Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Immune system wikipedia , lookup
Polyclonal B cell response wikipedia , lookup
Molecular mimicry wikipedia , lookup
Lymphopoiesis wikipedia , lookup
Psychoneuroimmunology wikipedia , lookup
Adaptive immune system wikipedia , lookup
Cancer immunotherapy wikipedia , lookup
Adoptive cell transfer wikipedia , lookup
This information is current as
of June 18, 2017.
Identification of an Important Immunological
Difference between Virulent Varicella-Zoster
Virus and Its Avirulent Vaccine: Viral
Disruption of Dendritic Cell Instruction
Cindy Gutzeit, Martin J. Raftery, Matthias Peiser, Karsten B.
Tischer, Martina Ulrich, Melanie Eberhardt, Eggert
Stockfleth, Thomas Giese, Andreas Sauerbrei, Craig T.
Morita and Günther Schönrich
References
Subscription
Permissions
Email Alerts
This article cites 54 articles, 29 of which you can access for free at:
http://www.jimmunol.org/content/185/1/488.full#ref-list-1
Information about subscribing to The Journal of Immunology is online at:
http://jimmunol.org/subscription
Submit copyright permission requests at:
http://www.aai.org/About/Publications/JI/copyright.html
Receive free email-alerts when new articles cite this article. Sign up at:
http://jimmunol.org/alerts
The Journal of Immunology is published twice each month by
The American Association of Immunologists, Inc.,
1451 Rockville Pike, Suite 650, Rockville, MD 20852
Copyright © 2010 by The American Association of
Immunologists, Inc. All rights reserved.
Print ISSN: 0022-1767 Online ISSN: 1550-6606.
Downloaded from http://www.jimmunol.org/ by guest on June 18, 2017
J Immunol 2010; 185:488-497; Prepublished online 4 June
2010;
doi: 10.4049/jimmunol.0902817
http://www.jimmunol.org/content/185/1/488
The Journal of Immunology
Identification of an Important Immunological Difference
between Virulent Varicella-Zoster Virus and Its Avirulent
Vaccine: Viral Disruption of Dendritic Cell Instruction
Cindy Gutzeit,*,1 Martin J. Raftery,* Matthias Peiser,† Karsten B. Tischer,‡
Martina Ulrich,x Melanie Eberhardt,* Eggert Stockfleth,x Thomas Giese,{
Andreas Sauerbrei,‖ Craig T. Morita,# and Günther Schönrich*
V
aricella-zoster virus (VZV), a member of the human herpesvirus family, infects the majority of the human population. Similar to most other members of the human
herpesvirus family, VZV has a narrow host range and does not infect mice or cells of murine origin (1). In humans, VZV is transmitted by aerosols and initially infects the upper respiratory tract.
During the incubation period, VZV spreads to regional lymph
nodes to infect T cells that subsequently transport virions to cutaneous sites (2). There, it efficiently replicates in epithelial cells
*Institute of Virology, †Institute of Molecular Biology and Bioinformatics, and xDepartment of Dermatology, Venerology and Allergology, Charité–Universitätsmedizin Berlin;
‡
Department of Veterinary Medicine, Institute of Virology, Freie Universität Berlin, Berlin;
{
Institute of Immunology, University of Heidelberg, Heidelberg; ‖Institute of Virology and
Antiviral Therapy, Friedrich-Schiller University of Jena, Jena, Germany; and #Division of
Rheumatology, Department of Internal Medicine and the Interdisciplinary Graduate Program in Immunology, University of Iowa College of Medicine, Iowa City, IA 52242
1
Current address: Clinical Allergy Research Unit, Department of Medicine Solna,
Karolinska Institutet, Karolinska University Hospital Solna, Stockholm, Sweden.
Received for publication August 26, 2009. Accepted for publication April 21, 2010.
This work was supported by grants from the Deutsche Forschungsgemeinschaft
(SFB421 and SCHO 592/6-1, to G.S.), the Universitäre Forschungsförderung of
the Charité – Universitätsmedizin Berlin (to C.G.), and the National Institutes of
Health (AR45504, AI057160 [Midwest Regional Center of Excellence for Biodefense and Emerging Infectious Diseases Research], and CA113874, to C.T.M.).
Address correspondence and reprint requests to Dr. Günther Schönrich, Institute of
Virology, Charité–Universitätsmedizin Berlin, Charitéplatz 1, 10117 Berlin, Germany.
E-mail address: [email protected]
Abbreviations used in this paper: DC, dendritic cell; gE, glycoprotein E; LC, Langerhans cell; LTA, lipoteichoic acid; MDDC, monocyte-derived dendritic cell; PI,
propidium iodide; poly(I:C), polyinosinic:polycytidylic acid; PRR, pattern recognition receptor; rh, recombinant human; V-Oka, vaccine Oka; Vd1+ T cells, gd T cells
expressing variable d1 chain; VZV, varicella-zoster virus.
Copyright Ó 2010 by The American Association of Immunologists, Inc. 0022-1767/10/$16.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.0902817
and fibroblasts, causing the typical varicella rash. After primary
infection, VZV establishes latency within sensory ganglia. From
there, it can reactivate to cause herpes zoster many years later.
This disease is characterized by papulovesicular skin lesions,
mostly restricted to a single dermatome, and is frequently followed by severe pain. Cellular and humoral immune responses
are essential for the control of VZV replication (3). VZV is the
first human herpesvirus for which a live attenuated vaccine (vaccine Oka [V-Oka]) has been generated and approved (4). It is used
successfully to immunize millions of children annually in the
United States, Europe, and Japan and is now considered a prophylactic option in older individuals to prevent herpes zoster (5).
Despite its unquestionable medical importance, the immunological difference between the vaccine and wild type virulent VZV has
not been defined.
Virulent VZV efficiently spreads in human skin, whereas
avirulent V-Oka only rarely causes clinical reactions. Thus, similar
to other members of the human herpesvirus family, virulent VZV
has evolved intricate immune-evasion strategies (6). These capabilities have been lost during the attenuation process generating the
avirulent VZV vaccine. It should be possible to define virulenceassociated evasion mechanisms by comparatively studying the capacity of virulent and avirulent VZV to subvert immune reactions.
Dendritic cells (DCs) represent a prime target for immunoevasive
viruses, because they couple innate to adaptive immunity to initiate
efficient antiviral T cell responses. In the steady-state, two major DC
subsets can be found in the skin (7): epidermal Langerhans cells
(LCs) and dermal DCs. They form a network of sentinels that sense
invading pathogens. Intriguingly, another DC subset derived from
immigrating monocytes accumulates under inflammatory conditions.
These inflammatory DCs replace steady-state DCs that disappear as
Downloaded from http://www.jimmunol.org/ by guest on June 18, 2017
Virulent varicella-zoster virus (VZV) can spread in immunocompetent humans, resulting in symptoms mostly of the skin. In
contrast, vaccine Oka (V-Oka), the attenuated VZV vaccine strain, only rarely causes clinical reactions. The mechanisms underlying these pathogenetic differences are unclear. In this study, we comparatively analyzed the ability of virulent VZV and
V-Oka to modulate instruction of dendritic cells (DCs) by innate signals. DCs isolated from normal human skin were susceptible to
infection with VZV and V-Oka. Moreover, inflammatory DCs, which play a crucial role in the stimulation of Th1 immune
responses, accumulated in herpes zoster lesions. Infection of inflammatory DCs generated in vitro with virulent VZV or V-Oka
resulted in upregulation of CD1c. Upon coculture with CD1c-restricted innate cells, DCs developed a mature phenotype whether
infected with virulent VZV or V-Oka. Intriguingly, a striking difference was detected on the functional level. The release of IFN-g
and IL-12, the signature cytokines of Th1 responses, was enhanced by V-Oka but blocked by virulent VZV. V-Oka and virulent
VZV efficiently synergized with CD40L, eliminating the possibility that CD40 signaling was a target of VZV-associated immune
evasion. Instead, virulent VZV selectively interfered with signaling through TLR2, which is known to sense VZV. Thus, virulent
VZV subverts Th1-promoting instruction of human DCs by blocking TLR2-mediated innate signals that prime IL-12 production
by DCs. Taken together, our results demonstrate a novel immune-evasion mechanism of virulent VZV that has been lost during
the attenuation process leading to the VZV vaccine strain. The Journal of Immunology, 2010, 185: 488–497.
