Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
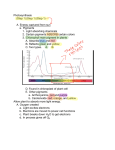
Journal of Plankton Research Vol.22 no.2 pp.305–319, 2000 Grazing experiments with two freshwater zooplankters: fate of chlorophyll and carotenoid pigments E.Pandolfini1, I.Thys2, B.Leporcq1 and J.-P.Descy1,3 1Laboratory of Freshwater Ecology, Department of Biology, FUNDP, B-5000 Namur, Belgium and 2CREBS, Centre de Recherches Public—Centre Universitaire (CRP/CU), 162a, av. de la Faïencerie, L-1511 Luxembourg 3To whom correspondence should be addressed Abstract. In order to evaluate the validity of the gut pigment method to assess grazing and diet in two freshwater zooplankters, experiments were carried out to check chlorophyll a and xanthophyll conservation during feeding. For both animals, two sets of experiments were conducted by incubating animals in the laboratory, either isolated from a reservoir (the calanoid copepod, Eudiaptomus gracilis) or cultured under high-food conditions (the cladoceran, Daphnia galeata). For both animals, gut pigments and clearance rates on different types of algae were determined from the same incubations. Chlorophyll a and derivatives, as well as major algal carotenoids, were analysed by High Performance Liquid Chromatography (HPLC). In copepods, the pigment profiles from the gut extracts reflected the diet of the animals poorly. The animal extracts contained almost exclusively alloxanthin (or an alloxanthin-like pigment) in large amounts, whereas the other pigments were lost in high proportions (>70% for lutein and fucoxanthin; 57 and 78% for a-phorbins). The cladocerans fed on the main types of algae abundant in the suspensions, with a preference, however, for small cells. Although the main xanthophylls from these algae were detected in the Daphnia extracts, some destruction of lutein and fucoxanthin may have occurred (18.7 and 30%). The loss rate for alloxanthin seemed more variable (0 and 68%), possibly depending on food concentration. As for the transformation of a-phorbins, E.gracilis and D.galeata behaved quite differently. The HPLC profiles of copepod extracts always showed a very small chlorophyll a peak, along with phaeophytin a and pyrophaeophytin a. Those from the cladoceran exhibited a large phaeophorbide a peak, along with some chlorophyll a and phaeophytin a. In fact, D.galeata did not destroy a-phorbins under our experimental conditions but converted chlorophyll a mainly into phaeophorbide. From a comparison of our results with data from other studies, it seems that in these two zooplankters, use of gut pigment data for quantitative grazing assessment should be considered with caution. Introduction The detailed analysis of algal pigments by High Performance Liquid Chromatography (HPLC) has increasingly been used in marine and freshwater ecology, for instance, in quantitative studies of phytoplankton composition and in detailed investigations of particulate fluxes to the sediment [review in (Millie et al., 1993)]. In particular, the ‘gut pigment method’, based on measurements of chlorophyll a and derivatives in zooplankton extracts, has been used widely for quantitative assessment of grazing by marine zooplankton. Indeed, chlorophyll a and derivatives found in animals’ guts were thought to provide reliable estimates of the algal biomass ingested by applying excretion rates determined experimentally to pigment amount per animal [e.g. (Mackas and Bohrer, 1976; Bautista and Harris, 1992; Landry et al., 1994)]. These techniques also seemed promising to study zooplankton grazing selectivity through the detection of carotenoid pigments specific to various algae classes in zooplankton extracts. However, there has been increasing evidence in the last decade that a-phorbins (i.e. chlorophyll a and phaeopigments) are extensively destroyed © Oxford University Press 2000 305 E.Pandolfini et al. during gut passage in marine copepods [(e.g. (Conover et al., 1986; Lopez et al., 1988; Penry and Frost, 1991; Mayzaud and Razouls, 1992; Head and Harris, 1992, 1994, 1996)], to such an extent that most authors agree on the need for re-evaluating the ‘gut pigment method’ (Drits, 1997). Equations have been proposed to correct for the destruction occurring at various stages of the feeding process in copepods (Stevens and Head, 1998) or in other marine herbivores (Bochdansky et al., 1998). Early studies on algal xanthophylls in marine zooplankton (Kleppel and Pieper, 1984; Kleppel et al., 1988, 1991; Nelson, 1989) suggested that carotenoids were more stable than chlorophylls in marine mesozooplankton guts, while degraded forms were detected in the faeces of macrozooplankton. It was therefore suggested that algal xanthophylls were well conserved in small planktonic