to contribute to an oxidative to glycolytic shift which occurs in iron-deficient skeletal muscle tissue. © 2016 The Authors European Journal of Heart Failure © 2016 European Society of Cardiology 5 Iron deficiency and skeletal muscles Populational effects Organism Economic productivity Overall physical work capacity Aerobic capacity Endurance capacity Aerobic & endurance adaptation after training Tissue level Cellular level − Altered muscle fiber composition (oxidative glycolytic shift) Muscle mass − Deranged mitochondrial morphology Number of mitochondria Myoglobin pool − AMPK activation Glycolytic activity Mitochondria Oxidative metabolism Iron-sulfur clusters synthesis Density of mitochondrial cristae Figure 2 Detrimental effects of iron deficiency on economic productivity, exercise capacity, and the functioning of skeletal muscle tissue. AMPK, 5’-AMP-activated protein kinase. © 2016 The Authors European Journal of Heart Failure © 2016 European Society of Cardiology 6 M. Stugiewicz et al. OXPHOS E E E OXPHOS Myoglobin KC XPHOS XP HOS OXPHOS E E OXPHOS Pyruvate Pyruva Pyr uvate uva te E KC KC Glycolysis Glu Glucos cose e E Glucose Myoglobin SUFFICI SUFFICIENT IRON STORES - Undisturbed oxidative metabolism - Efficient oxidative PHOSPHORYLATION (OXPHOS) AMPK Myoglobin E E Pyruvate Pyruva Pyr uvate uva te E E KC Glycolysis Glucose Glucos Glu cose e AMPK A E AMPK IRON DEFICIENCY Density of mitochondrial cristae (inner membrane) - Shift towards anaerobic metabolism - AMPK activation Myoglobin Glycolysis Possible mechanism of myocyte derangements in the course of iron deficiency Taking together all the changes that occur at different levels in a muscle cell in the course of ID it can be generally concluded that ................ Figure 3 Structural and functional alterations of human striated muscle tissue associated with iron deficiency. AMPK, 5’-AMP-activated protein kinase; KC, Krebs cycle; E, energy. in a low-iron state, a myocyte shifts its main pathway of energy generation from mitochondrial respiration to glycolysis. The mechanism that governs this energetically unfavourable process might comprise a regulatory pathway of local iron metabolism which is critical to cellular energetics. Presuming the presence of the recently discovered iron conservation mechanism also in a © 2016 The Authors European Journal of Heart Failure © 2016 European Society of Cardiology 7 muscle cell (see above), we might hypothesize that ID forces the myocyte to limit processes with high iron expenditure, including mitochondrial respiration. Links between iron deficiency, decreased exercise capacity, and impaired skeletal muscle performance: data from experimental animal models and human studies Decreased exercise performance in iron-deficient animals with and without concomitant anaemia Tissue oxidative capacity of skeletal muscles depends on an efficient activity of the mitochondrial enzymatic machinery involved in oxidative phosphorylation, and determines predominantly the endurance capacity (the ability to perform prolonged submaximal exercises).51,65,66 Maximal physical efforts and aerobic exercise depend on oxygen-carrying capacity, which reflects adequate oxygen supply provided by haemoglobin.51,65,66 There is evidence from experimental models that animals with iron deficiency anaemia (IDA) demonstrate decreased aerobic capacity.50,65 – 69 In turn, endurance capacity is reduced across all successive stages of progressive ID, either with or without concomitant anaemia.50,65 – 68 More than four decades ago, Edgerton et al.68 demonstrated that rats made anaemic with an iron-deficient diet presented with decreased exercise performance, and the subsequent repletion of iron improved functional capacity along with an increase in haemoglobin.68 Indeed, in several animal studies, low haemoglobin concentration (in the course of IDA) correlated with reduced aerobic capacity.65 Importantly, some studies showed that the relationship may be non-linear,66 and factors other than decreased haemoglobin may play a role.67 Nevertheless, studies on iron repletion in experimental animals with IDA further confirm an association between haemoglobin concentration and maximal oxygen consumption (VO2max ).51 Regarding ID without anaemia, the experiments of Finch et al.50 are of a particular relevance. The authors compared the running ability in dietary iron-deficient and control rats.50 In the first experiment, the exchange blood transfusion procedure lowered the haemoglobin concentration to 6 g/dL, which resulted in a reduction of the running time in animals both with and without previously induced ID.50 In the next experiment, the exchange blood transfusion increased the haemoglobin level to 10 g/dL, which improved the running ability in animals without ID, whereas in iron-deficient rodents the running time remained reduced.50 Importantly, impaired work capacity in iron-deficient animals with a restored haemoglobin level was thereafter quickly corrected with iron therapy.50 Furthermore, Davies et al.51 monitored the effects of dietary iron repletion in rats with severe IDA. The authors demonstrated that the normalization of haemoglobin occurred earlier and was ........................................................................................................................................................................ Iron deficiency and skeletal muscles © 2016 The Authors European Journal of Heart Failure © 2016 European Society of Cardiology accompanied by an increase in VO2max , whereas an improvement