The Journal of Immunology
489
VZV-infected fibroblast monolayers with a density of 1 3 106 cells/ml in
media containing recombinant human (rh)TGF-b (10 ng/ml), rhGM-CSF
(500 IU/ml), and rhIL-4 (200 IU/ml).
a result of migration to the lymph nodes or apoptosis (8, 9). Virulent
VZV is known to infect monocyte-derived DCs (MDDCs) generated
in vitro (10–12). However, no functional defect of immature MDDCs
after VZV infection has been defined (10, 11).
Powerful Th1-like adaptive immune responses are essential to
achieve virus control. To promote Th1 responses, DCs must provide three signals: signal 1 is delivered by Ag presentation through
specialized surface molecules; signal 2 is mediated by costimulatory molecules on the surface, such as CD40, CD80, and CD86; and
signal 3 is delivered by release of IL-12. Immature DCs receive
a combination of maturation stimuli from different sources, which
they integrate and translate into different qualities of adaptive T cell
responses. Innate lymphocytes, which are already active before the
onset of adaptive T cell responses, are crucial for DC maturation.
For example, CD1c-restricted gd T cells can drive DC differentiation toward a Th1-polarizing program (13). This bidirectional
interaction between innate lymphocytes and DCs, called DC instruction, is mediated by cytokines, such as IFN-g, and by direct
cell-to-cell interaction through ligands of the TNF family, most
notably CD40L (14, 15). Importantly, optimal IL-12 production,
the signature cytokine of Th1 responses, requires conditioning of
DCs by pathogen-associated danger signals that synergistically act
through multiple pattern recognition receptors (PRRs) (16–19). To
understand viral pathogenesis and develop effective viral vaccines,
it is of the utmost importance to unravel the intricate interactions
among DCs, innate lymphocytes, and pathogen-associated danger
signals.
In this study, we analyzed the modulation of DC instruction by
the VZV vaccine compared with virulent VZV clinical isolates. We
show that the VZV vaccine primes human MDDCs for IL-12 production. In contrast, virulent VZV disrupts the programming of
MDDCs toward a Th1-polarizing function. As an underlying mechanism, we identified a block in PRR signaling postinfection of
MDDCs with virulent VZV, a fundamental immunological difference between the vaccine and clinical isolates. In conclusion, we
define a novel virulence-associated immune-evasion strategy that is
important for understanding VZV-associated pathogenesis and has
to be considered in future attempts to develop novel vaccines.
Anti-CD1a (HI149) was purchased from Immunotools; anti-CD1b (4.A7.6)
and anti-CD1c (L161) were obtained from Immunotech (Hamburg,
Germany); anti-CD40 (5C3), anti-CD83 (HB15e), anti-CD86 (IT2.2), antiTCR-g/d-1 (11F2), anti-IgG1 (MOPC-21), and anti-IgG2b (MPC-11)
were purchased from BD Pharmingen (Heidelberg, Germany); and antiglycoprotein E (gE) (MAB8612) was obtained from Chemicon International
(Hofheim, Germany). As secondary Abs, Alexa Fluor 488-labeled anti-IgG2b
and Alexa Fluor 568-labeled anti-IgG1 from Invitrogen (Karlsruhe, Germany)
were used; FITC-, R-PE–, allophycocyanin-, and Cy5-labeled isotype-specific
Abs from Jackson ImmunoResearch Laboratories (West Grove, PA) were
used. For neutralization assays, mouse IgG1 (MOPC-21) and mouse antiCD1c (L161) were added at 20 mg/ml, and polyclonal anti–IL-12 (R&D
Systems, Wiesbaden, Germany) was added at 5 mg/ml. Stimulation with
LPS was performed at 1 mg/ml and with PHA at 100 ng/ml. Apoptotic death
was visualized by using FITC-labeled Annexin V and propidium iodide (PI;
Santa Cruz Biotechnology, Santa Cruz, CA).
Materials and Methods
T cell assays
Skin punch biopsies (4 mm) were taken from control subjects undergoing
breast reduction and from herpes zoster patients (vesicular stage, n = 2).
Patients with herpes zoster were immunocompetent (mean age, 62 6 10.5 y).
Skin biopsies were embedded in Jung tissue freezing medium (Leica
Instruments, Göttingen, Germany) and cryo-frozen. Five-micrometer sections were fixed for 10 min in acetone (220˚C), air-dried overnight, and
stored at 280˚C. Written informed consent for skin punch biopsies was
obtained in accordance with ethical approval EA1/169/06 of the Charité–
Universitätsmedizin Berlin.
Isolation and VZV infection of epidermal LCs and dermal DCs
Human skin was obtained from healthy volunteers undergoing plastic
surgery and prepared as recently described (20). Epidermal sheets were
detached from the dermis after overnight incubation with dispase I (2 U/ml;
Roche Applied Science, Mannheim, Germany) at 4˚C. After incubation
with trypsin solution containing 0.25% trypsin in PBS with 5 mM Mg2+ and
10 mg/ml DNase (Roche Applied Science) for 15 min at 37˚C, LCs were
positively selected from single-cell suspensions using MACS and human
CD1c (BDCA-1)+ Dendritic Cell Isolation Kit (Miltenyi Biotec, Bergisch
Gladbach, Germany). The dermal pulp was incubated for 1.5 h at 37˚C in
PBS solution containing 10% FCS, penicillin/streptomycin (10,000 U/ml),
amphotericin B (2.5 mg/ml), 5 mM MgSO4, 12 mg collagenase (CellSystems Biotechnologie Vertrieb, St. Katharinen, Germany), and 6.5 g
hyaluronidase type I S (Sigma-Aldrich, Deisendorf, Germany). The filtered suspension was used to isolate dermal DCs with the human CD1c
(BDCA-1)+ Dendritic Cell Isolation Kit. LCs and dermal DCs were
centrifuged at 160 3 g for 45 min at room temperature on mock- or
VZV was propagated on human embryonic lung fibroblasts. VZV clinical
isolates designated JoSt (genotype E1), M1_935/05 (genotype M1),
E2_769/05 (genotype E2), and E1_667/05 (genotype E1) were isolated from
varicella patients. All VZV strains used in this report were analyzed
and genotyped by A. Sauerbrei, as previously reported (21). The vaccine
strain V-Oka was grown from the live attenuated vaccine Varilrix (GlaxoSmithKline, Research Triangle Park, NC). The titers of VZV stocks
(VZV-infected cells) were determined by calculation of the tissue culture
infectious dose 50% using the Reed–Muench formula. CD40L-expressing
fibroblasts were kindly provided by R.A. Kroczek (Robert Koch Institute,
Berlin, Germany).
Generation and VZV infection of MDDCs
Buffy coats were obtained from the Deutsche Rote Kreuz (Dresden,
Germany). Monocytes were isolated through negative selection by the
MACS technique (Miltenyi Biotec) and differentiated within 3 d into immature MDDCs by culture with rhGM-CSF (500 IU/ml) and rhIL-4 (200 IU/
ml) (Immunotools, Friesoythe, Germany), as described previously (22). For
VZV infection, immature DCs were centrifuged at 150 3 g for 45 min at
room temperature on a VZV-infected fibroblast monolayer. After 24 h,
DCs were removed, and assays were performed 2 d after VZV infection.
Mock DCs were set up as described above using uninfected fibroblasts.
Abs and reagents
The well-characterized CD1c-restricted gd T cell clone (JR.2.28) was
cultured by periodic stimulation with PHA-P in the presence of irradiated
EBV-transformed B cells and PBMCs, as described (23). For cytokine
secretion, mock- or VZV-infected immature MDDCs were cocultured for
48 h with gd T cells at an E/T ratio of 1:1 (1 3 105 cells/well). Supernatants were collected and stored at 220˚C.
Measurement of cytokine production
Cytokine concentration of supernatants was measured by ELISA. ELISA
kits for human IL-12p70 (eBioscience, Vienna, Austria) and IFN-g
(Immunotools) were used.