herbivores. Subsequently, studies of selective grazing by freshwater zooplankton have been carried out, assuming complete carotenoid conservation in copepods, cladocerans and small zooplankton (Quiblier et al., 1994; Quiblier-Lloberas et al., 1996). However, views on the fate of algal carotenoids after zooplankton feeding have changed considerably. In marine copepods, Head and Harris suggested that fucoxanthin and 199hexanoyloxyfucoxanthin from diatoms and prymnesiophytes were extensively destroyed during gut passage (Head and Harris, 1992, 1994). Similar conclusions have been drawn from experimental studies on marine and freshwater calanoids; Kleppel showed substantial loss of fucoxanthin in Acartia californiensis and Calanus pacificus at high and low food levels (Kleppel, 1998), while Descy et al. found evidence of differential destruction of primary algal xanthophylls in the freshwater species Diaptomus minutus (Descy et al., 1999). Furthermore, Strom et al. have shown that both chlorophylls and carotenoids may be destroyed during feeding by microzooplankton, and that the extent of the degradation seems inversely related to protozoan size (Strom et al., 1998). Thus, the emerging picture from these few studies on the fate of carotenoids in zooplankton is totally different from what it was a few years ago, and the reliability of xanthophylls as markers of grazing by zooplankton may be questioned as much as for chlorophylls. Few experimental studies have been conducted on the fate of carotenoids in freshwater zooplankton to date. If the gut pigment technique should be applied with some caution in calanoid copepods, even less is known about how herbivorous cladocerans process algal pigments. Quiblier et al. conducted experiments on D.magna fed with mixtures of algae and observed a good correspondence between the carotenoid profiles in the animals and their food (Quiblier et al., 1994). Chlorophyll a was converted essentially to phaeophorbides and, probably, to ‘colourless’ products. However, when Daphnia was fed with lake water dominated by cyanobacteria, chlorophyll a was not degraded. Similar observations, i.e. decrease of chlorophyll a and increase of phaeophorbides a in feeding suspensions, were made by Fundel et al. who conducted long-term experiments with small and large Daphnia from Lake Constance (initial chlorophyll a range: 1.2–16.3 µg l–1) (Fundel et al., 1998). A good relationship between the phaeophorbide a/chlorophyll a ratio and Daphnia density was observed at low grazer density, but this ratio tended to decrease at high cladoceran density, suggesting 306 Pigments in freshwater zooplankton chlorophyll a destruction rather than conversion to phaeopigment at low food levels. This study presents results from feeding experiments with two common freshwater zooplankters, the calanoid copepod Eudiaptomus gracilis and the herbivorous cladoceran Daphnia galeata. We aimed to investigate the fate of algal chlorophylls and carotenoids after zooplankton feeding, and to determine classspecific clearance rates from the decrease of the amounts of specific carotenoids in the feeding suspensions (for copepods) or using algal counts in the control and feeding suspensions (for cladocerans). In particular, using actual clearance of algal groups from the suspension as a reference, we tried to assess the validity of gut pigment analysis for measuring grazing rates and for investigating the diet of these two species. Method Study lake and experimental material and design Both species were isolated from the lake of Esch-sur-Sûre, a reservoir located in the northern part of the Grand Duchy of Luxembourg. The lake is dimictic and has been considered as meso-eutrophic (annual mean chlorophyll a concentration in the euphotic zone: 5.0–8.8 µg l–1; annual maximum chlorophyll a concentration: 16.3–34.9 µg l–1; annual mean Secchi depth 4.2–4.7 m; annual minimum Secchi depth: 1.8–2.5m). The reservoir is the main drinking water supply for the country. Zooplankton dynamics have been followed for several years: in the summer zooplankton, Eudiaptomus gracilis and Daphnia galeata were often dominant species (Dohet and Hoffmann, 1995). In 1998, when this study was carried out, phytoplankton presented the following variations. After a spring dominance by Chrysophytes and diatoms, a clear-water phase developed in late May and June with mainly Cryptophytes remaining in the phytoplankton; this phase was followed by a return of Chrysophytes in July and by the development of various cyanobacteria, which became dominant in late summer. Chlorophytes were present throughout the growing season and dinoflagellates and Euglenophytes appeared sporadically. Chlorophyll a levels, measured by HPLC (see below) in epilimnetic