in muscle oxidative capacity and muscle mitochondrial content was delayed, and was associated with an increase in endurance capacity.51 Iron deficiency, global exercise capacity, and skeletal muscle performance in humans Links between ID, IDA, and decreased global exercise capacity have been the subject of interest in the field of sports medicine.70 Data from studies focusing on the effects of physical training suggest that ID even without anaemia can negatively affect physical performance in humans, which can be reversed by iron therapy.71 Burden et al.72 performed a meta-analysis of 17 trials regarding iron therapy in non-anaemic iron-deficient athletes and have shown that such therapy improves both systemic iron parameters and VO2max .72 Importantly, although in experimental animals ID negatively impacts skeletal muscle functioning, analogous data regarding humans are limited, and particular studies have yielded inconsistent results. With regard to interventional studies, it has been shown that beneficial effects of iron repletion in both untrained subjects and athletes with ID but without anaemia comprise increased endurance capacity and improved energetic efficiency.73 – 76 Conversely, some observational studies on IDA in humans have failed to confirm the associations between ID and muscle dysfunction. For example, Thompson et al. investigated skeletal muscle bioenergetics in women with IDA using 31 P magnetic resonance spectroscopy, and have not documented significant mitochondrial abnormality.77 Furthermore, Celsing et al. investigated the biochemical properties of skeletal muscle samples obtained from subjects with chronic moderate to severe IDA and controls, and have demonstrated that the maximal activities of glycolytic and oxidative enzymes in these two groups were similar.78 Importantly, the aforementioned studies were observational only and the study groups were relatively small. Therefore, these studies may not be sufficiently powered to identify subtle differences in the skeletal muscle energetics between subjects with and those without ID. Iron status, symptoms, and functional capacity in patients with chronic diseases accompanied by skeletal and respiratory myopathy Skeletal and respiratory myopathy in the course of heart failure and chronic obstructive pulmonary disease Skeletal and respiratory muscle dysfunction constitutes a common and important pathophysiological feature of both HF and COPD.3,5,79 – 82 It contributes to debilitating symptomatology of these disease syndromes (exercise intolerance), poor quality of Correction phase: 200 mg of iron Maximum 24 i.v. weekly until repletion dose weeks is achieved. Maintenance phase: 200 mg i.v. iron every 4 weeks. Randomized, LVEF ≤40% and Hb, 9.5–13.5 g/dL ID, serum ferritin double-blind, NYHA class II; <100 μg/L or placebo-controlled, LVEF ≤45% and 100–299 μg/L multicentre study NYHA class III with TSAT <20% Correction phase: 200 mg iron i.v. weekly until ferritin ≥500 mg/L. Maintenance phase: 200 mg iron i.v. every 4 weeks 16 weeks Randomized, Ferritin <100 μg/L, – NYHA class II–III, Hb <12.5 g/dL (anaemic group) open-label, or ferritin peakVO2 ≤ 18 mL/min/kg, LVEF or Hb observer-blinded, 100–300 μg/L 12.5–14.5 g/dL placebo-controlled, ≤45% with TSAT (non-anaemic double-centre <20% group) study FCM i.v. (n = 150) vs. placebo (n = 151) Correction phase: 500–2000 mg Maximum 36 (dosed at baseline and week 6). weeks Maintenance phase: 500 mg at each of weeks 12, 24, and 36, if ID was still present Maximum 12 days 5 weeks Prospective, NYHA class II-III, uncontrolled, systolic HF open-label, single-centre study Hb <12.0 g/dL No specific criteria ↑ 6MWT distance CrCl, creatinine clearance; FCM, ferric carboxymaltose, Hb, haemoglobin, HF, heart failure; ID, iron deficiency; ISC, iron sucrose; 6MWT, 6-min walking test; peakVO2 , peak oxygen consumption; TSAT, transferrin saturation. All patients (n = 16) 200 mg iron i.v. on days 1, 3, 5, treated with ISC i.v. and additional 200 mg on days (no placebo arm) 15 and 17 if serum ferritin <400 ng/mL on day 12 6. Bolger et al.111 200 mg i.v. iron weekly ISC i.v. (n = 20) vs. placebo (n = 20) 5. Toblli et al.112 – Randomized, LVEF ≤35%, Hb <12.5 g/dL for Ferritin <100 ↑ 6MWT distance – double-blind, NYHA II–IV, men and <11.5 ng/mL or TSAT placebo-controlled, CrCl ≤90 g/dL for women <20% single-centre study mL/min ................................................................................................................................................................................................................... 4. Ponikowski et al.114 , CONFIRM-HF study Randomized, NYHA class II or Hb <15.0 g/dL ID, serum ferritin All patients: ↑ – double-blind, III, LVEF ≤45%, <100 ng/mL, or 6MWT distance placebo-controlled, BNP >100 100–300 ng/mL (primary multicentre study pg/mL and/or if TSAT <20% endpoint); NT-proBNP anaemic patients: >400 pg/mL ↑ 6MWT distance ................................................................................................................................................................................................................... ISC i.v. (n = 24) vs. placebo (n = 11) All patients: ↑ peakVO2 per kg body mass (secondary endpoint); anaemic patients: ↑ peakVO2 per kg body mass, ↑ absolute peakVO2 ................................................................................................................................................................................................................... 