Immunohistochemistry
Skin sections (5 mm) were blocked with 5% goat serum (host species of the
secondary Ab) for 1 h at room temperature in a humidified chamber and
then were incubated with the specific primary Abs for 1 h at 37˚C. Specific
Ag was detected with Alexa Fluor 488-labeled anti-IgG2b and Alexa Fluor
568-labeled anti-IgG1 as secondary Abs (1:400) by incubating for 45 min
at room temperature. Nuclei were visualized with the DNA-intercalating
dye DAPI (1:20.000). Stained slides were analyzed using a BX60F-3 microscope with Cell D software (Olympus, Hamburg, Germany).
Flow cytometry
Cells were washed once in ice-cold FACS-wash solution (PBS with 1% FCS
and 0.02% sodium azide) and incubated in ice-cold FACS-block solution
(PBS with 10% FCS and 0.2% sodium azide) with primary Abs at 4˚C
for 1 h. Cells were washed in ice-cold FACS-wash solution and stained
Downloaded from http://www.jimmunol.org/ by guest on June 18, 2017
Human samples
Viruses
490
with secondary Abs for 45 min at 4˚C and finally fixed with 0.4% formaldehyde. For detection of apoptotic death, cells were washed in apoptosis
buffer (10 mM HEPES [pH 7.4], 0.14 M NaCl, 5 mM CaCl2, 5% FCS,
0.02% sodium azide) and stained for 20 min at 4˚C with Annexin V. PI was
added shortly before measuring. A FACSCalibur (BD Biosciences, Heidelberg, Germany) was used for flow cytometry, and data were analyzed with
CellQuest Pro (BD Bioscience), as described recently (22).
Statistical analysis
SPSS (Chicago, IL) and GraphPad Prism (GraphPad, San Diego, CA)
software programs were used to perform statistics. The Wilcoxon test was
used to determine the significance of differences between groups; p values
,0.05 were considered significant (two-tailed). The actual p values are
indicated in the box-and-whisker plots.
Results
Virulent VZV and avirulent V-Oka infect skin DC subsets with
similar efficiency
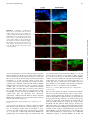
FIGURE 1. VZV infection of DC subsets
isolated from normal human skin. LCs and dermal DCs were freshly isolated from the epidermis and dermis, respectively, by magnetic bead
separation. A, LCs were stained for langerin and
CD11c (left panel shows isotype control). C,
Dermal DCs were stained for CD1c and CD11c
(left panel shows isotype control). LCs (B) and
dermal DCs (D) were cocultured with mock or
V-Oka– or JoSt-infected fibroblasts, and the surface expression of viral gE was measured 2 d
postinfection. Percentages of gE+ cells are indicated. One representative experiment of two is
shown. Additionally, in vitro-generated MDDCs
were analyzed 2 d after mock, V-Oka, or JoSt
infection. Clustering (E) and gE surface expression (F) are shown. Phase contrast microscopy
images (E) were taken with Axiovert 25 microscope (Carl Zeiss, Jena, Germany) (magnification 310). Data are representative of seven
experiments.
Taken together, these findings demonstrate that virulent VZV and
V-Oka do not differ with regard to their capacity to infect skin DC
subsets that are found during steady state or under inflammatory
conditions.
Inflammatory DCs accumulate in herpes zoster lesions
Inflammatory DCs play an important role in the development of
Th1-polarized T cells that produce IFN-g and protect against
viruses (24). Therefore, we determined whether inflammatory
DCs occur during VZV-induced inflammation. For this purpose,
we studied skin sections from papulovesicular lesions of herpes
zoster patients. Staining with DAPI (Fig. 2A) or with Papanicolaou’s solution and eosin (data not shown) revealed a strong cellular infiltration. Using immunofluorescence microscopy, we localized myeloid CD11c-expressing DCs and assessed their CD1
expression while visualizing VZV infection by staining for viral
gE. CD11c+ DCs strongly infiltrated VZV-infected skin lesions
(Fig. 2B). In contrast, CD1a+ LCs disappeared from herpes zoster
lesions and were found only in the epidermis of the control skin
(Fig. 2C). The infiltrating DCs also expressed CD1b (Fig. 2D). DCs
with detectable CD1c expression were mainly detected in close
proximity to VZV-containing vesicles in inflamed skin tissue, but
they were evenly distributed in the upper dermis of control skin
sections (Fig. 2E). Probably because papulovesicular lesions represent an advanced stage of VZV infection in the dermis, viral gE
was detectable only in a few CD1c+ cells (Fig. 2E). Altogether,
these findings suggest that steady-state DCs disappear during viral
spread, whereas inflammatory DCs strongly accumulate in herpes
zoster lesions.
MDDCs infected with virulent VZV fail to stimulate IFN-g
secretion by innate cells
Next, we comparatively studied the impact of virulent VZV and
V-Oka on phenotype and function of MDDCs in vitro. Because herpesviruses, including HSV (25, 26) and human CMV (22), can
block CD1 expression on MDDCs, we investigated the effect of
VZV on group 1 CD1 molecules. Although the expression of
CD1a (Fig. 3A, 3B) and CD1b (Fig. 3A) remained unaltered,
VZV infection with either strain significantly increased CD1c expression on immature MDDCs (Fig. 3A, 3B). This was not due to
Downloaded from http://www.jimmunol.org/ by guest on June 18, 2017
First we assessed whether DC subsets found in human skin, the
major replication site for VZV, show differential susceptibility to
infection with virulent VZV and the avirulent VZV vaccine strain.
For this purpose, LCs and dermal DCs were isolated from normal
human skin. Surface staining for myeloid marker CD11c and
langerin (LCs, Fig. 1A) or CD11c and CD1c (dermal DCs, Fig.
1C) confirmed the purity of the preparations. Freshly isolated LCs
and dermal DCs were infected with VZV, and the surface expression of viral gE was determined. After mock infection, LCs
and dermal DCs did not express gE. In contrast, 14% and 20%,
respectively, of VZV-infected LCs (Fig. 1B) and 12% and 17%,
respectively, of dermal DCs (Fig. 1D) strongly expressed gE after
VZV infection. No differences in the efficiency of infection could
be observed between the avirulent vaccine strain V-Oka and JoSt,
a virulent VZV strain. Neither LCs nor dermal DCs upregulated
costimulatory molecules postinfection (data not shown). Moreover, in vitro-generated MDDCs, which model DCs accumulating
under inflammatory conditions, were susceptible to VZV infection, as previously described (10). V-Oka and JoSt induced similar
DC clustering (Fig. 1E) and infected immature MDDCs with the
same efficiency (Fig. 1F). The majority of cells (73%) expressed
viral gE at low density, whereas high amounts of this molecule
were found on a minor population (18% and 21%, respectively).
VIRAL DISRUPTION OF DENDRITIC CELL INSTRUCTION
The Journal of Immunology
491
an increased abundance of CD1c-encoding transcripts, suggesting
that the underlying mechanism was operating on the protein level
(data not shown). We used the CD1c-restricted Vd1+ T cell clone
JR.2.28 (23) to study the functional impact of VZV-induced CD1c
upregulation. gd T cells expressing variable d1 chain (Vd1+
T cells) represent the major gd lymphocyte population in human
epithelial tissue. Strikingly, V-Oka–infected immature MDDCs
stimulated CD1c-restricted Vd1+ T cells to secrete large amounts
of IFN-g (Fig. 3C, 3D). In contrast, JoSt-infected immature
MDDCs failed to stimulate IFN-g secretion by CD1c-restricted
Vd1+ T cells. Abs to CD1c, but not a respective isotype control,
blocked IFN-g secretion, showing that stimulation of gd T cells
required CD1c (Fig. 3E). Altogether, these observations
demonstrated a striking functional difference between immature
MDDCs infected with the vaccine strain V-Oka and those infected
with the VZV clinical isolate JoSt.