phytoplankton <100 µm, were between 2.5 and 30 µg l–1. The copepods used in the feeding experiments were picked, using Pasteur pipettes under a dissecting microscope, from a zooplankton sample collected in the lake with a closing plankton net (53 µm mesh size). On average, 50–70 adult copepods (males and egg-bearing females in equivalent numbers, as far as possible) were placed in PVC cylinders with a 150 µm Nitex screen fitted to the bottom and suspended in 250 ml beakers filled with lake water sieved through a 63 µm screen in an incubator set at the lake temperature. The animals were allowed to acclimate, in the dark, at least overnight. The next day, the PVC cylinders containing the animals were transferred into filtered lake water for 1–3 h, and then placed in fresh lake water for 13–18 h. Afterwards, the cylinders were immersed for 2 min in soda water; the anaesthetized animals were collected under the dissecting microscope, counted, and placed on a 25 mm GF/C filter, which 307 E.Pandolfini et al. was placed in a glass scintillation vial and frozen in liquid nitrogen. Meanwhile, the feeding suspension was fractionated on a 63 µm screen in order to separate faecal pellets from the non-ingested algae. Both fractions were collected by filtration under a mild vacuum on 47 mm GF/C filters. Other details of the experimental protocol, including extraction in 90% acetone (HPLC grade), can be found elsewhere (Descy et al., 1999). Two experiments were carried out using phytoplankton of different composition, and each experiment consisted of six sets of experimental beakers with or without animals (controls), incubated under the same conditions. Daphnia galeata adults (mean length 800 µm) were available from cultures run at CRP/CU, where the cladocerans were fed with a culture of the unicellular Chlorophyte Kirchneriella subcapitata Korš. at saturating concentration. For the feeding experiments, 50 animals were selected and allowed to acclimate in an artificial algal assemblage made of river water sieved on a 63 µm Nitex screen and supplemented with a culture of Cryptomonas ozolini Skuja, in order to obtain a suspension containing Chlorophytes, diatoms and Cryptophytes. All experiments were run in 250 ml suspensions and followed a similar design as for the copepods, except that the PVC cylinders containing the cladocerans were designed in such a way that the animals remained immersed throughout the transfer steps. Also, the final feeding suspensions were not fractionated as cladocerans do not produce coherent faecal pellets. Again, acetone extracts of the animals were obtained following the procedure of Descy et al. (Descy et al., 1999). Two experiments were carried out, at low and high algal biomass, and five replicates of controls and incubations with 50 animals were run through each experiment. In addition, 50 ml from the filtered feeding suspension were eluted through Waters Sep Pak cartridges to allow detection of possible pigments in the filtrate from passage of subcellular material through the GF/C filters. As the experiments with both species were run at higher animal densities than in natural conditions, they may a priori not reflect feeding behaviour at lower densities. However, the purpose of these incubations was to test whether gut pigments were indicative of the algae ingested, and to evaluate possible pigment loss. Working at lower animal densities would have required the adoption of longer incubation times which would have led to increased risk of pigment degradation due to bacterial activity. Nevertheless, crowded conditions may have affected pigment loss, particularly in the case of cladocerans, as discussed below. Similarly, all incubations were run in the dark to avoid photochemical degradation of pigments, and we did not investigate variations of feeding activity possibly related to diel variations and photoperiod. We did, however, check for the absence of significant variation of pigment concentration in algal suspensions by analysing control suspensions at the beginning and at the end of the incubations. Given that the incubations were run in the dark and that the water was fairly rich in inorganic N and P, it was assumed that the results were not affected by algal growth from nutrient recycling in the suspensions containing the animals. 