Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
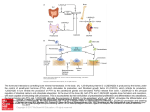
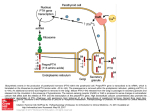
Clinical Science( 1986) 71, 231-238 23 1 ~ EDITORIAL REVIEW Parathyroid hormone and Lone J. REEVE’ AND JOAN M. ZANELLI’ I Bone Disease Research Group, MRC Clinical Research Centre and Northwick Park Hospital, Harrow, Middlesex, U.K., and ZHormonesDivision, National Institute for Biological Standards and Control, Harnpstead, London Introduction Nearly sixty years ago Mandl demonstrated that removal of a parathyroid tumour could reverse the associated bone softening and cyst formation of osteitis fibrosa [ l , 21. Over the next 20 years, various investigators showed that animals treated with impure parathyroid extract (PTE) in moderate dosage given daily or on alternate days could increase their trabecular bone density [3-91. In these experiments, in the first few days, osteoclastic resorption and fibrous tissue proliferation were observed; an increase in osteoblastic activity followed later [7]. Higher doses of PTE led to the more familiar pictures of massive continued bone loss or osteitis fibrosa [4,9]. Considerable advances have since been made in understanding how parathyroid peptides stimulate the cellular events leading to bone renewal, both physiologically and in pathological states in which high plasma concentrations of parathyroid hormone (PTH) are found. However, their precise role in the regulation of the balance between bone formation and resorption still remains obscure. Within a small volume of bone, at the intermediary level between the cell and the organ, tissue renewal is organized by a co-operating group of cells, named the ‘basic multicellular unit’ (BMU)by Frost [lo]. The normal skeleton contains 105-106 BMU. Some cells (eg. osteoblasts and osteocytes) are derived from the stromal (fibroblastic) system of bone or marrow [ll]; osteoclasts are believed to originate in the haemopoietic system and in some respects resemble macrophages [ 121, but do not share their surface markers [ 131. Previous speculations that the two bone cell types shared a common stem cell [ 141have been disproved [ 111. Abbreviations: BMU, basic multicellular unit(s); PTE, parathyroid extract; PTH, parathyroid hormone; hPTH 1-34, 1-34 fragment of synthetic human PTH. Correspondence: Dr J. Reeve, Bone Disease Research Group, Clinical Research Centre, Watford Road, Harrow, Middlesex HA1 3UJ, U.K. Normally, in man, iliac trabecular bone remodelling takes about 4 months. Resorption is initiated by osteoclasts, perhaps prompted by another cell type. First a collagen-poor cement line is laid down during a phase when the resorption surface is predominantly populated with cells with single nuclei, including pre-osteoblasts [ 151; bone formation then begins 5-10 weeks after the initiation of bone resorption [16]. Osteoid is laid down on the cement line and the lag time to its mineralization is normally 12 days or more, being very much longer in diseases such as osteomalacia [ 171. However, even in health not all osteoid is mineralizing simultaneously; in normal man 30% appears to be ‘resting’ [lo, 181. After remodelling is complete it is thought that perhaps 30% of osteoblasts survive to transform into resting bone surface or ‘buried‘ osteocytes, which have distinctive responses to PTH in vivo as will be discussed. Because osteocytes do not make bone in significant amounts, an important functional distinction is made by histomorphometrists between these cells and their parents, the osteoblasts. The origin of the two functionally distinct cell types from the same lineage presents potential difficulties in the interpretation of studies in vitro of bone cells grown in culture. In this review, the influence of PTH peptides on bone tissue in vivo and in vitro is contrasted with their effects on isolated cells. An attempt is made to identify those problems which must be addressed in future research. For an excellent in-depth review of this topic up to 1976, readers are referred to Parfitt [19-221 and earlier information was summarized by Parsons & Potts [23]. Effects of PTH on cultured bone tissue Studies by Barnicot [24] and Chang [25] on the osteolytic effects of parathyroid tissue transplanted into close proximity to cranial bone heralded the development of tissue culture techniques by Gail- 232 J. Reeve and J. M . Zatielli lard [26]for studying effects of parathyroid peptides on bone in vitro. Bone from neonates, or fetal bone from the last third of the gestational period, was found to dissolve over a few days in culture in the presence of high concentrations