3. Okonko et al.115 , FERRIC-HF study ISC i.v. + placebo p.o. Group 1: ISC 200 mg i.v. once a Iron/placebo i.v., Randomized, 2. Beck-da-Silva LVEF <40%, Hb: ≥9.0 and TSAT <20% and – ↑ peakVO2 per kg body mass (group 1, n = 10) vs. week (30 min infusions) for 5 et al.116 , 5 weeks; double-blind, NYHA class ≤12.0 g/dL ferritin <500 IRON-HF study ferrous sulfate p.o. weeks + oral placebo three iron/placebo placebo-controlled, II–IV μg/L + placebo i.v. (group times a day for 8 weeks. p.o., 8 weeks multicentre study 2, n = 7) vs. placebo Group 2: ferrous sulfate 200 mg p.o. + placebo i.v. p.o. three times a day for 8 (group 3, n = 6) weeks + i.v. placebo once a week for 5 weeks. Group 3: oral placebo three times a day for 8 weeks + i.v. placebo once a week for 5 weeks. ................................................................................................................................................................................................................... 1. Anker et al.113 , FCM i.v. (n = 304) vs. FAIR-HF study placebo (n = 155) (+ subanalysis, Filippatos et al.117 ) ↑ 6MWT distance – (secondary endpoint, also in anaemic and non-anaemic subjects, separately) ................................................................................................................................................................................................................... Study details Results: 6MWT Results: peakVO2 Duration of Study design Inclusion criteria Inclusion criteria Inclusion Intervention: study Iron dosing scheme criteria iron therapy regarding clinical regarding arms with numbers regarding characteristics haemoglobin of patients, iron status concentration iron compound ................................................................................................................................................................................................................... Table 1 Summary of studies investigating the effects of intravenous iron therapy without erythropoiesis-stimulating agents on indices of exercise capacity in patients with heart failure 8 M. Stugiewicz et al. © 2016 The Authors European Journal of Heart Failure © 2016 European Society of Cardiology 9 life, and predicts worse outcomes.3,5,79,80,82 Skeletal muscle mass and volume assessed in different body regions are decreased in patients with HF,81,83 and muscles demonstrate increased exertional fatigability.84,85 In histopathological evaluation, skeletal muscle tissue in HF demonstrates fatty infiltration, diminished capillary length density, fibre atrophy, and a shift regarding fibre type distribution (from type I slow aerobic fatigue-resistant fibres to type IIx fast glycolytic fibres vulnerable to fatigue; in a diaphragm, the direction is opposite, i.e. IIx→I).6,80,86 – 88 Skeletal muscle tissue in HF is characterized by altered cellular fuel homeostasis, namely a switch towards earlier glycolysis.7,89,90 Skeletal myocytes in HF also present with decreased size and number of mitochondria, a diminished surface density of mitochondrial cristae,83,86,91,92 a decreased oxidative capacity, an altered turnover of high-energy phosphates,7,87,89,90,93,94 and decreased expression of enzymes involved in different metabolic pathways.87,93 Similar abnormalities within skeletal muscles have been demonstrated in patients with COPD. COPD is characterized not only by a generalized fat-free mass loss, but also by a decrease in cross-sectional area of skeletal muscles.5,79,80,95,96 Histopathological examination of skeletal muscle tissue in COPD reveals fibre atrophy79 and a fiber type distribution shift analogous to that observed in HF.79,82,97 Mitochondrial abnormalities include a decreased number and decreased fractional area of these organelles,79 respiratory chain dysfunction, less efficient regeneration of high-energy phosphates, early exertional acidification, and diminished oxidative metabolism of glucose along with an enhanced glycolysis.5,77,79,98 – 100 Importantly, the precise pathomechanisms underlying skeletal and respiratory myopathy in HF and COPD are not fully explained, and striated muscle dysfunction is probably multifactorial, with several overlapping factors.6,80,82 Recently, particular attention has been paid to metabolic and hormonal derangements, which favour catabolic–anabolic imbalance and are followed by marked impairment of cellular energy metabolism.6,80,82,101 Additional potential causes of skeletal myopathy developing in HF and COPD include: local hypoxia and/or hypoperfusion, oxidative stress, low-grade systemic inflammation, muscle disuse, a sedentary lifestyle, administered medications, and nutritional deficits, to name but a few.6,80,82 Importantly, one of the potential pathomechanisms contributing to muscle dysfunction in the course of chronic diseases such as HF is ID.102 Iron deficiency as a potential cause of skeletal and respiratory myopathy in heart failure and chronic obstructive pulmonary disease Iron deficiency constitutes a frequent co-morbid condition in patients with HF and affects 30–60% of them.103 – 105 In an international pooled cohort of >1500 European patients with HF, ID affected 50% of subjects.104,106 Regarding COPD, only a few analogous data are available. Among 107 consecutive patients admitted to hospital due to an acute exacerbation of COPD, all 18 anaemic patients who had their systemic iron status assessed met ........................................................................................................................................................................................................................ Iron