308 Pigments in freshwater zooplankton Grazing rates and pigment loss Eudiaptomus gracilis grazing rate was calculated from the difference between a given pigment concentration in the control and the final suspension (after removal of the faecal pellets), assuming that the decrease resulted from copepod feeding. Clearance rate (µl copepod–1 h–1) was calculated according to Adrian (Adrian, 1991). Pigment loss related to consumption by copepods was calculated as the difference between the amount of pigment ingested and the amount of pigment retrieved (animals + faecal pellets), expressed as a percentage of ingested pigment. Daphnia galeata clearance rates were determined by counting algae units (cells or colonies), using an inverted microscope, in three control and feeding suspensions. Most taxa were identified at the species level but clearance rates were determined by summing algal numbers by class. The proportion of marker pigment ingested was calculated from the percentage of cells of the corresponding algal class cleared by cladoceran grazing. Estimates of pigment loss were based on the same formula as for copepods, except that the amounts of pigments found in the filtrate of the feeding suspension were added to the pigments retrieved in the particulate fraction. For both animals, ingestion rates were determined by multiplying the clearance rate by the algal concentration, expressed as µg C l–1. Carbon biomass was obtained from chlorophyll a biomass using a conversion factor of 35; this value takes into account the average C:chlorophyll a ratio for the river phytoplankton used here (Descy and Gosselain, 1994) and the ratio found for the Cryptomonas culture. Chlorophyll a biomass of each algal class was estimated using the CHEMTAX program (Mackey et al., 1996). Class-specific grazing rates of copepods were determined in a similar way, using carotenoids for estimating the biomass of the dominant algae in the feeding suspensions, i.e. fucoxanthin for diatoms and Chrysophytes, lutein for Chlorophytes, zeaxanthin for Cyanophytes and alloxanthin for Cryptophytes. Further details on carotenoid distribution across algal classes, along with their amounts relative to chlorophyll a in various categories of planktonic algae, can be found elsewhere [e.g. (Mackey et al., 1996)]. HPLC analysis The HPLC system used comprised a Waters autosampler, a Waters 996 PDA detector, and a Waters 470 fluorescence detector set up for optimal detection of phaeopigments (lexc 412 nm; lem 674 nm). The separation was achieved using a 25 cm Waters Nova-Pak C18 column and the ternary gradient of Wright et al. (Wright et al., 1991), slightly modified to allow elution of xanthophylls over a larger range of retention times. The standard procedure consisted of the injection of 50–100 µl of extract run through a 30 min gradient. Calibration was achieved using three or four point calibration curves established with a mixed standard of fucoxanthin, lutein, zeaxanthin, chlorophyll a, chlorophyll b, echinenone and bcarotene, as well as a separate phaeophytin a standard. A separate standard was also used for alloxanthin. Standard solutions of fucoxanthin, alloxanthin and 309 E.Pandolfini et al. chlorophyll a were purchased from VKI, Denmark, chlorophyll b from Sigma, and lutein, zeaxanthin, echinenone and b-carotene were donated by HoffmannLaroche, Switzerland. Carotenoids not present in the mixed standard were quantified against fucoxanthin using as relative response the ratio of the specific absorbance coefficients at 440 nm in methanol. Similarly, phaeophorbides a and phaeophytins a were quantified against the phaeophytin a standard, assuming an equivalent fluorescence yield. Results Typical absorbance and fluorescence chromatograms obtained from HPLC analysis of animal samples after incubation are shown in Figure 1. Fucoxanthinol, a transformation product of fucoxanthin (Repeta and Gagosian, 1982), was found in some Daphnia extracts. As it was irregularly detected, it could not be taken into account in the fucoxanthin mass balance calculations made for estimating pigment loss. Experiments with Eudiaptomus gracilis The pigment profiles of the algal suspensions used in the two experiments (Figure 2, controls) indicate that they differed substantially, both in biomass and composition. In the first profile, Cryptophytes dominated the phytoplankton (with probably as much as 75% of the chlorophyll a biomass), and were accompanied by some Chrysophytes and cyanobacteria. In the second, Chrysophytes were dominant (with ~66% of the chlorophyll a biomass), accompanied by Cryptophytes and Chlorophytes. Clearance rates of the copepods on all algae, based on chlorophyll a removal (Figure 3A), reached 408 µl copepod–1 h–1 in the first experiment and was about four times lower in the second (92 µl copepod–1 h–1). This difference can probably be related to the contrasting biomass range; copepods of the first set of incubations started feeding at 3.6 µg chlorophyll a l–1 and reduced the algal concentration by a factor of 3. They were most likely feeding at their maximum filtration rate. By contrast, in the second experiment, the copepods were exposed to