of PTE [26-321. The degrees of histological and biochemical changes seen in the bone and culture fluid were in some cases proportional to the concentration of added parathyroid activity, leading to the development of bioassay systems [28, 321. Gaillard showed that PTE increased both the number and activity of osteoclasts [27]. More recently, Stern et al. [33] have found that a calcium ionophore, which increases intracellular calcium, promotes osteoclast formation, and Lowik et al. [34] using the indicator Quin 2 have demonstrated that intracellular calcium is increased by PTH. Parathyroid hormone is well known to increase cyclic AMP production in the kidney [35] and some bone systems [36, 371. It may therefore act on cells by increasing the intracellular concentrations of two recognized secondary messengers which may act independently or synergistically [38]. Further progress in understanding the biochemical actions of PTH at the target cell membrane would be facilitated by the characterization of the PTH receptor molecule(s), a task which has presented considerable techniml difficulties [39]. Using longer-term cultures, tioldhaber showed that despite the early disappearance of osteoblasts, spontaneous bone resorption was eventually followed by bone formation [30]. Subsequently, Herrmann-Erlee et al. [40] showed that, at low concentrations, PTH and particularly its 1-34 human N-terminal fragment caused net increases in the amount of bone formed in vitro. Howard et al. obtained a soluble factor (molecular weight approximately 83 000 daltons) released from bone after stimulation by bone resorbing agents such as PTH, which induced subsequent bone formation [41], probably because it was mitogenic for osteoblast precursors [42]. This harked back to earlier work of Bingham et al. [43], who showed that tritiated uridine incorporation into osteoblast-like cells showed a biphasic response to PTH. Effects of PTH on bone cells in v i m The difficulties of unravelling the biochemical mechanisms of PTH actions on cell populations in intact bone have emphasized the importance of studying PTH effects on isolated populations of identified bone cells. However, techniques for purifying bone cells have not yet progressed to the point at which functionally homogeneous preparations of osteoblastic or osteoclastic cell lines can be obtained. By studying separated individual multinucleated osteoclasts which can be shown under the microscope to resorb bone, Chambers and his colleagues [44, 451 have circumvented some of the difficulties associated with mixed cell cultures and shed light on the relationship between PTH and osteoclastic bone resorption. Whereas calcitonin inhibited the motility of osteoclasts and prevented them from creating resorption pits on slices of devitalized cortical bone, PTH in various concentrations was without any effect. Braidman and her colleagues [46] used an enzymatic digestion procedure applied to early postnatal calvaria followed by Percoll density centrifugation to obtain cultures or osieoclast-like cells more homogeneous than those previously obtained with the techniques of Wong & Cohn [47]. Whereas Wong & Cohn [48] had observed that their ‘osteoclast-like’ cells responded to PTH, Braidman et al. did not [46], supporting the conclusions of Chambers et al. Several approaches to the study of osteoblasts in culture have been developed. The sequential enzyme digestion technique of Wong & Cohn [48] was applied to embryonic or neonatal bone and used to harvest cells released late during digestion. These cells had morphological and biochemical characteristics of osteoblasts, and responded to PTH, but not to calcitonin [49]. In Braidman’s refined system these observations were confirmed [46]. Another approach has been to culture bone marrow cells in media which favour stromal cell populations. Fibroblastic cells with many characteristics of osteoblasts have been grown, cloned and serially passaged from rabbit, guinea pig and human bone marrow [50, 511. Rabbit cells, when enclosed within diffusion chambers implanted into the peritoneal cavities of host rabbits (separating the cells from host cells, but not from macromolecules), form bone, cartilage and fibrous tissue in variable proportions [52, 531. Further experiments with cloned cells by Friedenstein, who reimplanted them under the kidney capsule in uivo [54], have shown that a single clone can develop a whole stromal environment for colonization by host haemopoietic cells to form a marrow ‘organ’.This and other work summarized by Owen [ 111 has led to the realizations that osteoblasts may share a common stromal stem