deficiency and skeletal muscles © 2016 The Authors European Journal of Heart Failure © 2016 European Society of Cardiology the aforementioned criteria of ID.107 Further, in the second parallel part of the study, the authors assessed iron parameters in 12 ambulatory anaemic patients with COPD and diagnosed ID in 11 of them.107 Iron deficiency correlates with impaired functional capacity in patients with HF, as assessed using peak oxygen consumption (VO2peak ) and ventilatory response to exercise (VE–VCO2 slope).108,109 In patients with systolic HF and concomitant ID, intravenous110 iron therapy improves functional capacity as assessed using distance in a 6-min walking test (6MWT)111 – 114 and VO2peak 115,116 (Table 1). Importantly, beneficial effects of iron supplementation are reported irrespective of the presence of anaemia.113 – 115,117 In two large randomized, double blind, placebo-controlled clinical trials on intravenous ferric carboxymaltose in patients with HF and ID (FAIR-HF and CONFIRM-HF trials113,114,117 – 119 ), iron therapy improved 6MWT distance in both anaemic and non-anaemic subjects. Regarding COPD, the effects of iron therapy on physical performance have not been investigated. Skeletal and respiratory muscle dysfunction due to ID102 constitutes a potential pathophysiological link between impaired iron status and decreased exercise capacity in patients with HF and COPD. Nevertheless, there are no direct experimental or clinical data confirming that the clinical correlation between ID and decreased exercise capacity in HF or COPD results from muscle dysfunction caused by impaired iron homeostasis. Further studies are needed to establish pathophysiologial links between ID and skeletal muscle dysfunction in these disease syndromes. Conclusions Accumulated evidence from physiological experiments in animals and interventional studies in the field of sports medicine suggest that undisturbed systemic iron homeostasis significantly contributes to the effective functioning of skeletal muscles. Although ID constitutes a frequent co-morbid condition in debilitating chronic diseases such as HF, the hypothesis of whether disordered iron homeostasis favours or aggravates skeletal and respiratory muscle dysfunction has not yet been verified. Further studies investigating pathophysiological links between ID and myopathy in chronic diseases are therefore warranted. Funding This research was financially supported by the National Science Centre (Kraków, Poland) grant allocated on the basis of decision number DEC-2012/05/E/NZ5/00590. Conflict of interest: Wroclaw Medical University received an unrestricted grant from Vifor Pharma. E.A.J. reports grants and personal fees from Vifor Pharma. P.P. reports grants and personal fees from Vifor Pharma, and personal fees from AMGEN. All other authors report no conflict of interest. References 1. Mosterd A, Hoes AW. Clinical epidemiology of heart failure. Heart 2007;93: 1137–1146. 10 ........................................................................................................................................................................................................................ 2. Mannino DM. COPD: epidemiology, prevalence, morbidity and mortality, and disease heterogeneity. Chest 2002;121(5 Suppl):121S–126S. 3. Coats AJ, Clark AL, Piepoli M, Volterrani M, Poole-Wilson PA. Symptoms and quality of life in heart failure: the muscle hypothesis. Br Heart J 1994;72(2Suppl):S36–S39. 4. Gosker HR, Wouters EF, van der Vusse GJ, Schols AM. Skeletal muscle dysfunction in chronic obstructive pulmonary disease and chronic heart failure: underlying mechanisms and therapy perspectives. Am J Clin Nutr 2000;71:1033–1047. 5. Barnes PJ, Celli BR. Systemic manifestations and comorbidities of COPD. Eur Respir J 2009;33:1165–1185. 6. Zizola C, Schulze PC. Metabolic and structural impairment of skeletal muscle in heart failure. Heart Fail Rev 2013;18:623–630. 7. Lipkin DP, Jones DA, Round JM, Poole-Wilson PA. Abnormalities of skeletal muscle in patients with chronic heart failure. Int J Cardiol 1988;18:187–195. 8. Rabinovich RA, Vilaró J. Structural and functional changes of peripheral muscles in chronic obstructive pulmonary disease patients. Curr Opin Pulm Med 2010;16:123–133. 9. Buller NP, Jones D, Poole-Wilson PA. Direct measurement of skeletal muscle fatigue in patients with chronic heart failure. Br Heart J 1991;65:20–24. 10. Minotti JR, Christoph I, Oka R, Weiner MW, Wells L, Massie BM. Impaired skeletal muscle function in patients with congestive heart failure. Relationship to systemic exercise performance. J Clin Invest 1991;88:2077–2082. 11. Beard JL. Iron biology in immune function, muscle metabolism and neuronal functioning. J Nutr 2001;131(2S-2):568S–579S. 12. Galy B, Ferring-Appel D, Sauer SW, Kaden S, Lyoumi S, Puy H, Kölker S, Gröne HJ, Hentze MW. Iron regulatory proteins secure mitochondrial iron sufficiency and function. Cell Metab 2010;12:194–201. 13. Bayeva M, Gheorghiade M, Ardehali H. Mitochondria as a therapeutic target in heart failure. J Am Coll Cardiol 2013;61:599–610. 14. Puntarulo S. Iron, oxidative stress and human health. Mol Aspects Med 2005;26: 299–312. 15. Iyengar MR. Creatine kinase as an intracellular regulator. J Muscle Res Cell Motil 1984;5:527–534. 16. Atalay M, Hänninen OOP. Muscle energy metabolism. In Atalay M, Hänninen O, ed. Encyclopedia of Life Support Systems: Physiology and Maintenance, Vol. IV. Eolss Publishers Company Limited: 2009; p. 26–47. 