relatively high biomass throughout the incubation, possibly leading to a reduced filtration rate. Eudiaptomus gracilis selected food items very differently from among the algal groups present. In the first case (Figure 3A), they cleared mainly Cryptophytes, ingested some Chrysophytes and diatoms and, to a much lesser extent, Chlorophytes and cyanobacteria. In the second case, they Fig. 1. Typical absorbance and fluorescence chromatograms obtained from HPLC analysis of animal samples after incubation; the x-axis is retention time in minutes. (A) Absorbance chromatogram, at 436 nm, of a Daphnia galeata extract (50 animals); (B) fluorescence chromatogram of the same extract; (C) absorbance chromatogram, at 436 nm, of Eudiaptomus gracilis (70 animals); (D) fluorescence chromatogram of a faecal pellets sample of E.gracilis. Identified pigments are as follows: 1. fucoxanthinol; 2. fucoxanthin; 3. neoxanthin; 4. violaxanthin; 5. phaeophorbide a; 6. diadinoxanthin; 7. astaxanthin; 8. alloxanthin (spectrum shown in the box in C); 9. lutein; 10. zeaxanthin; 11. canthaxanthin; 12. phaeophorbide?; 13. chlorophyll b; 14. chlorophyll a; 15. echinenone; 16. phaeophytin a; 17. bb-carotene; 18. pyrophaeophytin a. 310 Pigments in freshwater zooplankton 311 E.Pandolfini et al. Fig. 2. Results of two feeding experiments with the freshwater copepod Eudiaptomus gracilis, isolated from Esch-sur-Sûre Lake, incubated in lake water. Pigment amounts (pmoles) measured in the control algal suspensions and in the different fractions (final algal suspensions, faecal pellets, animals), at the end of a feeding experiment with adult copepods. Estimates of the percentage of pigment lost indicated in the diagram. Fig. 3. Results of two feeding experiments with the freshwater copepod Eudiaptomus gracilis, isolated from Esch-sur-Sûre Lake, incubated in lake water. (A) Clearance rates calculated from pigment removal from the feeding suspensions; fucoxanthin is a marker for Chrysophytes and diatoms, alloxanthin for Cryptophytes, lutein for Chlorophytes, zeaxanthin for cyanobacteria, and chlorophyll a for total algal biomass. (B) Pigments detected in copepod extracts at the end of the incubations. (C) Pigments detected in the faecal pellets at the end of the incubations. 312 Pigments in freshwater zooplankton 313 E.Pandolfini et al. consumed exclusively Chrysophytes. In fact, calculations of ingestion rates show that they ingested the same amount of food on both occasions (~80 ng C copepod–1 h–1) by feeding on the most abundant prey. Pigments found in copepod guts (Figure 1; Figure 3B) were only partly consistent with the feeding behaviour described above. Indeed, a major peak of alloxanthin was detected in the animal extracts after both experiments, although Cryptophytes were not consumed in the second set of incubations (no decrease of alloxanthin in the final suspension; see Figure 2). By contrast, pigments in the faecal pellets (Figure 3C) seemed to reflect more closely the preferential ingestion of Cryptophytes and Chrysophytes in the first and the second experiment, respectively. However, as shown by the pigment budget and by the estimation of pigment loss, the destruction of xanthophylls was variable; 95% of the fucoxanthin ingested was lost in the second experiment, and close to 70% of lutein and zeaxanthin in the first. Surprisingly, an excess of alloxanthin was retrieved from the three fractions collected at the end of the incubations, especially in the second experiment where no Cryptophytes were consumed. The bulk of this pigment was present in the copepod extracts rather than in the faecal pellet fraction. Therefore, the presence of an animal pigment with a retention time close to alloxanthin was suspected, but analysis of the absorption spectrum showed a perfect match with that of alloxanthin (Figure 1C). Experiments with Daphnia galeata Daphnia galeata cleared the total algal biomass at similar filtration rates (Figure 4A) in both experiments (560 and 470 µl cladoceran–1 h–1, c.v. ~30% for both values), although the incubations were run at relatively low (9.2 µg chlorophyll a l–1) and high (35.8 µg chlorophyll a l–1) food levels. The low food level was actually close to the incipient limiting level known for the species [0.7 mg C l–1 (Stuchlik, 1991)] so that the clearance rate was not accelerated to any great extent. The ingestion rate was lower at low food (160 ng C cladoceran–1 h–1) compared with high food levels (765 ng C cladoceran–1 h–1). According to class-specific clearance rates estimated from the decrease in algal numbers (Figure 4A), all main groups of algae were consumed, with some variation. For instance, Chlorophytes in the first experiment, in which they comprised various small taxa mostly