cell, not only with osteocytes, but with other marrow cell types such as the fibroblastic cells of the marrow reticular network, chondroblasts and adipocytes [ 11, 551. It is also generally agreed that many osteoblasts eventually transform into osteocytes in vivo, but the possibility that analogous changes may occur in long-term bone cell cultures in vitro has not yet received attention. Parathyroid hormone and bone Mixed cell cutures are well known to cause difficulties in studying single cell types because alterations occur in relative growth rates when different cell types are grown together in vitro. Therefore it is important to recognize that studies on ‘osteoblastlike’ cells grown in mixed cultures will require validation when techniques become available to differentiate further cells able to form bone from cells which are not. In particular, since the proportion of ‘osteoblasts’ in a cell culture population may decrease under culture conditions, negative results from experiments should be treated with caution. As an example of this problem from another field, Tyrrell et al. were able to show that the reported insensitivity of many mixed cultures of pneumocytes to respiratory virus infections was due to the relative disappearance in culture of the rather slowly dividing type II pneumocytes; when cloned, the vulnerability of the type II cells to infection was demonstrable [56]. However, cell cloning alone will not solve the problem of obtaining pure preparations of osteoblasts or osteocytes if each colony is generated by a multipotential ‘stem’ cell. Because of such problems, a number of groups have used malignant cell lines derived or cloned from induced transplantable osteogenic sarcomata from rats, to study the biochemical interactions between PTH peptides and the osteoblast-like cell [57-601. These studies have been useful in defining the actions of PTH on a number of cellular activities [60]. However, the intrinsic growth potential of transformed cells, with their tendency to produce transforming growth factors, renders them unsuitable for consideration of the effect of PTH on growth patterns of normal osteoblasts. Based on studies with transformed and untransformed cells, certain conclusions may be drawn [60]. Isolated osteoblast-like cells respond to PTH with cyclic AMP production. They also respond to the prostaglandin PGE, and calcitriol (1,25-dihydroxyvitamin D). They are capable of producing collagenase, plasminogen activator [613, prostaglandins [60] and osteocalcin [62]. When added to the osteoclast culture system of Chambers et al., mixed populations of stromal cells which come in close proximity to active osteoclasts can promote their recovery from calcitonin-induced immobility [63]. Braidman et al., using cytochemical techniques, have shown that PTH indirectly stimulates an increase in osteoclast acid phosphatase activity, provided that the osteoclast is co-cultured with an ‘osteoblast-like’ cell 1641. This evidence of an indirect effect of PTH on osteoclasts may help resolve the paradox of the differing effects of PTH on osteoclasts in tissue culture and on isolated osteoclasts observed in the systems of Chambers and Braidman. 233 When continuously exposed to PTH, osteoblastlike cells reduce their synthesis of collagen [65]. On the other hand, short ( < 24 h) exposures of such cells to PTH increase their mitotic activity after transfer to a PTH-free medium [66]. Studies of PTH in vivo The intact organism integrates various processes which determine the overall effects of PTH on the skeleton. There is reason to believe that the first action of PTH is to modulate the number of BMU that are active in a given volume of bone [20]. Secondly, at the level of the individual BMU, PTH may influence the activity [27] of the osteoclasts and the functional lifespan of the osteoblasts [67], as well as the birth rates of the two cell types. The third action of PTH on bone is to promote, via an exchange with K + [68], calcium efflux from the so-called exchangeable pools of bone calcium in proximity to osteocytes [69, 701, which leads to a rapid increase in the plasma calcium level. It is believed that this effect is the result of a direct action of PTH on these cells, and that calcitonin exerts its rapid hypocalcaemic action by having the opposite effect [70]. With the action of PTH in causing renal retention of calcium [71], the osteocytes are substantially responsible for short-term plasma calcium homoeostasis, to which the other actions of PTH in promoting changes in bone turnover contribute comparatively little [711. Interestingly, the immediate effect of