Virulent VZV and the avirulent VZV vaccine strain infect gd
T cells
In the previous experiments, we could not exclude that virulent
VZV directly targets the function of innate lymphocytes. Therefore, we investigated whether VZV is transmitted to CD1crestricted gd T cells. For this purpose, the expression of viral gE
on gd T cells after coculture with VZV-infected MDDCs was
quantified (Fig. 4A). Avirulent V-Oka and clinical isolate JoSt
were transmitted from infected immature MDDCs to gd T cells
with the same low efficiency (6–7%). However, IFN-g secretion
by gd T cells in the presence of JoSt-infected immature MDDCs
could be restored by stimulation with PHA, demonstrating that
virulent VZV does not destroy the functional integrity of gd
T cells (Fig. 4B). In conclusion, the block of IFN-g release by
innate lymphocytes after stimulation with JoSt-infected immature
MDDCs was due to a virus-induced defect in MDDCs.
Interference of virulent VZV with DC function is independent
of its genotype
We next tested whether the failure of JoSt-infected immature
DCs to activate CD1c-restricted innate cells is a general characteristic of virulent VZV strains. To this end, we studied the currently circulating European VZV strains that are grouped into the
main genotypes M1, E1, and E2 (21). According to phylogenetic
analysis, the isolate M1_935/05 belonged to genotype M1, the
isolate E2_769/05 belonged to genotype E2, and the isolates
E1_667/05 and JoSt clustered within genotype E1 (Fig. 5A).
These VZV strains grew equally well in human embryonic lung
fibroblasts (data not shown). Intriguingly, all VZV clinical isolates,
regardless of their genotype, strongly blocked the capacity of immature MDDCs to activate CD1c-restricted gd T cells (Fig. 5B).
These functional differences could have been due to differential
VZV-induced apoptosis of immature MDDCs. To exclude this
Downloaded from http://www.jimmunol.org/ by guest on June 18, 2017
FIGURE 2. Accumulation of inflammatory
DCs in herpes zoster lesions. Skin sections of
healthy control or herpes zoster patients were
stained for DNA with the DNA-intercalating dye
DAPI (blue) (A), for viral gE (green) (B–E), for
CD11c (red) as a myeloid marker (B), for CD1a
(red) as an LC marker (C), and for CD1b (red)
(D) and CD1c (red) (E) as dermal DC markers.
The enlarged area in E (right panel) shows a few
cells (yellow) expressing CD1c and viral gE.
Scale bar, 200 mm. One representative biopsy
of two is depicted.
492
VIRAL DISRUPTION OF DENDRITIC CELL INSTRUCTION
FIGURE 3. Stimulation of innate cells by VZV-infected DCs. Immature
MDDCs were analyzed 2 d after mock, V-Oka, or JoSt infection. A, Graphs
showing surface expression of group 1 CD1 molecules (MFI is indicated).
Data are representative of at least six experiments. B, Box-and-whisker
plot of MFI of CD1a (n = 4) and CD1c expression (n = 16). C, IFN-g
secretion by CD1c-restricted gd T cells cocultured with immature MDDCs
2 d after mock, V-Oka, or JoSt infection. One of six independent experiments done in triplicate is shown (error bars are 6 1 SD). D, Box-andwhisker plot of MFI of IFN-g secretion by CD1c-restricted gd T cells
cocultured with immature MDDCs 2 d after mock, V-Oka, or JoSt infection (n = 6). E, Anti-CD1c Abs block IFN-g secretion by CD1c-restricted
gd T cells cocultured with immature MDDCs 2 d after mock, V-Oka, or
JoSt infection. Anti-CD1c Abs or the respective isotype control were used
at 20 mg/ml. Data are representative of two independent experiments done
in triplicate.
possibility, VZV-infected immature MDDCs were stained with FITCAnnexin V and PI and analyzed by flow cytometry (Fig. 5C). Mockinfected immature MDDCs contained ,3% Annexin V single-positive
(apoptotic) cells and ,1% double-positive (necrotic) cells. By contrast,
VZVinduced apoptosis in 18–26% of immature MDDCs, regardless of
whether they were infected with the vaccine or VZV clinical isolates. In
all cases, the percentage of necrotic cells was ,5%. Moreover, there
was no difference between V-Oka and virulent VZV with regard to the
infection rate, as determined by viral gE expression on MDDCs (Fig.
5D). Thus, the functional difference between MDDCs infected with
V-Oka and MDDCs infected with clinical isolates was not due to
different survival rates of infected DCs or different infection rates. In
conclusion, virulent VZV, irrespective of its genotype and geographical
origin, interferes with the capacity of immature DCs to stimulate IFN-g
secretion by CD1c-restricted gd T cells.
DCs infected with virulent VZV are resistant to Th1-promoting
instruction
Innate lymphocytes, such as CD1c-restricted gd T cells, confer on
immature MDDCs the capacity to efficiently prime Th1 cells,
a process called DC instruction (13, 15, 27). We investigated
whether VZV-infected immature MDDCs were still susceptible
to instruction by CD1c-restricted gd T cells. For this purpose,
the expression of CD83 (Fig. 6A) and CD86 (Fig. 6B) on VZVinfected immature MDDCs was determined in the absence or
presence of gd T cells. As a control, immature MDDCs were
treated with LPS. In the absence of gd T cells, VZV-infected
immature MDDCs did not show a mature phenotype. Surprisingly,
immature MDDCs, whether infected with the vaccine V-Oka or
the clinical isolate JoSt, phenotypically matured after adding gd
T cells or LPS. We next studied the ability of CD1c-restricted gd
T cells to induce VZV-infected immature MDDCs to secrete IL12, a key cytokine for the stimulation of Th1 cell responses (Fig.
6C). In the absence of gd T cells, neither mock-infected nor VZVinfected immature MDDCs secreted bioactive IL-12p70. Strikingly, in the presence of gd T cells, immature MDDCs infected
with vaccine strain V-Oka secreted increased amounts of IL-12p70
into the supernatant compared with mock-infected MDDCs. By
contrast, immature MDDCs infected with VZV clinical isolates
and cocultured with gd T cells failed to produce significant
amounts of IL-12p70. Similar to the European VZV strains, a Japanese VZV clinical isolate belonging to the genotype J, like
V-Oka, blocked the induction of IL-12 release in cocultures of
MDDCs with gd T cells (data not shown). To assess the relevance
of IL-12 by MDDCs on IFN-g secretion by gd T cells, we added
neutralizing IL-12 Abs to the cultures. The addition of neutralizing IL-12 Abs blocked .50% of IFN-g secretion by gd T cells
when cultured with V-Oka–infected immature MDDCs (Fig. 6D).
These results indicated that the VZV vaccine and virulent VZV
allow phenotypic maturation of immature MDDCs upon cross-talk
with innate lymphocytes. Strikingly, however, only the VZV vaccine promotes functional instruction of MDDCs, leading to the
release of Th1-polarizing IL-12.
Virulent VZV and the VZV vaccine synergize with stimulation
through CD40
Next, we analyzed whether disruption of DC instruction by virulent
VZV was due to a viral block in CD40 signaling that is crucial for
inducing IL-12 secretion by MDDCs (28). Postinfection with the
clinical isolate JoSt, upregulation of CD40 on the surface of immature MDDCs was impaired compared with cells infected with
the VZV vaccine (Fig. 7A, 7B). Similarly, fewer CD40-encoding
transcripts were found in JoSt-infected immature MDDCs
Downloaded from http://www.jimmunol.org/ by guest on June 18, 2017
FIGURE 4. Viral transmission from VZV-infected DCs to gd T cells.
CD1c-restricted gd T cells were cocultured with immature MDDCs 2 d after
mock, V-Oka, or JoSt infection. A, The T cell population was gated according the FSC/SSC profile and analyzed for the coexpression of gd TCR
and the viral gE (percentage of cells is indicated in each quadrant). Data
are representative of three independent experiments. B, IFN-g secretion
by CD1c-restricted gd T cells cocultured with VZV-infected immature
MDDCs after stimulation with PHA (100 ng/ml). One of three independent
experiments done in triplicate is shown (error bars are 6 1 SD).
The Journal of Immunology
493
compared with V-Oka–infected immature MDDCs (data not
shown). However, after stimulation with CD40L-expressing fibroblasts, JoSt-infected and V-Oka–infected immature MDDCs
secreted more IL-12p70 than mock-infected immature MDDCs
(Fig. 7C). This increased IL-12 release was mirrored by enhanced
CD40L-mediated activation of the MAPK pathway in VZV-infected
DCs compared with mock-infected DCs (data not shown). Next, gd
T cells were added to the cultures, and IFN-g secretion was measured
(Fig. 7D). Intriguingly, the ability of JoSt-infected immature DCs
to stimulate IFN-g secretion by gd T cells was fully rescued in the presence of CD40L-expressing fibroblasts. Collectively,
these data demonstrate that despite blocking CD40 upregulation on
FIGURE 6. Phenotype and function of
VZV-infected DCs after instruction by innate
cells. VZV-infected immature MDDCs were
left untreated as a control, cocultured with gd
T cells, or stimulated with LPS (1 mg/ml).