in the edible range for cladocerans, were preferred, while the reduced grazing rate on green algae in the second experiment may be explained by the dominance of colonies of Actinastrum, Dictyosphaerium and Scenedesmus, which were probably much less edible for Daphnia. Similarly, the higher clearance of Cryptophytes in the second experiment was probably related to the high abundance of small Chroomonas, as their concentrations were proportionately more reduced by cladoceran grazing than those of the larger Cryptomonas ozoloni. The pigments found in cladoceran guts generally reflected the composition of their diet (Figure 1; Figure 4B), i.e. the main xanthophylls of the ingested algae were detected by HPLC and were present in proportions that corresponded well with the group-specific clearance rates (Figure 4A). Phaeophorbide a was the dominant form of a phorbin in the cladocerans and its concentration also 314 Pigments in freshwater zooplankton Fig. 4. Results of two feeding experiments with the cladoceran Daphnia galeata, cultured at high food level, incubated in a mixed algal suspension. (A) Clearance rates calculated using cell counts in control and feeding suspensions. (B) Pigments detected in cladoceran extracts at the end of the incubation; fucoxanthin is a marker for Chrysophytes and diatoms, alloxanthin for Cryptophytes, lutein for Chlorophytes, zeaxanthin for cyanobacteria, and chlorophyll a for total algal biomass. increased in the feeding suspensions. Chlorophyll a concentrations decreased in those suspensions, whereas phaeophytin a concentrations were similar in controls and feeding incubations. Estimates of pigment loss rates, based on calculations of the amounts of algae ingested from the microscope counts in the control and final suspensions, showed relatively low and variable degrees of pigment disappearance (Figure 5). When diatoms and green algae were ingested, loss rates for fucoxanthin and lutein were ~30 and 18.7%, respectively. The loss rates for alloxanthin differed greatly between experiments, with 68% loss in the first experiment and no significant loss in the second. Ingestion rates for Cryptophytes, however, were higher in the first experiment. The estimates of losses, in the case of cladocerans, were calculated taking 315 E.Pandolfini et al. into account pigment retrieved from the ‘soluble’ fraction, which represented between 1% (fucoxanthin) and 2.5% (alloxanthin) of the total pigment at the end of the incubations. There was some variability associated with pigment loss estimates as they were estimated on the basis of cell counts in control and final suspensions, which have a limited accuracy (coefficient of variation 20–30%). In addition, it is worth emphasizing that total a phorbins, after grazing by D.galeata, did not change significantly, indicating that chlorophyll a was converted to phaeophorbide a with 100% efficiency, without significant destruction. Fig. 5. Results of two feeding experiments with the cladoceran Daphnia galeata, cultured at high food level, incubated in an artificial algal suspension. Pigment amounts (pmoles) measured in the control and final algal suspensions, at the end of a feeding experiment with adult cladocerans, and amount of ingested pigment estimated from algae removal. Estimates of the percentage of pigment lost indicated in the diagram. 316 Pigments in freshwater zooplankton Discussion The main conclusion of this experimental study is that pigment processing during gut passage appears to be different in Eudiaptomus gracilis and Daphnia galeata, and that it may vary for the same pigment in different species of ingested algae. Recent research on the fate of chlorophyll a and carotenoids points to substantial pigment destruction during feeding by marine and freshwater calanoid copepods, although literature specific to xanthophylls is more sparse (Head and Harris, 1992, 1994; Kleppel, 1998; Descy et al., 1999). These papers have reported extensive destruction of fucoxanthin during consumption of diatoms, which may vary with season and food concentration (Kleppel, 1998), with 0 recovery after copepod grazing at low food levels. For chlorophyll a, most studies now agree on variable degrees of transformation to products undetectable by HPLC and gut fluorescence (Drits, 1997; Stevens and Head, 1998; Tirelli and Mayzaud, 1998). These findings, together with possible variations in gut evacuation rates, are serious restrictions to quantitative estimates of grazing rates by the gut fluorescence technique. Chlorophyll and carotenoid destruction also occurs as a result of microzooplankton grazing (Strom et al., 1998) and seems to be inversely related to protozoan size. The results reported here for the freshwater copepod E.gracilis, and