intravenously injected PTH on bone cells is to promote an influx of plasma calcium into bone [72];the more familiar calcaemic action of PTH develops subsequently, and in chicks may be augmented by prior intravenous injection of calcium [73]. After surgical removal of a parathyroid adenoma, Mazzuoli has observed a complementary response in that plasma calcium transiently rises before the expected fall [74]; termination of chronic hypercalcaemic PTH infusions in dogs gave results compatible with these observations [75]. All of these effects of PTH are influenced by its presentation to bone cell receptors. The pattern of exposure, as well as the integrated dose of PTH, might potentially affect the secondary responses of individual bone receptors as well as their density on individual cells. Therefore, in addition to the mean rate of hormone secretion, the glandular secretion pattern and the disposal dynamics of endogenous PTH are potentially important regulators of bone cell function. Indeed, Atkinson et al. have found evidence for altered PTH disposal in primary biliary cirrhosis (possibly due to Kupffer cell dysfunction) [76] and in low turnover idiopathic osteoporosis 1771. 234 J. Reeve and J. M. Zanelli When administered to the intact mammal in supraphysiological doses, PTH and its derivatives produce a remarkable range of effects. Iliac trabecular bone volume in primary hyperparathyroidism is usually normal or near-normal [67], with evidence of increased remodelling due to a corresponding increase in the rate of initiation of new BMU. In the trabecular bone of the distal radius, osteopenia is usually seen, as it is in the cortical bone of the periphery [78,79]. In primary hyperparathyroidism without overt osteitis fibrosa, the history of the osteoblastic phase of the typical axial trabecular BMU has been reconstructed by Charhon et al. [67]. Except in premenopausal women (the only sub-group to suffer a measurable deficit in axial trabecular bone), the effective lifespan of the osteoblasts was increased, although at the cell level there was a slight reduction in the rate of work of the individual osteoblast. In very severe hyperparathyroidism, when accompanied by osteitis fibrosa, the differing effects on cortical and trabecular bone may become more accentuated, with severe cortical osteopenia accompanying preserved values of axial trabecular bone volume. In hyperparathyroidism associated with renal failure, disordered vitamin D metabolism and hyperphosphataemia commonly modify the effects of excess parathyroid hormone concentrations, and axial trabecular bone volume may be increased. In contrast to clinical hyperparathyroidism and continuous infusions of active N-terminal parathyroid peptides [80,811, daily subcutaneous or intravenous injections of these peptides have been found to cause marked effects on indices of trabecular bone mass and turnover without resulting in more than transient post-injection hypercalcaemia [3-9, 82-84]. The diaphyseal sclerosis of rat long bones resulting from daily injections of a crude parathyroid extract was also shown to be due to PTH [84] rather than to calcitonin [85]. In patients with osteoporosis, a trial of the 1-34 fragment of synthetic human PTH (hPTH 1-34) given by daily injections was found to cause a substantial increase, to a mean of about 70% above baseline values, in trabecular bone volume in the iliac crest as well as increased indices of bone turnover [82]. Increases in trabecular bone volume were found to correlate with maximal achieved indices of bone formation [82]. A recent report has noted similar increases in the trabecular bone density of the lumbar spine, measured with computed tomography [86], although in this study calcitriol in low dosage was added to the treatment regimen. There have been suggestions that bone is more responsive to N-terminal active fragments of PTH than to the intact hormone. The synthetic 1-34 fragment caused a larger cyclic AMP response by bone than did the intact hormone in dogs [87] and these results have been confirmed in mice 1881. However, cyclic AMP has not been shown to mediate the physiological response of bone to PTH, and Herrmann-Erlee et al. found a dissociation between bone resorption and cyclic AMP response to PTH in tissue culture [89]. Studies were carried out in dogs and rats to investigate the mechanisms of the anabolic effects of hPTH 1-34. Daily injections markedly increased iliac trabecular bone volumes in both species [81, 831just as they did in patients with idiopathic osteoporosis [82]. In rats, daily injections were also found to result in increases in total body calcium