Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
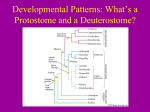
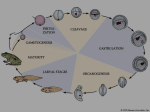
BIOLOGY 52/SECTION 7 DEVELOPMENT-DR. LILJEGREN Lecture 3 Early Cleavage Patterns Fertilization>Cleavage>Gastrulation>Organ Formation. Cleavage and Gastrulation are two critical stages of embryogenesis. 1. After fertilization, early development in most animals starts with a series of rapid cell divisions called CLEAVAGE that serves several purposes: a. rapidly increases cell number, dividing the cytoplasm of the egg into smaller and smaller cells called blastomeres i. Length of "cell cycle" is regulated in most cell types. ii. In the early embryo it's much shorter- many divisions very rapidly iii. How so fast? No growth phases (just mitosis and DNA synthesis) & relies on stored material- no transcription. So faster, and no volume increases iv. Transition from fertilization to cleavage caused by activation of MPF (mitosis-promoting factor). Active MPF dependent on presence of cyclin B, which accumulates during S and is then degraded after cells reach M phase. v. Cyclin regulators stored in egg cytoplasm, so cell cycle doesn’t require zygotic transcription for numerous cell divisions. b. distributes egg contents, often asymmetrically, to the cells 2. What controls the process of cleavage? a. The cleavage plane forms perpendicular to the mitotic spindles (microtubules made of tubulin). b. The contractile ring (microfilaments made of actin) that forms creates a cleavage furrow. c. Placement of the mitotic spindle is regulated by attachment sites that form on the inside of the cell membrane. The composition of the sites is unknown, but in some animals is controlled by cytoplasmic factors made by mom. This is an example of a MATERNAL EFFECT, in which the phenotype of the EMBRYOS is controlled by the genotype of the MOM. 3. Different organisms have different patterns of cleavage. Cleavage is quite variable among organisms, but like most developmental processes the underlying mechanisms and goals are similar. The type of cleavage primarily depends on: a. the amount and location of yolk in the egg Cell divisions occur at a faster rate in yolk-free parts of egg, in general yolk represses cleavage. Yolk-rich pole = vegetal pole, vs. animal pole 1. Completeness of cleavage • HOLOBLASTIC is complete cleavage, cleavage furrow extends through the entire egg. Sea urchins, snails, frogs, mammals. • MEROBLASTIC is partial cleavage (usually stops when it hits yolk). Yolk inhibits membrane formation. Flies, fish & birds 2. Pattern of cleavage • ISOLECITHAL (=equal yolk) eggs have a little yolk that is evenly distributed. Sea urchins, nematodes (C. elegans), mammals (including mouse and human). Since little yolk, embryos have to obtain food elsewhere. • TELOLECITHAL eggs have a lot of yolk with only one small spot that is free of yolk. This is where the embryo develops. Fish (zebrafish) and birds (chick) • CENTROLECITHAL eggs have yolk in the egg center. Most insects, like flies. b. factors in the egg cytoplasm that influence the angle of the mitotic spindle and the timing of its formation. 1. The relationship of the cleavage planes to each other and egg axis: • MERIDIONAL cuts through embryo poles like a geographic meridian • EQUATORIAL cuts through the area of greatest diameter 2. timing of mitotic spindle formation • cleavages can be SYNCHRONOUS, occuring at the same time. ie. sea urchin • or ASYNCHRONOUS, occuring at different times. ie. mammals 3. symmetric (equal): daughter cells are same size vs. assymetric (unequal) cell divisions: some daughter cells are a different size 4. Molluscs and Spiral cleavage: a. Unlike radial cleavage, cleavage planes are not parallel or perpendicular to the animal-vegetal axis of the egg, rather cleavage planes are oblique. b. Cells touch one another at more places c. Fewer cell divisions before gastrulation so easier to make fate maps 5. Frogs (A type of HOLOBLASTIC cleavage): displaced radial cleavage due to more yolk (moderate yolk disposition--between isolecithal and telolecithal). Cell divisions much slower as can be seen by the cleavage furrows. 