17. Hänninen O, Atalay M. Oxidative metabolism in skeletal muscle. In: Reznick AZ, Packer L, Sen CK, Holloszy JO, Jackson MJ, eds. Oxidative Stress in Skeletal Muscle. MCBU Molecular and Cell Biology Updates. Springer: Berlin; 1998. p.29–42. 18. Westerblad H, Bruton JD, Katz A. Skeletal muscle: energy metabolism, fiber types, fatigue and adaptability. Exp Cell Res 2010;316:3093–3099. 19. Bottinelli R, Reggiani C. Human skeletal muscle fibres: molecular and functional diversity. Prog Biophys Mol Biol 2000;73:195–262. 20. Polla B, D’Antona G, Bottinelli R, Reggiani C. Respiratory muscle fibres: specialisation and plasticity. Thorax 2004;59:808–817. 21. Mettauer B, Zoll J, Garnier A, Ventura-Clapier R. Heart failure: a model of cardiac and skeletal muscle energetic failure. Pflugers Arch Eur J Physiol 2006;452: 653–666. 22. Baldwin KM, Haddad F. Effects of different activity and inactivity paradigms on myosin heavy chain gene expression in striated muscle. J Appl Physiol 2001;90: 345–357. 23. Schiaffino S, Reggiani C. Molecular diversity of myofibrillar proteins: gene regulation and functional significance. Physiol Rev 1996;76:371–423. 24. Gollnick PD, Armstrong RB, Saubert CW 4th, Piehl K, Saltin B. Enzyme activity and fiber composition in skeletal muscle of untrained and trained men. J Appl Physiol 1972;33:312–319. 25. Ingwall JS. Energy metabolism in heart failure and remodelling. Cardiovasc Res 2009;81:412–419. 26. Traczyk WZ, Trzebski A, eds. Fizjologia Człowieka z Elementami Fizjologii Stosowanej i Klinicznej. Warszawa: Wydawnictwo Lekarskie PZWL; 2007. 27. Spriet LL. Anaerobic metabolism in human skeletal muscle during short-term, intense activity. Can J Physiol Pharmacol 1992;70:157–165. 28. Murray RK, Granner DK, Rodwell VW, eds. Harper’s Illustrated Biochemistry, 27th ed. New York: The McGraw-Hill Companies; 2006. 29. Rose AJ, Richter EA. Skeletal muscle glucose uptake during exercise: how is it regulated? Physiology 2005;20:260–270. 30. Heinonen I, Saltin B, Kemppainen J, Sipilä HT, Oikonen V, Nuutila P, Knuuti J, Kalliokoski K, Hellsten Y. Skeletal muscle blood flow and oxygen uptake at rest and during exercise in humans: a PET study with nitric oxide and cyclooxygenase inhibition. Am J Physiol Heart Circ Physiol 2011;300:H1510–H1517. 31. Hultman E. Fuel selection, muscle fibre. Proc Nutr Soc 1995;54:107–121. 32. Zhang L, Keung W, Samokhvalov V, Wang W, Lopaschuk GD. Role of fatty acid uptake and fatty acid 𝛽-oxidation in mediating insulin resistance in heart and skeletal muscle. Biochim Biophys Acta 2010;1801:1–22. M. Stugiewicz et al. 33. Boulton F. The myoglobin content of human skeletal muscle. Br J Haematol 1973;25:281. 34. Robach P, Cairo G, Gelfi C, Bernuzzi F, Pilegaard H, Viganò A, Santambrogio P, Cerretelli P, Calbet JA, Moutereau S, Lundby C. Strong iron demand during hypoxia-induced erythropoiesis is associated with down-regulation of iron-related proteins and myoglobin in human skeletal muscle. Blood 2007;109:4724–4731 35. Ohira Y, Gill SL. Effects of dietary iron deficiency on muscle fiber characteristics and whole-body distribution of hemoglobin in mice. J Nutr 1983;113:1811–188. 36. Kuninger D, Kuns-Hashimoto R, Kuzmickas R, Rotwein P. Complex biosynthesis of the muscle-enriched iron regulator RGMc. J Cell Sci 2006;119:3273–3283. 37. Gkouvatsos K, Wagner J, Papanikolaou G, Sebastiani G, Pantopoulos K. Conditional disruption of mouse Hfe2 gene: mMaintenance of systemic iron homeostasis requires hepatic but not skeletal muscle hemojuvelin. Hepatology 2011;54:1800–1807. 38. Hentze M, Muckenthaler MU, Galy B, Camaschella C. Two to tango: regulation of mammalian iron metabolism. Cell 2010;142:24–38. 39. Finberg KE. Regulation of systemic iron homeostasis. Curr Opin Hematol 2013;20:208–214. 40. Zhang AS. Control of systemic iron homeostasis by the hemojuvelin-hepcidin axis. Adv Nutr 2010;1:38–45. 41. Polonifi A, Politou M, Kalotychou V, Xiromeritis K, Tsironi M, Berdoukas V, Vaiopoulos G, Aessopos A. Iron metabolism gene expression in human skeletal muscle. Blood Cells Mol Dis 2010;45:233–237. 42. Sekyere EO, Dunn LL, Richardson DR. Examination of the distribution of the transferrin homologue, melanotransferrin (tumour antigen p97), in mouse and human. Biochim Biophys Acta 2005;1722:131–142. 43. Masso-Silva J, Diamond G, Macias-Rodriguez M, Ascencio F. Genomic organization and tissue-specific expression of hepcidin in the pacific mutton hamlet, Alphestes immaculatus (Breder, 1936). Fish Shellfish Immunol 2011;31:1297–1302. 44. Anderson CP, Shen M, Eisenstein RS, Leibold EA. Mammalian iron metabolism and its control by iron regulatory proteins. Biochim Biophys Acta 2012;1823: 1468–1483. 45. Guo B, Yu Y, Leibold EA. Iron regulates cytoplasmic levels of a novel ironresponsive element-binding protein without aconitase activity. J Biol Chem 1994;269:24252–24260. 46. Philpott CC, Leidgens S, Frey AG. Metabolic remodeling in iron-deficient fungi. Biochim Biophys Acta 2012;1823:1509–1520. 47. Ackrell B, Maguire JJ, Dallman PR, Kearney EB. Effect of iron deficiency on succinate- and NADH-ubiquinone oxidoreductases in skeletal muscle mitochondria. J Biol Chem 1984;259:10053–10059. 48. Cartier LJ, Ohira Y, Chen M, Cuddihee RW, Holloszy JO. Perturbation of mitochondrial composition in muscle by iron deficiency. J Biol Chem 1986;261: 13827–13832. 49. Mackler B, Grace R, Finch CA. Iron deficiency in the rat: effects on oxidative metabolism in distinct types of skeletal muscle. Pediatr Res 1984;18:499–500. 