relative to values in controls [go]. However, when administered by continuous intravenous or subcutaneous infusions, there were no increases in iliac trabecular bone [80,811, and the study by Tam et al. in rats [83] suggested that the difference was due to a relatively greater increase in resorption with the infusion regimen, associated with hypercalcaemia at higher doses. When given by a single subcutaneous injection to man, the appearance of hPTH 1-34 in the circulation was found to be brief [91], and the measurable effects on renal function to last little more than 4 h after injection [92]. It is generally appreciated that PTH bioassays in vivo which depend on the hypercalcaemic response to intravenously injected PTH involve massive, if transient, elevations of plasma PTH levels. For example, in the relatively sensitive chick hypercalcaemia assay [73], to achieve a rise in plasma calcium of 1.5 mmol/l it is necessary to raise the plasma PTH concentration by 4-5 orders of magnitude, assuming that bioactive PTH levels in man and the chick are comparable [93]. It therefore seems likely that the absence of hypercalcaemia associated with daily injections of parathyroid peptides is related to the brevity of the effects of such dose regimens on the kidney and the osteocytes regulating the exchangeable bone pools of calcium. In contrast, the increase in bone formation, which takes from 4 to 7 days in the rat [7] and probably longer than 5 weeks to develop in man [82, 941, implies that daily injections of parathyroid peptides require a similar period to cause a significant increase in the number of activated BMU. This encouraged the idea that these newly activated BMU could be further influenced by ‘coherence’ therapy [95, 961. This concept involves increasing the birthrate of new BMU by cyclic periods of treatment (eg. with PTH), after each of which osteoclastic resorption should be selectively depressed and bone formation encouraged to continue, mediated by the newly induced population of normally relatively long-lived osteoblasts. However, activation of a new BMU may require more than Parathyroid hormone and bone just a brief triggering stimulus with a parathyroid peptide. The only trial of such an approach so far reported with PTH,in which each of the 12 activation phases was of just 1 week‘s duration (and osteoclast depression was attempted by 3 weeks of PTH withdrawal), did not induce increased osteoblast activation despite clearcut success in depressing osteoclastic resorption [97].Thus the result of a total of 12 weeks’ treatment with hPTH 1-34 spread over a year was an overall reduction in bone turnover, in direct contrast to the daily injection regimen. There remain many fascinating questions concerning the interaction of other agents with the PTH-bone cell relationship. In normal women, the role of oestrogens in reducing bone resorption relative to formation [98]might be through a modification of the response of bone to PTH. The biochemical basis for such an action remains obscure; as yet there is no convincing demonstration of oestrogen receptors in bone. Thyroid hormone concentrations, besides affecting overall levels of remodelling activity, may influence the balance between bone formation and resorption [95, 991, as glucocorticoids certainly do [loo]. Malignant cells may markedly influence responses to endogenous PTH either locally through secretion of cytokines [ l o l l or at a distance by secretion of circulating factors which have some of the characteristics of PTHitself [ 1021. Conclusions and future investigations Despite the vast literature that has accumulated on the action of PTH on bone, this remains a fertile field for future investigation.There is now a general acceptance that parathyroid peptides act directly on cells of the stromal system leading to osteoblast proliferation, and a variety of effects on the osteocytes which are still being explored. Thereby, PTH is intimately involved in both the long-termregulation of bone matrix turnover and the short-term regulation of the concentration of calcium in the extracellular fluid. However, much remains to be learned of the biochemical mechanisms involved in the interaction between the hormone and its target cells. Specifically, the biochemistry of the FTH receptor and the significance of intracellular events triggered by hormone-receptor interactions, such as increases in glucose-6-phosphate dehydrogenase activity [103], will deservedly receive attention in the immediate future. The biochemistry of hormone-receptor interactions, incidentally, is of considerable clinical interest, not least because of the ability of non-PTH circulating products to activate the PTH receptor in some cases of malignant hypercalcaemia [ 1041. 