After 24 h, cell surface expression of CD83
(A, numbers indicate percentage of positive
cells) or CD86 (B, numbers indicate MFI)
was detected by flow cytometry. Graphs are
representative of three independent experiments. C, IL-12p70 secretion by VZVinfected immature MDDCs in the absence
or presence of gd T cells, as assessed by
ELISA 2 d postinfection. One of two independent experiments done in triplicate is
shown (error bars are 6 1 SD). D, IFN-g
secretion by gd T cells stimulated with
VZV-infected immature MDDCs in the presence of neutralizing polyclonal anti–IL-12
(5 mg/ml), as measured by ELISA 2 d postinfection. One of two independent experiments done in triplicate is shown (error bars
are 6 1 SD).
Downloaded from http://www.jimmunol.org/ by guest on June 18, 2017
FIGURE 5. Influence of VZV genotype on DC
function and DC apoptosis. A, Phylogenetic tree of
virulent VZV strains circulating in Europe (M1_935/
05, E2_769/05, E1_667/05, and JoSt) with respect to
the reference strains V-Oka, CA123, VZV11, and
Dumas (in italics). The length of the indicated marker
represents 0.1 nucleotide exchanges. B, IFN-g secretion
by CD1c-restricted gd T cells cocultured with immature MDDCs 2 d postinfection with different VZV genotypes. One of three independent experiments done in
triplicate is shown (error bars are 6 1 SD). C, Apoptotic death of immature MDDCs 2 d postinfection. Cells
were stained with FITC-Annexin V and PI (percentage
of cells is indicated in each quadrant). Data are representative of three independent experiments. D, Cell
surface expression of viral gE on immature MDDCs
2 d postinfection. Percentages of gE+ cells are indicated. One of three independent experiments is shown.
494
VIRAL DISRUPTION OF DENDRITIC CELL INSTRUCTION
FIGURE 8. TLR signaling in VZV-infected DCs. Immature MDDCs were
left untreated (co); were mock, V-Oka, or JoSt infected (black columns); or
were stimulated with ultraviolet-inactivated mock-, V-Oka–, or JoSt-infected
cells (white columns) for 2 d. In addition, immature MDDCs were infected
with HSV for 12 h. Thereafter, cells were stimulated for 48 h with TLR2 ligand
LTA (A, 1 mg/ml) or TLR3 ligand poly(I:C) (B, 25 mg/ml). Secretion of bioactive IL-12 was quantified by ELISA. One representative experiment of two
is shown (error bars are 6 1 SD).
infected DCs, virulent VZV amplifies CD40 signaling, similar to the
VZV vaccine.
Virulent VZV selectively inhibits TLR2 signaling
Next, we investigated DC priming by signaling through TLRs,
a prerequisite of DC instruction (13, 29). In the absence of innate
cells, signaling through multiple PRRs can synergize to efficiently
prime IL-12 secretion by DCs (17–19). Therefore, we tested
whether VZV can cooperate with signaling through TLR2 and
TLR3, which are involved in the detection of HSV and VZV,
the human alphaherpesviruses (30–33). They are located on the
cell surface (TLR2) and endosomes (TLR3). TLR2 recognizes
viral glycoproteins in the viral envelope, whereas TLR3 detects
dsRNA. VZV-infected immature DCs were challenged with lipoteichoic acid (LTA), a TLR2 agonist, or polyinosinic:polycytidylic acid [poly(I:C)], a TLR3 agonist. As shown in Fig. 8A,
uninfected or mock-infected DCs treated with LTA did not secrete
IL12p70. Intriguingly, DCs infected with V-Oka or HSV released
large amounts of IL-12p70 after challenge with LTA. In sharp
contrast, DCs infected with JoSt, the virulent VZV strain, secreted
minimal amounts of IL-12 in response to LTA. Strikingly, efficient
priming of DCs required viral replication, because LTA did not
synergize with ultraviolet-inactivated virus to induce the release of
IL-12. Similar to LTA, the TLR3 agonist poly(I:C) could not induce uninfected or mock-infected DCs to secrete substantial
amounts of IL-12 (Fig. 8B). However, V-Oka and, to a lesser extent,
virulent VZV, but not HSV, cooperated with poly(I:C) to trigger IL-12
secretion. DCs infected with ultraviolet-inactivated virus did not respond to poly(I:C), as observed for LTA. Taken together, these data
demonstrated that replicating VZV can prime DCs for IL-12 release.
However, there was a fundamental difference between V-Oka and
virulent VZV. The former synergized with TLR2 and TLR3, whereas
the latter selectively blocked signaling through TLR2, which is
known to detect virulent VZV.
Discussion
In this study, we demonstrated the immunological difference
between V-Oka, the vaccine strain, and VZV clinical isolates: instruction of inflammatory DCs toward a Th1-polarizing program is
promoted by V-Oka but disrupted by virulent VZV.
We showed that different DC subsets are susceptible to VZV
infection. LCs and dermal DCs isolated from normal human skin
could be infected by VZV. Other investigators demonstrated recently that plasmacytoid DCs isolated from human blood and LCs
generated from MUTZ-3 cells are permissive to VZV infection
(34). Additionally, immature DCs generated in vitro from monocytes were susceptible to VZV infection, as previously reported
(10, 11). The majority of these MDDCs expressed viral gE at low
levels; only 10–20% showed strong surface expression of this
molecule. The distinct peaks can be explained by the highly
cell-associated nature of VZV. For VZV infection, MDDCs have
to be centrifuged on a VZV-infected fibroblast monolayer. During
the first round of virus transmission, the virus infects only a few
MDDCs because of the close cell-to-cell contact required. In this
induction phase, innate responses are weak, and the virus multiplies efficiently, resulting in strong viral gE surface expression. In
contrast, most MDDCs are probably infected later when the innate
defense is already fully established in the cell culture, resulting in
less efficient viral replication and low viral gE surface expression.
Importantly, however, there was no difference with regard to the
infection efficiency between the VZV vaccine and virulent VZV.
Skin biopsies from zoster lesions revealed that LCs disappear in
the vicinity of virus-containing vesicles, confirming other reports
(34, 35). This could have been due to virus-induced apoptosis.
Alternatively, VZV-infected LCs could have emigrated to the regional lymph nodes to transfer VZV Ag to resident DCs. In contrast, inflammatory DCs strongly infiltrated VZV-infected skin.
Similar observations were made for lepromatous skin lesions in
humans (8) and in an experimental mouse model of leishmanial
infection in the dermis (9). It is likely that the majority of VZVinfected inflammatory DCs had already undergone apoptosis or
migrated to the lymph nodes at the time when the skin punch
biopsies were taken. This could explain the scarcity of CD1c+
cells expressing viral gE in papulovesicular lesions, a relatively
late stage of VZV infection in the dermis. Alternatively, VZVinfected inflammatory DCs might be difficult to detect in skin
sections. This assumption is supported by the fact that most
Downloaded from http://www.jimmunol.org/ by guest on June 18, 2017
FIGURE 7. IL-12 production by VZV-infected DCs after CD40 triggering. A, Surface expression of CD40 on VZV-infected immature MDDCs 2 d
postinfection (MFI is indicated). Data are representative of 11 experiments.
B, Box-and-whisker plot of CD40 expression (MFI) on VZV-infected immature MDDCs (n = 11). C, IL-12 release by VZV-infected immature MDDCs
cocultured for 48 h with CD40L-expressing fibroblasts or CD40L2 control
cells. One of three independent experiments done in triplicate is shown
(error bars are 6 1 SD). D, IFN-g secretion by CD1c-restricted gd T cells
cocultured with VZV-infected immature DCs in the presence of CD40Lexpressing fibroblasts or CD40L2 control cells. One of four independent
experiments done in triplicate is shown (error bars are 6 1 SD).