50. Finch CA, Miller LR, Inamdar AR, Person R, Seiler K, Mackler B. Iron deficiency in the rat. Physiological and biochemical studies of muscle dysfunction. J Clin Invest 1976;58:447–453. 51. Davies KJ, Maguire JJ, Brooks GA, Dallman PR, Packer L. Muscle mitochondrial bioenergetics, oxygen supply, and work capacity during dietary iron deficiency and repletion. Am J Physiol 1982;242:E418–E427. 52. Henderson SA, Dallman PR, Brooks GA. Glucose turnover and oxidation are increased in the iron-deficient anemic rat. Am J Physiol 1986;250:E414–E421. 53. Ohira Y, Hegenauer J, Strause L, Chen CS, Saltman P, Beinert H. Mitochondrial NADH dehydrogenase in iron-deficient and iron-repleted rat muscle: an EPR and work performance study. Br J Haematol 1982;52:623–630. 54. Thompson CH, Green YS, Ledingham JG, Radda GK, Rajagopalan B. The effect of iron deficiency on skeletal muscle metabolism of the rat. Acta Physiol Scand 1993;147:85–90. 55. Ohlendieck K. Proteomic identification of biomarkers of skeletal muscle disorders. Biomark Med 2013;7:169–186. 56. Zick M, Rabl R, Reichert AS. Cristae formation—linking ultrastructure and function of mitochondria. Biochim Biophys Acta 2009;1793:5–19. 57. Maguire JJ, Davies KJ, Dallman PR, Packer L. Effects of dietary iron deficiency of iron-sulfur proteins and bioenergetic functions of skeletal muscle mitochondria. Biochim Biophys Acta 1982;679;210–220. 58. Hagler L, Askew EW, Neville JR, Mellick PW, Coppes RI Jr, Lowder JF Jr. Influence of dietary iron deficiency on hemoglobin, myoglobin, their respective reductases, and skeletal muscle mitochondrial respiration. Am J Clin Nutr 1981;34:2169–2177. 59. Merrill JF, Thomson DM, Hardman SE, Hepworth SD, Willie S, Hancock CR. Iron deficiency causes a shift in AMP-activated protein kinase (AMPK) subunit composition in rat skeletal muscle. Nutr Metab (Lond) 2012;9:104. © 2016 The Authors European Journal of Heart Failure © 2016 European Society of Cardiology 11 60. Liew YF, Shaw NS. Mitochondrial cysteine desulfurase iron–sulfur cluster S and aconitase are post-transcriptionally regulated by dietary iron in skeletal muscle of rats. J Nutr 2005;135:2151–2158. 61. O’Dell BL, Sunde RA. Handbook of Nutritionally Essential Mineral Elements. Boca Raton, FL: CRC Press; 1997 62. Han DH, Hancock CR, Jung SR, Higashida K, Kim SH, Holloszy JO. Deficiency of the mitochondrial electron transport chain in muscle does not cause insulin resistance. PLoS One 2011;6:e19739. 63. Winder WW, Holmes BF, Rubink DS, Jensen EB, Chen M, Holloszy JO. Activation of AMP-activated protein kinase increases mitochondrial enzymes in skeletal muscle. J Appl Physiol 2000;88:2219–2226. 64. McGee SL, Mustard KJ, Hardie DG, Baar K. Normal hypertrophy accompanied by phosphoryation and activation of AMP-activated protein kinase alpha1 following overload in LKB1 knockout mice. J Physiol 2008;586:1731–1741. 65. Haas JD, Brownlie T 4th. Iron deficiency and reduced work capacity: a critical review of the research to determine a causal relationship. J Nutr 2001;131(2S-2):676S–688S; discussion 688S-690S. 66. Perkkiö MV, Jansson LT, Brooks GA, Refino CJ, Dallman PR. Work performance in iron deficiency of increasing severity. J Appl Physiol 1985;58:1477–1480. 67. Davies KJ, Donovan CM, Refino CJ, Brooks GA, Packer L, Dallman PR. Distinguishing effects of anemia and muscle iron deficiency on exercise bioenergetics in the rat. Am J Physiol 1984;246:E535–E543. 68. Edgerton VR, Bryant SL, Gillespie CA, Gardner GW. Iron deficiency anemia and physical performance and activity of rats. J Nutr 1972;102:381–399. 69. Ohira Y, Koziol BJ, Edgerton VR, Brooks GA. Oxygen consumption and work capacity in iron-deficient anemic rats. J Nutr 1981;111:17–25. 70. Beard J, Tobin B. Iron status and exercise. Am J Clin Nutr 2000;72(2 Suppl):594S–597S. 71. Brownlie T 4th, Utermohlen V, Hinton PS, Giordano C, Haas JD. Marginal iron deficiency without anemia impairs aerobic adaptation among previously untrained women. Am J Clin Nutr 2002;75:734–742. 72. Burden RJ, Morton K, Richards T, Whyte GP, Pedlar CR. Is iron treatment beneficial in, iron-deficient but non-anaemic (IDNA) endurance athletes? A meta-analysis. Br J Sports Med 2015;49:1389–1397. 73. Hinton PS, Giordano C, Brownlie T, Haas JD. Iron supplementation improves endurance after training in iron-depleted, nonanemic women. J Appl Physiol 2000;88:1103–1111. 74. Hinton PS, Sinclair LM. Iron supplementation maintains ventilatory threshold and improves energetic efficiency in iron-deficient nonanemic athletes. Eur J Clin Nutr 2007;61:30–39. 75. Brutsaert TD, Hernandez-Cordero S, Rivera J, Viola T, Hughes G, Haas JD. Iron supplementation improves progressive fatigue resistance during dynamic knee extensor exercise in iron-depleted, nonanemic women. Am J Clin Nutr 2003;77:441–448. 76. Brownlie T 4th, Utermohlen V, Hinton PS, Haas JD. Tissue iron deficiency without anemia impairs adaptation in endurance capacity after aerobic training in previously untrained women. Am J Clin Nutr 2004;79:437–443. 77. Thompson CH, Kemp GJ, Taylor DJ, Radda GK, Rajagopalan B. No evidence of mitochondrial abnormality in skeletal muscle of patients with iron-deficient anaemia. J Intern Med 1993;234:149–154. 78. Celsing F, Blomstrand E, Werner B, Pihlstedt P, Ekblom B. Effects of iron deficiency on endurance and muscle enzyme activity in man. Med Sci Sports Exerc 1986;18:156–161. 79. Gosker HR, Hesselink MK, Duimel H, Ward KA, Schols AM. Reduced mitochondrial density in the vastus lateralis muscle of patients with COPD. Eur Respir J 2007;30:73–79. 