those for another species, Diaptomus minutus (Descy et al., 1999) point to similar and extensive pigment alterations as a result of feeding. Therefore, pigments in animal guts are not a reliable indication of copepod diet and should not be used to assess grazing quantitatively. An additional problem reported here is the presence of alloxanthin in animals, even when they have not fed on Cryptophytes. A similar intriguing observation was made with D.minutus starved for 8 h in filtered water, and on copepods which had not fed on Cryptophytes (Descy et al., 1999). The most likely explanation is that the pigment systematically detected in these copepods is an alloxanthin-like animal pigment, which perhaps has a photo-protective role. It should be noted that in another study (Descy et al., 1999), D.minutus contained large amounts of the animal carotenoid astaxanthin, which, in this study, was observed only at low concentrations in E.gracilis. Our results with D.galeata are consistent with those reported in two other studies, one by Quiblier et al. (Quiblier et al., 1994) who observed relatively high levels of carotenoid conservation in D.magna along with chlorophyll a degradation, and one by Fundel et al. (Fundel et al., 1998) who reported conversion of chlorophyll a to phaeophorbide a except at high grazer density, where a phorbin destruction occurred. This indicates that with regard to other grazers, food concentration may have an effect on pigment conservation in freshwater cladocerans. In our experiments, a phorbins were not destroyed significantly, and phaeophorbides a present in the animals increased in concentration in feeding suspensions. It is worth mentioning, however, that the Daphnia used in these incubations came from laboratory cultures, where they had been fed at high algal concentrations, and that the incubations also were not run at very low food levels. A similar chlorophyll a transformation pattern, with dominance of phaeophorbide a among the degradation products, was found in another freshwater cladoceran, Bosmina longirostris 317 E.Pandolfini et al. (Descy, unpublished). Another factor which may have affected the fate of pigments in our experiments is the experimental design itself. The cladocerans, which were at higher density than under natural conditions, may have re-ingested algae during the incubation as faeces accumulated in the feeding suspensions (whereas faecal pellets were settled out through the Nitex screen in the copepod experiments). Pigment re-ingestion was similarly identified as a potential cause of further pigment degradation in experiments with microzooplankton (Strom et al., 1998). To summarize, whereas algal carotenoids found in the freshwater copepod E.gracilis were not reliable for assessing their diet, those found in the cladoceran D.galeata reflected qualitatively the type of algae ingested. In both species, pigment amounts in the guts were not directly related to the group-specific clearance rates and therefore should not be used for estimating quantitatively the class-specific ingestion rates. Moderate to extensive destruction of these pigments occurred, as shown by our calculations of loss rates and by the presence of fucoxanthinol in some Daphnia extracts. Therefore, caution is recommended in using gut pigments as indicators of cladoceran diet. Acknowledgements We are grateful to David Courtois for his help in the experiments with Daphnia galeata. Part of this study was carried out with financial support of the project MODEL of the CRP-CU. E.P. benefited from a research fellowship awarded by the university La Sapienza, Roma, Italy. References Adrian,R. (1991) Filtering and feeding rates of cyclopoids feeding on phytoplankton. Hydrobiologia, 210, 217–223. Bautista,B. and Harris,R.P. (1992) Copepod gut contents, ingestion rates and grazing impact on phytoplankton in relation to size structure of zooplankton and phytoplankton during a spring bloom. Mar. Ecol. Progr. Ser., 82, 41–50. Bochdansky,A.B., Deibel,D. and Hatfield,E.A. (1998) Chlorophyll a conversion and gut passage time for the pelagic tunicate Oikopleura vanhoeffenii (Appendicularia). J. Plankton Res., 20, 2179–2197. Conover,R.J., Durvasula,R., Roy,S. and Wang,R. (1986) Probable loss of chlorophyll-derived pigments during passage through the gut of zooplankton, and some of the consequences. Limnol. Oceanogr., 31, 878–887. Descy,J.-P. and Gosselain,V. (1994) Development and ecological importance of phytoplankton in a large lowland river (River Meuse, Belgium). Hydrobiologia, 289, 139–155. Descy,J.