The Journal of Immunology
inability of DCs infected with virulent VZV strains to stimulate
IFN-g secretion by CD1c-restricted gd T cells was not due to a viral
block in upregulation of costimulatory molecules (signal 2). However, only the VZV vaccine licensed release of IL-12 (signal 3),
a potent inducer of IFN-g secretion from T cells and of Th1 cell
differentiation. By contrast, all DCs infected with VZV clinical
isolates, regardless of their genotype, were unable to secrete significant amounts of IL-12 upon interaction with CD1c-restricted gd
T cells. Neutralization of IL-12 by Abs during coculture of V-Oka–
infected DCs with CD1c-restricted gd T cells decreased IFN-g
secretion 60–70%. These results emphasize that DC instruction is
a bidirectional process, whereby DCs and innate lymphocytes
stimulate each other (43, 47).
In addition to IFN-g, a second signal provided by CD40
engagement is required for triggering IL-12 secretion by DCs
(48, 49). We observed a moderate, although significant, increase
in CD40 on vaccine strain-infected DCs, whereas DCs infected
with virulent VZV did not show this effect. However, this difference in CD40 surface expression on infected DCs was functionally irrelevant. In comparison with unstimulated control cells,
JoSt-infected and V-Oka–infected DCs released large amounts
of IL-12 after stimulation with CD40L-expressing cells. Remarkably, the CD40L-induced IL-12 secretion by VZV-infected DCs
was even greater than the amount of IL-12 measured after stimulation of mock-infected DCs. This result supports the concept
that IL-12 release triggered by CD40–CD40L interaction is amplified by pathogen-derived danger signals (19). Additionally,
analysis of the CD40 signaling pathway in VZV-infected DCs
cocultured with CD40L-expressing cells revealed enhanced phosphorylation of JNK, ERK, and p38 kinases compared with mockinfected DCs (data not shown). In this regard, DCs infected with
virulent VZV differ from HSV-infected DCs, which do not produce IL-12 in response to CD40L-expressing cells (50). Finally,
IFN-g secretion by the cocultured gd T cells could be fully
rescued after stimulating virulent VZV-infected DCs with
CD40L-expressing cells. These data clearly showed that virulent
VZV synergizes with CD40 signaling as efficiently as the VZV
vaccine and does not target CD40 signaling to interfere with DC
instruction.
There was the possibility that virulent VZV subverts priming of
DCs for optimal IL-12 production, which requires synergistic signaling through multiple PRRs (17, 18). Multiple PRRs at different
cellular locations are involved in the detection of alphaherpesviruses, such as VZV and HSV. Viral glycoproteins within the viral
envelope are recognized by TLR2 at the cell surface (30–32). In
contrast, TLR3, which recognizes dsRNA, is located in endosomes and was shown to contribute to the innate response against
HSV (33). Dual recognition of HSV by different TLRs is crucial
to optimize the antiviral response (51). We observed that signaling
through TLR2 or TLR3 in uninfected or mock-infected DCs did
not induce IL-12 release, confirming a previous report (18). In
contrast, DCs infected with VZV vaccine or HSV secreted high
amounts of IL-12 after stimulation with LTA, a TLR2 agonist.
Intriguingly, LTA-induced IL-12 release was abrogated in DCs
infected with virulent VZV. In contrast, a VZV clinical isolate
was shown to trigger TLR2 on stably transfected HEK293 cells
(32). This finding excludes the possibility that TLR2 simply
neglects virulent VZV because of polymorphisms that occur in
all nine VZV envelope glycoproteins (52). It is unknown which of
these glycoproteins acts as a TLR2 ligand. V-Oka and, to a lesser
extent, virulent VZV synergized with poly(I:C), a TLR3 agonist.
In contrast, HSV-infected DCs did not support TLR3 signaling.
The latter finding is in accordance with a recent report showing
that DCs use TLR3-independent pathways to prevent further
Downloaded from http://www.jimmunol.org/ by guest on June 18, 2017
MDDCs infected in vitro with VZV show only low gE expression
in FACS analysis.
The infiltrating DCs are derived from monocytes that are
recruited to the site of inflammation and resemble dermal DCs
found in normal skin. After maturation, they migrate to the regional
lymph nodes to induce a protective Th1 cell response (36). Recently, it was demonstrated in the mouse model that inflammatory
DCs are required to induce a Th1-polarized response that protects
against viruses (24). For this reason, we modeled inflammatory
DCs in vitro and analyzed in detail whether VZV can interfere
with their ability to couple innate to adaptive immune responses.
In accordance with a previous report, no changes in CD1a and
CD86 expression were detected postinfection of immature DCs
with JoSt, a VZV clinical isolate (10). Surprisingly, vaccine and
VZV clinical isolates significantly enhanced CD1c expression on
immature DCs. Although CD1c can present synthetic lipopeptides
that mimic N-terminally acylated peptides found in cellular and
viral proteins (37), the self ligand and possible viral ligands recognized by CD1c-restricted gd T cells are still unknown (23).
Strikingly, V-Oka–infected DCs, but not immature DCs infected
with VZV clinical isolates, could efficiently stimulate IFN-g
release by CD1c-restricted gd T cells. In addition, VZV was
demonstrated to interfere with IFN-g signal transduction via the
Jak/Stat pathway (38). Such mechanisms aiming at IFN-g release
or IFN-g signal transduction represent an important viral defense
strategy. First, IFN-g is a key cytokine polarizing the adaptive
immune response toward Th1 lymphocytes, which provide help
for cytotoxic T cells and can themselves lyse virus-infected cells.
Second, a rapid supply of IFN-g in an early phase of VZV
infection could prevent viral spread and skin lesions, through
noncytolytic virus clearance (39).
Viral pathogens have developed multiple mechanisms that
eliminate DCs (40). In our study, VZV clinical isolates and the
VZV vaccine induced apoptosis to a similar extent: 18–26% of
DCs underwent programmed cell death postinfection. Thus, the
lack of IFN-g release during cross-talk of DCs infected with virulent VZV with innate cells was not due to enhanced apoptosis
induction. Apoptotic debris from VZV-infected immature DCs
could be phagocytosed by uninfected bystander DCs that subsequently mature, migrate to the regional lymph nodes, and crosspresent VZV-derived peptides to conventional ab T lymphocytes.
Such a mechanism could explain why high frequencies of VZVspecific ab T cells are observed in immunocompetent individuals
after natural VZV infection (41), despite multiple and highly efficient viral immune-evasion mechanisms. In contrast, it is also
possible that viruses, such as VZV, trigger DC apoptosis to blunt
the antiviral immune response because apoptotic blebs may constitute a tolerogenic signal (42).
VZV is known to possess T cell tropism and can be transferred
from infected DCs to ab T cells (38). Infected ab T lymphocytes
in peripheral blood transport VZV to the skin during the cellassociated viremia of primary infection (2). We found that
infected DCs transmit VZV to CD1c-restricted gd T cells, suggesting that these intraepithelial lymphocytes contribute to viral
dissemination in the skin during reactivation of VZV from sensory
ganglia. However, infection with virulent VZV did not silence
CD1c-restricted gd T cells, suggesting that the pathogen disrupts
DC instruction at the site of DCs.
Upon interaction with innate lymphocytes, DCs upregulate the
surface expression of CD83 and CD86, secrete IL-12, and efficiently induce Th1-like adaptive immune responses (13, 15, 28,
29, 43–46). We observed that in the presence of gd T cells, DCs
acquired a fully mature phenotype, whether infected with the
vaccine V-Oka or the clinical isolate JoSt. Consequently, the
495
496
Acknowledgments
We thank T. Kaiser (Max Planck Institute for Infection Biology, Berlin, Germany) for assistance in flow cytometry; U. Noack, M. Bigalke, and M. Alexi for
excellent technical support; R.A. Kroczek (Robert Koch Institute, Berlin,
Germany) for providing CD40L-expressing fibroblasts; D. Ernst (Charité–
Universitätsmedizin Berlin) for help in performing immunohistochemistry
of skin biopsies; and B. Eilers (Max Planck Institute for Infection Biology)
for providing reagents.
Disclosures
The authors have no financial conflicts of interest.