80. Gosker HR, Wouters EF, van der Vusse GJ, Schols AM. Skeletal muscle dysfunction in chronic obstructive pulmonary disease and chronic heart failure: underlying mechanisms and therapy perspectives. Am J Clin Nutr 2000;71:1033–1047. 81. Piepoli MF, Kaczmarek A, Francis DP, Davies LC, Rauchhaus M, Jankowska EA, Anker SD, Capucci A, Banasiak W, Ponikowski P. Reduced peripheral skeletal muscle mass and abnormal reflex physiology in chronic heart failure. Circulation 2006;114:126–134. 82. Agustí AG, Noguera A, Sauleda J, Sala E, Pons J, Busquets X. Systemic effects of chronic obstructive pulmonary disease. Eur Respir J 2003;21:347–360. 83. Mancini DM, Walter G, Reichek N, Lenkinski R, McCully KK, Mullen JL, Wilson JR. Contribution of skeletal muscle atrophy to exercise intolerance and altered muscle metabolism in heart failure. Circulation 1992;85:1364–1373. 84. Buller NP, Jones D, Poole-Wilson PA. Direct measurement of skeletal muscle fatigue in patients with chronic heart failure. Br Heart J 1991;65:20–24. 85. Harridge SD, Magnusson G, Gordon A. Skeletal muscle contractile characteristics and fatigue resistance in patients with chronic heart failure. Eur Heart J 1996;17:896–901. 86. Drexler H, Riede U, Münzel T, König H, Funke E, Just H. Alterations of skeletal muscle in chronic heart failure. Circulation 1992;85:1751–1759. ........................................................................................................................................................................................................................ Iron deficiency and skeletal muscles © 2016 The Authors European Journal of Heart Failure © 2016 European Society of Cardiology 87. Sullivan MJ, Green HJ, Cobb FR. Skeletal muscle biochemistry and histology in ambulatory patients with long-term heart failure. Circulation 1990;81:518–527. 88. Sabbah HN, Hansen-Smith F, Sharov VG, Kono T, Lesch M, Gengo PJ, Steffen RP, Levine TB, Goldstein S. Decreased proportion of type I myofibers in skeletal muscle of dogs with chronic heart failure. Circulation 1993;87:1729–1737. 89. Chati Z, Zannad F, Robin-Lherbier B, Escanye JM, Jeandel C, Robert J, Aliot E. Contribution of specific skeletal muscle metabolic abnormalities to limitation of exercise capacity in patients with chronic heart failure: a phosphorus 31 nuclear magnetic resonance study. Am Heart J 1994;128:781–792. 90. Mancini DM, Coyle E, Coggan A, Beltz J, Ferraro N, Montain S, Wilson JR. Contribution of intrinsic skeletal muscle changes to 31P NMR skeletal muscle metabolic abnormalities in patients with chronic heart failure. Circulation 1989;80:1338–1346. 91. Mancini DM, Henson D, LaManca J, Levine S. Respiratory muscle function and dyspnea in patients with chronic congestive heart failure. Circulation 1992;86 909–918. 92. Mancini DM, Ferraro N, Tuchler M, Chance B, Wilson JR. Detection of abnormal calf muscle metabolism in patients with heart failure using phosphorus-31 nuclear magnetic resonance. Am J Cardiol 1988;62:1234–1240. 93. Mettauer B, Zoll J, Sanchez H, Lampert E, Ribera F, Veksler V, Bigard X, Mateo P, Epailly E, Lonsdorfer J, Ventura-Clapier R. Oxidative capacity of skeletal muscle in heart failure patients versus sedentary or active control subjects. J Am Coll Cardiol 2001;38:947–954. 94. Massie BM, Conway M, Yonge R, Frostick S, Sleight P, Ledingham J, Radda G, Rajagopalan B. 31P nuclear magnetic resonance evidence of abnormal skeletal muscle metabolism in patients with congestive heart failure. Am J Cardiol 1987;60:309–315. 95. Agusti A, Soriano JB. COPD as a systemic disease. COPD 2008;5:133–138. 96. Bernard S, LeBlanc P, Whittom F, Carrier G, Jobin J, Belleau R, Maltais F. Peripheral muscle weakness in patients with chronic obstructive pulmonary disease. Am J Respir Crit Care Med 1998;158:629–634. 97. Levine S, Kaiser L, Leferovich J, Tikunov B. Cellular adaptations in the diaphragm in chronic obstructive pulmonary disease. N Engl J Med 1997;337:1799–1806. 98. Puente-Maestu L, Pérez-Parra J, Godoy R, Moreno N, Tejedor A, González-Aragoneses F, Bravo JL, Alvarez FV, Camaño S, Agustí A. Abnormal mitochondrial function in locomotor and respiratory muscles of COPD patients. Eur Respir J 2009;33:1045–1052. 99. Wuyam B1, Payen JF, Levy P, Bensaïdane H, Reutenauer H, Le Bas JF, Benabid AL. Metabolism and aerobic capacity of skeletal muscle in chronic respiratory failure related to chronic obstructive pulmonary disease. Eur Respir J 1992;5:157–162. 100. Payen JF, Wuyam B, Levy P, Reutenauer H, Stieglitz P, Paramelle B, Le Bas JF. Muscular metabolism during oxygen supplementation in patients with chronic hypoxemia. Am Rev Respir Dis 1993;147:592–598. 101. Josiak K1, Jankowska EA, Piepoli MF, Banasiak W, Ponikowski P. Skeletal myopathy in patients with chronic heart failure: significance of anabolic–androgenic hormones. J Cachexia Sarcopenia Muscle 2014;5:287–296. 102. McMurray JJ, Adamopoulos S, Anker SD, Auricchio A, Bohm M, Dickstein K, Falk V, Filippatos G, Fonseca C, Gomez-Sanchez MA, Jaarsma T, Kober L, Lip GY, Maggioni AP, Parkhomenko A, Pieske BM, Popescu BA, Ronnevik PK, Rutten FH, Schwitter J, Seferovic P, Stepinska J, Trindade PT, Voors AA, Zannad F, Zeiher A, Bax JJ, Baumgartner H, Ceconi C, Dean V, Deaton C, Fagard R, Funck-Brentano C, Hasdai D, Hoes A, Kirchhof P, Knuuti J, Kolh P, McDonagh T, Moulin C, Reiner Z, Sechtem U, Sirnes PA, Tendera M, Torbicki A, Vahanian A, Windecker S, Bonet LA, Avraamides P, Ben Lamin HA, Brignole M, Coca A, Cowburn P, Dargie H, Elliott P, Flachskampf FA, Guida GF, Hardman S, Iung B, Merkely B, Mueller C, Nanas JN, Nielsen OW, Orn S, Parissis JT, Ponikowski P. ESC guidelines for the diagnosis and treatment of acute and chronic heart failure 2012: the Task Force for the Diagnosis and Treatment of Acute and Chronic Heart Failure 2012 of the European Society of Cardiology. Developed in collaboration with the Heart Failure Association (HFA) of the ESC. Eur J Heart Fail 2012;14:803–869. 