-P., Frost,T.M. and Hurley,J.P. (1999) Assessment of grazing by the freshwater copepod Diaptomus minutus using carotenoid pigments: a caution. J. Plankton Res., 21, 127–146. Dohet,A. and Hoffmann,L. (1995) Seasonal succession and spatial distribution of the zooplankton community in the reservoir of Esch-sur-Sûre (Luxembourg). Belg. J. Zool., 125, 109–123. Drits,A.V. (1997) On the possibility of the application of gut fluorescence method for studies of herbivorous zooplankton feeding. Okeanlogiya, 37, 753–760. Fundel,B., Stich,H.-B., Schmid,H. and Maier,G. (1998) Can phaeopigments be used as markers for Daphnia grazing in Lake Constance? J. Plankton Res., 20, 1449–1462. Head,E.J.H. and Harris,L.R. (1992) Chlorophyll and carotenoid transformation and destruction by Calanus spp. grazing on diatoms. Mar. Ecol. Prog. Ser., 86, 229–238. Head,E.J.H. and Harris,L.R. (1994) Feeding selectivity by copepod grazing on natural mixtures of phytoplankton determined by HPLC analysis of pigments. Mar. Ecol. Prog. Ser., 110, 74–83. Head,E.J.H. and Harris,L.R. (1996) Chlorophyll destruction by Calanus spp. grazing on 318 Pigments in freshwater zooplankton phytoplankton: kinetics, effects of ingestion rate and feeding history, and a mechanistic interpretation. Mar. Ecol. Prog. Ser., 135, 223–235. Kleppel,G.S. (1998) The fate of the carotenoid pigment fucoxanthin during passage through the copepod gut: pigment recovery as a function of copepod species, season and food concentration. J. Plankton Res., 20, 2017–2028. Kleppel,G.S. and Pieper,R.E. (1984) Phytoplankton pigments in the gut contents of planktonic copepods from coastal waters off southern California. Mar. Biol., 78, 193–198. Kleppel,G.S., Frazel,D., Pieper,R.E. and Holliday,D.V. (1988) Natural diets of zooplankton off southern California. Mar. Ecol. Prog. Ser., 49, 231–241. Kleppel,G.S., Holliday,D.V. and Pieper,R.E. (1991) Trophic interactions between copepods and microplankton: a question about the role of diatoms. Limnol. Oceanogr., 36, 172–178. Landry,M.R., Peterson,W.K. and Fagerness,V.L. (1994) Mesozooplankton grazing in the Southern California Bight. I. Population abundances and gut pigment contents. Mar. Ecol. Prog. Ser., 115, 55–71. Lopez,M.D.G., Huntley,M.E. and Sykes,P.F. (1988) Pigment destruction by Calanus pacificus: impact on the estimation of water column fluxes. J. Plankton Res., 10, 715–734. Mackas,D. and Bohrer,R. (1976) Fluorescence analysis of phytoplankton gut contents and an investigation of diel feeding patterns. J. Exp. Mar. Biol. Ecol., 25, 77–85. Mackey,M., Mackey,D.J., Higgins,H.W. and Wright,S.W. (1996) CHEMTAX—a program for estimating class abundances from chemical markers: application to HPLC measurements of phytoplankton. Mar. Ecol. Progr. Ser., 144, 265–283. Mayzaud,P. and Razouls,S. (1992) Degradation of gut pigment during feeding by a subantarctic copepod: importance of feeding history and digestive acclimation. Limnol. Oceanogr., 37, 393–404. Millie,D.F., Paerl,H.W. and Hurley,J.P. (1993) Microalgal pigment assessment using high performance liquid chromatography: a synopsis of organismal and ecological applications. Can. J. Fish. Aquat. Sci., 50, 2513–2527. Nelson,J.R. (1989) Phytoplankton pigments in macrozooplankton feces: variability in carotenoids alterations. Mar. Ecol. Prog. Ser., 52, 129–144. Penry,D.L. and Frost,B.W. (1991) Chlorophyll a degradation by Calanus pacificus: Dependence on ingestion rate and digestive acclimation to food resources. Limnol. Oceanogr., 36, 147–159. Quiblier,C., Bourdier,G., Amblard,C. and Pépin,D. (1994) Separation of phytoplanktonic pigments by HPLC for the study of phyto-zooplankton trophic relationships. Aquat. Sci., 56, 29–39. Quiblier-Lloberas,C., Bourdier,G., Amblard,C. and Pépin,D. (1996) A qualitative study of zooplankton grazing in an oligo-mesotrophic lake using phytoplanktonic pigments as organic markers. Limnol. Oceanogr., 41, 1767–1779. Stevens,C.J. and Head,E.J.H. (1998) A model of chlorophyll a destruction by Calanus spp. and implications for the estimation of ingestion rates using the gut fluorescence method. Mar. Ecol. Prog. Ser., 171, 187–198. Repeta,D.J. and Gagosian,R.B. (1982) Carotenoid transformations in coastal marine waters. Nature, 295, 51–54. Strom,S.L., Morello,T.A. and Bright,K.J. (1998) Protozoan size influences algal pigment degradation during grazing. Mar. Ecol. Prog. Ser., 164, 189–197. Stuchlik,E. (1991) Feeding behaviour and morphology of filtering combs of Daphnia galeata. Hydrobiologia, 225, 155–156. Tirelli,V. and Mayzaud,P. (1998) Gut pigment destruction by the copepod Acartia clausi. J. Plankton Res., 20, 1953–1962. Wright,S.W., Jeffrey,S.W., Mantoura,R.F.C., Llewellyn,C.A., Bjornland,T., Repeta,D. and Welschmeyer,N. (1991) Improved HPLC method for the analysis of chlorophylls and carotenoids in marine phytoplankton. Mar. Ecol. Progr. Ser., 77, 183–196. Received on March 31, 1999; accepted on August 25, 1999 319