References
1. Weber, O. 2000. Novel mouse models for the investigation of experimental drugs
with activity against human varicella-zoster virus. Antivir. Chem. Chemother. 11:
283–290.
2. Ku, C. C., L. Zerboni, H. Ito, B. S. Graham, M. Wallace, and A. M. Arvin. 2004.
Varicella-zoster virus transfer to skin by T Cells and modulation of viral replication by epidermal cell interferon-alpha. J. Exp. Med. 200: 917–925.
3. Gershon, A. A., and S. P. Steinberg. 1979. Cellular and humoral immune
responses to varicella-zoster virus in immunocompromised patients during and
after varicella-zoster infections. Infect. Immun. 25: 170–174.
4. Arvin, A. M., and A. A. Gershon. 1996. Live attenuated varicella vaccine. Annu.
Rev. Microbiol. 50: 59–100.
5. Oxman, M. N., M. J. Levin, G. R. Johnson, K. E. Schmader, S. E. Straus,
L. D. Gelb, R. D. Arbeit, M. S. Simberkoff, A. A. Gershon, L. E. Davis, et al;
Shingles Prevention Study Group. 2005. A vaccine to prevent herpes zoster and
postherpetic neuralgia in older adults. N. Engl. J. Med. 352: 2271–2284.
6. Abendroth, A., and A. M. Arvin. 2001. Immune evasion as a pathogenic
mechanism of varicella zoster virus. Semin. Immunol. 13: 27–39.
7. Kupper, T. S., and R. C. Fuhlbrigge. 2004. Immune surveillance in the skin:
mechanisms and clinical consequences. Nat. Rev. Immunol. 4: 211–222.
8. Krutzik, S. R., B. Tan, H. Li, M. T. Ochoa, P. T. Liu, S. E. Sharfstein,
T. G. Graeber, P. A. Sieling, Y. J. Liu, T. H. Rea, et al. 2005. TLR activation
triggers the rapid differentiation of monocytes into macrophages and dendritic
cells. Nat. Med. 11: 653–660.
9. León, B., M. López-Bravo, and C. Ardavı́n. 2007. Monocyte-derived dendritic
cells formed at the infection site control the induction of protective T helper 1
responses against Leishmania. Immunity 26: 519–531.
10. Abendroth, A., G. Morrow, A. L. Cunningham, and B. Slobedman. 2001. Varicellazoster virus infection of human dendritic cells and transmission to T cells: implications for virus dissemination in the host. J. Virol. 75: 6183–6192.
11. Hu, H., and J. I. Cohen. 2005. Varicella-zoster virus open reading frame 47
(ORF47) protein is critical for virus replication in dendritic cells and for spread
to other cells. Virology 337: 304–311.
12. Morrow, G., B. Slobedman, A. L. Cunningham, and A. Abendroth. 2003. Varicellazoster virus productively infects mature dendritic cells and alters their immune
function. J. Virol. 77: 4950–4959.
13. Leslie, D. S., M. S. Vincent, F. M. Spada, H. Das, M. Sugita, C. T. Morita, and
M. B. Brenner. 2002. CD1-mediated gamma/delta T cell maturation of dendritic
cells. J. Exp. Med. 196: 1575–1584.
14. Vieira, P. L., E. C. de Jong, E. A. Wierenga, M. L. Kapsenberg, and P. Kalinski.
2000. Development of Th1-inducing capacity in myeloid dendritic cells requires
environmental instruction. J. Immunol. 164: 4507–4512.
15. Vincent, M. S., D. S. Leslie, J. E. Gumperz, X. Xiong, E. P. Grant, and M. B. Brenner.
2002. CD1-dependent dendritic cell instruction. Nat. Immunol. 3: 1163–1168.
16. Gautier, G., M. Humbert, F. Deauvieau, M. Scuiller, J. Hiscott, E. E. Bates,
G. Trinchieri, C. Caux, and P. Garrone. 2005. A type I interferon autocrineparacrine loop is involved in Toll-like receptor-induced interleukin-12p70 secretion by dendritic cells. J. Exp. Med. 201: 1435–1446.
17. Napolitani, G., A. Rinaldi, F. Bertoni, F. Sallusto, and A. Lanzavecchia. 2005.
Selected Toll-like receptor agonist combinations synergistically trigger a T helper
type 1-polarizing program in dendritic cells. Nat. Immunol. 6: 769–776.
18. Roses, R. E., S. Xu, M. Xu, U. Koldovsky, G. Koski, and B. J. Czerniecki. 2008.
Differential production of IL-23 and IL-12 by myeloid-derived dendritic cells in
response to TLR agonists. J. Immunol. 181: 5120–5127.
19. Schulz, O., A. D. Edwards, M. Schito, J. Aliberti, S. Manickasingham, A. Sher,
and C. Reis e Sousa. 2000. CD40 triggering of heterodimeric IL-12 p70 production by dendritic cells in vivo requires a microbial priming signal. Immunity
13: 453–462.
20. Aliahmadi, E., R. Gramlich, A. Grützkau, M. Hitzler, M. Krüger, R. Baumgrass,
M. Schreiner, B. Wittig, R. Wanner, and M. Peiser. 2009. TLR2-activated human
langerhans cells promote Th17 polarization via IL-1beta, TGF-beta and IL-23.
Eur. J. Immunol. 39: 1221–1230.
21. Sauerbrei, A., R. Zell, A. Philipps, and P. Wutzler. 2008. Genotypes of varicellazoster virus wild-type strains in Germany. J. Med. Virol. 80: 1123–1130.
22. Raftery, M. J., M. Hitzler, F. Winau, T. Giese, B. Plachter, S. H. E. Kaufmann,
and G. Schönrich. 2008. Inhibition of CD1 antigen presentation by human cytomegalovirus. J. Virol. 82: 4308–4319.
23. Spada, F. M., E. P. Grant, P. J. Peters, M. Sugita, A. Melián, D. S. Leslie,
H. K. Lee, E. van Donselaar, D. A. Hanson, A. M. Krensky, et al. 2000. Selfrecognition of CD1 by gamma/delta T cells: implications for innate immunity. J.
Exp. Med. 191: 937–948.
24. Nakano, H., K. L. Lin, M. Yanagita, C. Charbonneau, D. N. Cook, T. Kakiuchi,
and M. D. Gunn. 2009. Blood-derived inflammatory dendritic cells in lymph
nodes stimulate acute T helper type 1 immune responses. Nat. Immunol. 10:
394–402.
25. Raftery, M. J., F. Winau, S. H. Kaufmann, U. E. Schaible, and G. Schönrich.
2006. CD1 antigen presentation by human dendritic cells as a target for herpes
simplex virus immune evasion. J. Immunol. 177: 6207–6214.
26. Yuan, W., A. Dasgupta, and P. Cresswell. 2006. Herpes simplex virus evades
natural killer T cell recognition by suppressing CD1d recycling. Nat. Immunol.
7: 835–842.
27. Fujii, S., K. Shimizu, H. Hemmi, and R. M. Steinman. 2007. Innate Valpha14(+)
natural killer T cells mature dendritic cells, leading to strong adaptive immunity.
Immunol. Rev. 220: 183–198.
28. Fujii, S., K. Liu, C. Smith, A. J. Bonito, and R. M. Steinman. 2004. The linkage
of innate to adaptive immunity via maturing dendritic cells in vivo requires
CD40 ligation in addition to antigen presentation and CD80/86 costimulation. J.
Exp. Med. 199: 1607–1618.
29. Brigl, M., L. Bry, S. C. Kent, J. E. Gumperz, and M. B. Brenner. 2003. Mechanism of CD1d-restricted natural killer T cell activation during microbial infection. Nat. Immunol. 4: 1230–1237.
30. Aravalli, R. N., S. Hu, T. N. Rowen, J. M. Palmquist, and J. R. Lokensgard.
2005. Cutting edge: TLR2-mediated proinflammatory cytokine and chemokine
production by microglial cells in response to herpes simplex virus. J. Immunol.
175: 4189–4193.
31. Kurt-Jones, E. A., M. Chan, S. Zhou, J. Wang, G. Reed, R. Bronson,
M. M. Arnold, D. M. Knipe, and R. W. Finberg. 2004. Herpes simplex virus 1
interaction with Toll-like receptor 2 contributes to lethal encephalitis. Proc. Natl.