103. Cohen-Solal A, Damy T, Terbah M, Kerebel S, Baguet JP, Hanon O, Zannad F, Laperche T, Leclercq C, Concas V, Duvillié L, Darné B, Anker S, Mebazaa A. High prevalence of iron deficiency in patients with acute decompensated heart failure. Eur J Heart Fail 2014;16:984–991. 104. Klip IT, Comin-Colet J, Voors AA, Ponikowski P, Enjuanes C, Banasiak W, Lok DJ, Rosentryt P, Torrens A, Polonski L, van Veldhuisen DJ, van der Meer P, Jankowska EA. Iron deficiency in chronic heart failure: an international pooled analysis. Am Heart J 2013;165:575–582. 105. Yeo TJ, Yeo PS, Ching-Chiew Wong R, Ong HY, Leong KT, Jaufeerally F, Sim D, Santhanakrishnan R, Lim SL, M Y Chan M, Chai P, Low AF, Ling LH, Ng TP, Richards AM, Lam CS. Iron deficiency in a multi-ethnic Asian population with and without heart failure: prevalence, clinical correlates, functional significance and prognosis. Eur J Heart Fail 2014;16:1125–1132. 12 ................................................................... 106. Klip IT, Jankowska EA, Enjuanes C, Voors AA, Banasiak W, Bruguera J, Rozentryt P, Polonski L, van Veldhuisen DJ, Ponikowski P, Comin-Colet J, van der Meer P. The additive burden of iron deficiency in the cardiorenal–anaemia axis: scope of a problem and its consequences. Eur J Heart Fail 2014;16:655–662. 107. Silverberg DS, Mor R, Weu MT, Schwartz D, Schwartz IF, Chernin G. Anemia and iron deficiency in COPD patients: prevalence and the effects of correction of the anemia with erythropoiesis stimulating agents and intravenous iron. BMC Pulm Med 2014;14:24. 108. Jankowska EA, Rozentryt P, Witkowska A, Nowak J, Hartmann O, Ponikowska B, Borodulin-Nadzieja L, von Haehling S, Doehner W, Banasiak W, Polonski L, Filippatos G, Anker SD, Ponikowski P. Iron deficiency predicts impaired exercise capacity in patients with systolic chronic heart failure. J Card Fail 2011;17: 899–906. 109. Okonko DO, Mandal AK, Missouris CG, Poole-Wilson PA. Disordered iron homeostasis in chronic heart failure: prevalence, predictors, and relation to anemia, exercise capacity, and survival. J Am Coll Cardiol 2011;58: 1241–1251. 110. McDonagh T, Macdougall IC. Iron therapy for the treatment of iron deficiency in chronic heart failure: intravenous or oral? Eur J Heart Fail 2015;17:248–262. 111. Bolger AP, Bartlett FR, Penston HS, O’Leary J, Pollock N, Kaprielian R, Chapman CM. Intravenous iron alone for the treatment of anemia in patients with chronic heart failure. J Am Coll Cardiol 2006;48:1225–1227. 112. Toblli JE, Lombraña A, Duarte P, Di Gennaro F. Intravenous iron reduces NT-pro-brain natriuretic peptide in anemic patients with chronic heart failure and renal insufficiency. J Am Coll Cardiol 2007;50:1657–1665. 113. Anker SD, Comin Colet J, Filippatos G, Willenheimer R, Dickstein K, Drexler H, Lüscher TF, Bart B, Banasiak W, Niegowska J, Kirwan BA, Mori C, von Eisenhart Rothe B, Pocock SJ, Poole-Wilson PA, Ponikowski P; FAIR-HF Trial Investigators. Ferric carboxymaltose in patients with heart failure and iron deficiency. N Engl J Med 2009;361:2436–2448. M. Stugiewicz et al. 114. Ponikowski P, van Veldhuisen DJ, Comin-Colet J, Ertl G, Komajda M, Mareev V, McDonagh T, Parkhomenko A, Tavazzi L, Levesque V, Mori C, Roubert B, Filippatos G, Ruschitzka F, Anker SD; CONFIRM-HF Investigators. Beneficial effects of long-term intravenous iron therapy with ferric carboxymaltose in patients with symptomatic heart failure and iron deficiency. Eur Heart J 2015;36: 657–668. 115. Okonko DO, Grzeslo A, Witkowski T, Mandal AK, Slater RM, Roughton M, Foldes G, Thum T, Majda J, Banasiak W, Missouris CG, Poole-Wilson PA, Anker SD, Ponikowski P. Effect of intravenous iron sucrose on exercise tolerance in anemic and nonanemic patients with symptomatic chronic heart failure and iron deficiency FERRIC-HF: a randomized, controlled, observer-blinded trial. J Am Coll Cardiol 2008;51:103–112. 116. Beck-da-Silva L, Piardi D, Soder S, Rohde LE, Pereira-Barretto AC, de Albuquerque D, Bocchi E, Vilas-Boas F, Moura LZ, Montera MW, Rassi S, Clausell N. IRON-HF study: a randomized trial to assess the effects of iron in heart failure patients with anemia. Int J Cardiol 2013;168:3439–3442. 117. Filippatos G, Farmakis D, Colet JC, Dickstein K, Lüscher TF, Willenheimer R, Parissis J, Gaudesius G, Mori C, von Eisenhart Rothe B, Greenlaw N, Ford I, Ponikowski P, Anker SD. Intravenous ferric carboxymaltose in iron-deficient chronic heart failure patients with and without anaemia: a subanalysis of the FAIR-HF trial. Eur J Heart Fail 2013;15:1267–1276. 118. Van Craenenbroeck EM, Conraads VM, Greenlaw N, Gaudesius G, Mori C, Ponikowski P, Anker SD. The effect of intravenous ferric carboxymaltose on red cell distribution width: a subanalysis of the FAIR-HF study. Eur J Heart Fail 2013;15:756–762. 119. Ponikowski P, Filippatos G, Colet JC, Willenheimer R, Dickstein K, Lüscher T, Gaudesius G, von Eisenhart Rothe B, Mori C, Greenlaw N, Ford I, Macdougall I, Anker SD; FAIR-HF Trial Investigators. The impact of intravenous ferric carboxymaltose on renal function: an analysis of the FAIR-HF study. Eur J Heart Fail 2015;17:329–339. © 2016 The Authors European Journal of Heart Failure © 2016 European Society of Cardiology