6. Fish and bird cleavage: a. Fish and bird eggs are telolecithal (lot of yolk with only one small spot free of yolk). So cell divisions only occur in the small disc of cytoplasm=DISCOIDAL cleavage pattern 7. Insect (fly) cleavage: a. Fly embryos are CENTROLECITHAL (yolk at center of egg). They have a central yolk-rich ENDOPLASM and a peripheral yolk-free area called the PERIPLASM. Cells do not divide until after several rounds of nuclear division. b. Laser confocal micrographs showing stained chromatin. Nuclei start to divide in the endoplasm, but move to the periplasm, where they align against the outer membrane. Divisions of the cytoplasm only occur in the cell’s peripheral region=SUPERFICIAL cleavage pattern. c. POLE CELL NUCLEI get to posterior end first, they are pinched off to form POLE CELLS that will be the germ line of the embryo. d. rest of nuclei at edge make incomplete cleavage planes after 13 rounds of mitosis. This is the CELLULAR BLASTODERM STAGE. Finally, nuclei are pinched off into cells. 8. Mammalian cleavage is unique in several ways: (most studies have used mice as model organisms) a. First, initial cell divisions in mammalian eggs are slow (12-24 hours) compared to embryos that develop outside the mother (ie. flies 10 minutes), presumably because mammalian embryos are protected. b. c. d. e. f. g. h. Second, mammals have a ROTATIONAL cleavage pattern. In mammals, the first cleavage is MERIDIONAL, but during the second cleavage, one blastomere divides meridionally and the other blastomere divides equatorially. Third, mammalian cleavage is asynchronous from an early timepoint. In other words, mammlian blastomeres do not all divide at the same time. So, embryos frequently contain odd numbers of cells, ie. 3 or 6 blastomeres, instead of exponential 2-to 4-to 8-cell stages. Fourth, unlike most other animals, zygotic transcription occurs during early cleavage in mammals, and produces proteins necessary for subsequent cleavages to occur. In mice, this switch from maternal to zygotic control occurs at the 2-cell stage. Fifth, at the 8 cell stage the mammalian embryo undergoes COMPACTION. The outer cells form tight junctions with each other, while the inner cells form gap junctions, enabling small molecules and ions to pass between them. Compaction requires the cell adhesion protein E-cadherin. The 16 cell stage embryo is called the morula, and it has a small group of internal cells surrounded by a larger group of outer cells. This environment of outer vs. inner cells sets the stage for differentiation in development. Most of the descendents of the outer cells form the TROPHOBLAST (trophectoderm), while the inner cells become the INNER CELL MASS. These cells will give rise to all parts of the embryo proper and some extraembryonic structures, while the trophoblast cells will participate in placenta formation and implantation. This distinction between trophoblast and inner cell mass cell fate is the first differentiation event in mammalian development. The BLASTOCYST forms at 32-64 cells by the outer (trophoblast) cells secreting fluid into the morula to form a cavity called the BLASTOCOEL. Hatching from the zona pellucida is prevented until the blastocyst reaches the uterus. Premature hatching/implantation in the oviduct is known as ectopic or tubal pregnancy and can cause a life-threatening hemorrhage. 9. After fixed number of divisions, Cleavage ends in a controlled manner called the MID-BLASTULA TRANSITION (MBT) a. if it ended haphazardly there would be huge differences in cell numbers among embryos of the same species. b. controlled by the nuclear/cytoplasmic ratio, which is almost 0 in a fertilized egg with tons of cytoplasm, but approaches 1 in cells after many nuclei are made without extra cytoplasm. c. during the MBT the cell cycle lengthens and often G1 is used for the first time. d. the embryo starts to synthesize its own RNA (often maternal RNA is actively destroyed at this time). Remember this is a difference between mammals and other animals e. the cells become motile (prelude to gastrulation). 10. Maternal effect mutations and cleavage: SNAILS a. In a certain kind of snail, the shell normally spirals in a right or DEXTRAL COIL. Mutants make embryos with left or SINISTRAL COILS. b. The direction of the coils follows the cleavage pattern. Remember that the cleavage plane forms perpendicular to the mitotic spindles, whose placement is regulated by the spindle attachment sites. c. In snails, cytoplasmic factors made by mom determine these attachment sites. d. Alfred Sturtevant came up with the idea in 1923 that left-coiling snails are caused by a recessive maternal effect mutation, ie. the phenotype of the EMBRYOS is controlled by the genotype of MOM. Since the mutation is recessive, this means that homozygous D/D mothers and heterozygous D/d mothers make right-handed embryos, and homozygous d/d mothers make left-handed embryos. e. How does this work? (to be continued)