Acad. Sci. USA 101: 1315–1320.
32. Wang, J. P., E. A. Kurt-Jones, O. S. Shin, M. D. Manchak, M. J. Levin, and
R. W. Finberg. 2005. Varicella-zoster virus activates inflammatory cytokines in
human monocytes and macrophages via Toll-like receptor 2. J. Virol. 79: 12658–
12666.
33. Zhang, S. Y., E. Jouanguy, S. Ugolini, A. Smahi, G. Elain, P. Romero, D. Segal,
V. Sancho-Shimizu, L. Lorenzo, A. Puel, et al. 2007. TLR3 deficiency in patients
with herpes simplex encephalitis. Science 317: 1522–1527.
34. Huch, J. H., A. L. Cunningham, A. M. Arvin, N. Nasr, S. J. Santegoets,
E. Slobedman, B. Slobedman, and A. Abendroth. 2010. Impact of varicellazoster virus on dendritic cell subsets in human skin during natural infection.
J. Virol. 84: 4060–4072 .
35. Drijkoningen, M., C. De Wolf-Peeters, H. Degreef, and V. Desmet. 1988.
Epidermal Langerhans cells, dermal dendritic cells, and keratinocytes in viral
lesions of skin and mucous membranes: an immunohistochemical study. Arch.
Dermatol. Res. 280: 220–227.
36. León, B., and C. Ardavı́n. 2008. Monocyte-derived dendritic cells in innate and
adaptive immunity. Immunol. Cell Biol. 86: 320–324.
37. Van Rhijn, I., D. C. Young, A. De Jong, J. Vazquez, T. Y. Cheng, R. Talekar,
D. C. Barral, L. León, M. B. Brenner, J. T. Katz, et al. 2009. CD1c bypasses
lysosomes to present a lipopeptide antigen with 12 amino acids. J. Exp. Med.
206: 1409–1422.
38. Abendroth, A., B. Slobedman, E. Lee, E. Mellins, M. Wallace, and A. M. Arvin.
2000. Modulation of major histocompatibility class II protein expression by
varicella-zoster virus. J. Virol. 74: 1900–1907.
Downloaded from http://www.jimmunol.org/ by guest on June 18, 2017
dissemination of HSV (33). In conclusion, our results suggest that
virulent VZV actively interferes with signaling downstream of
surface TLR2. It was shown that vaccinia virus and hepatitis C
virus encode proteins that block intracellular TLR signaling components (53, 54). Similarly, virulent VZV might encode a TLR
antagonist that disrupts intracellular TLR2 signaling.
The molecular mechanisms underlying attenuation of the VZV
vaccine are not understood. Our results showed that virulent VZV
disrupts instruction of DCs toward a Th1-polarizing program. During DC instruction by innate cells, virulent VZV strongly blocked
the release of IL-12 (signal 3). As an underlying mechanism, we
identified a viral block of synergistic signaling through TLR2,
which detects an unknown VZV envelope glycoprotein. The capacity of virulent VZV to interfere with DC instruction may be crucial
for extensive and, therefore, symptomatic viral replication in the
skin after reactivation from cranial and dorsal root ganglia. Moreover, this immune-evasion strategy is likely to be relevant for the
viral spread from infected mucosa in the respiratory tract to the skin
during primary infection. Our findings expand our understanding of
VZV-associated pathogenesis and have practical consequences.
Novel vaccination approaches have to verify that the candidate vaccines allow priming of DCs for IL-12 secretion, which is essential
for the timely induction of an efficient antiviral immune response
that prevents disease.
VIRAL DISRUPTION OF DENDRITIC CELL INSTRUCTION
The Journal of Immunology
39. Guidotti, L. G., and F. V. Chisari. 2001. Noncytolytic control of viral infections
by the innate and adaptive immune response. Annu. Rev. Immunol. 19: 65–91.
40. Hay, S., and G. Kannourakis. 2002. A time to kill: viral manipulation of the cell
death program. J. Gen. Virol. 83: 1547–1564.
41. Diaz, P. S., S. Smith, E. Hunter, and A. M. Arvin. 1989. T lymphocyte cytotoxicity with natural varicella-zoster virus infection and after immunization with
live attenuated varicella vaccine. J. Immunol. 142: 636–641.
42. Tournier, J. N., and A. Quesnel-Hellmann. 2006. Host-pathogen interactions:
a biological rendez-vous of the infectious nonself and danger models? PLoS
Pathog. 2: e44.
43. Gerosa, F., B. Baldani-Guerra, C. Nisii, V. Marchesini, G. Carra, and
G. Trinchieri. 2002. Reciprocal activating interaction between natural killer
cells and dendritic cells. J. Exp. Med. 195: 327–333.
44. Hermans, I. F., J. D. Silk, U. Gileadi, M. Salio, B. Mathew, G. Ritter, R. Schmidt,
A. L. Harris, L. Old, and V. Cerundolo. 2003. NKT cells enhance CD4+ and CD8+
T cell responses to soluble antigen in vivo through direct interaction with dendritic
cells. J. Immunol. 171: 5140–5147.
45. Ismaili, J., V. Olislagers, R. Poupot, J. J. Fournié, and M. Goldman. 2002. Human
gamma delta T cells induce dendritic cell maturation. Clin. Immunol. 103: 296–302.
46. Mocikat, R., H. Braumüller, A. Gumy, O. Egeter, H. Ziegler, U. Reusch,
A. Bubeck, J. Louis, R. Mailhammer, G. Riethmüller, et al. 2003. Natural killer
cells activated by MHC class I(low) targets prime dendritic cells to induce
protective CD8 T cell responses. Immunity 19: 561–569.
47. Conti, L., R. Casetti, M. Cardone, B. Varano, A. Martino, F. Belardelli, F. Poccia,
and S. Gessani. 2005. Reciprocal activating interaction between dendritic cells
497
48.
49.
50.
51.
52.
53.
54.
and pamidronate-stimulated gammadelta T cells: role of CD86 and inflammatory
cytokines. J. Immunol. 174: 252–260.
Cella, M., D. Scheidegger, K. Palmer-Lehmann, P. Lane, A. Lanzavecchia, and
G. Alber. 1996. Ligation of CD40 on dendritic cells triggers production of high
levels of interleukin-12 and enhances T cell stimulatory capacity: T-T help via
APC activation. J. Exp. Med. 184: 747–752.
Snijders, A., P. Kalinski, C. M. Hilkens, and M. L. Kapsenberg. 1998. High-level
IL-12 production by human dendritic cells requires two signals. Int. Immunol.
10: 1593–1598.
Salio, M., M. Cella, M. Suter, and A. Lanzavecchia. 1999. Inhibition of dendritic
cell maturation by herpes simplex virus. Eur. J. Immunol. 29: 3245–3253.
Sato, A., M. M. Linehan, and A. Iwasaki. 2006. Dual recognition of herpes
simplex viruses by TLR2 and TLR9 in dendritic cells. Proc. Natl. Acad. Sci.
USA 103: 17343–17348.
Storlie, J., L. Maresova, W. Jackson, and C. Grose. 2008. Comparative analyses
of the 9 glycoprotein genes found in wild-type and vaccine strains of varicellazoster virus. J. Infect. Dis. 197(Suppl. 2): S49–S53.
Li, K., E. Foy, J. C. Ferreon, M. Nakamura, A. C. Ferreon, M. Ikeda, S. C. Ray,
M. Gale, Jr., and S. M. Lemon. 2005. Immune evasion by hepatitis C virus NS3/
4A protease-mediated cleavage of the Toll-like receptor 3 adaptor protein TRIF.
Proc. Natl. Acad. Sci. USA 102: 2992–2997.
Stack, J., I. R. Haga, M. Schröder, N. W. Bartlett, G. Maloney, P. C. Reading,
K. A. Fitzgerald, G. L. Smith, and A. G. Bowie. 2005. Vaccinia virus protein
A46R targets multiple Toll-like-interleukin-1 receptor adaptors and contributes
to virulence. J. Exp. Med. 201: 1007–1018.
Downloaded from http://www.jimmunol.org/ by guest on June 18, 2017