235 Further studies on the interaction between bone surface osteocytes and osteoclasts will be crucial to understanding the role of PTH in regulating bone resorption and the rate of initiation of new basic multicellular units. The hypothesis of Rodan & Martin [lo51 may generate a number of detailed variants which will need to be examined critically. Experimental studies have shown that PTH increases the spaces between adjacent osteocytes [ 1061 and this has suggested a mechanism whereby PTH may allow the osteoclasts access to uncovered bone surfaces, leading to the initiation of ,resorption. The thin layer of under-mineralized matrix beneath all surface osteocytes might still inhibit resorption unless first removed by collagenase secreted by cells related to the osteocyte-osteoblast lineage [ 1071. While the rate at which new BMU are initiated determines the rate of bone turnover, it does not in itself determine the rate of bone loss or gain. In a given volume of bone this is determined by the product of the mean loss or gain of bone in each BMU and the rate of initiation of new BMU. In the Haversian systems within the cortex of bone there is, of course, no potential room for expansion; but patterns of both loss and gain may be seen within the three other bony ‘envelopes’: periosteal, endosteal and trabecular [108]. At the BMU level the initial loss of bone is determined by the number of osteoclasts, the rate at which they work individually and the duration of the osteoclastic phase. From use of a stochastic model of BMU dynamics to analyse the effects of daily injections of hPTH 1-34 in patients with osteoporosis, it was concluded that increases in trabecular bone volume of the magnitude observed in some patients could only have been the result of a substantial increase in effective osteoblast lifespan [109], a phenomenon also observed directly in patients treated successfully with sodium fluoride plus calcium [110]. Conversely, before treatment, many similar patients showed evidence of a reduced effective osteoblast lifespan [ 1113 associated with prolonged periods when the associated osteoid was not mineralizing [17].The relationship of parathyroid peptides to the maturation and stabilization of osteoblast populations clearly requires further investigation. The application of new techniques to study the proliferative potential of the osteoblast stem cell and the study of the effects of parathyroid peptides on the intermediary metabolism of bone cells and proteins synthesized by such cells in isolation and in tissue sections should have a profound influence on our future understanding of the regulation of bone in health and disease. J. Reeve and J. M. Zanelli 236 Acknowledgments We thank Maureen Owen, Pierre Meunier and Michael Parfitt for helpful discussions. References 1. Mandl, F. (1926) ThLrapeutischer Versuch bei einem Falle von Ostitis fibrosa generalisata mittels Exstirpation eines Epithelkorperchentumors. Zentralblatrfirr Chirurgie, 5,260-264. 2 . Mandl, F. (1929) Zur Frage der Exstirpation eines Epithelkorpertumors bei der allgemeinen Ostitisfibrosa. Zentralblatt f i r Chirurgie, 28, 1739-1745. 3. Bauer, W., Aub, J.C. & Albright, F. (1929) Studies of calcium and phosphorus metabolism. V. A study of the bone trabeculae as a readily available reserve supply of calcium. Journal of Experimental Medicine, 49, 145-162. 4. Selye, H. (1932) On the stimulation of new bone-formation with parathyroid exfract and irradiated ergosterol. Endocrinology, 16, 547-558. 5. Pugsley, L.1. & Selye, H. (1933) The histological changes in the bone responsible for the action of parathyroid hormone on the calcium metabolism of the rat. Journal of Physiology (London). 79,113-117. 6. Shelling, D.H., Asher, D.E. & Jackson, D.A. (1933) Calcium and phosphorus studies: (vii) the effects of variations in dosage of parathormone and calcium and phosphorus in the diet on the concentrations of calcium and phosphorus in the serum and on the histology and chemical composition of the bones of rats. Bulletin of the Johns Hopkins Hospital, 53, 348-389. 7. Burrows, R.B. (1938) Variations produced in bones of growing rats by parathyroid extracts. American Journal of Anatomy, 62, 237-290. 8. Barnicot, N.A. (1945) Some data on the effect of parathormone on the grey-lethal mouse. JournalofAnatomy, 79,83-93. 9. Jaffe, H.L. (1933) Hyperparathyroidism (von Recklinghausen’s disease of bone). Archives of Pathology, 16.63- 112. 10. Frost, H.M. (1970) Tetracycline-based histological analysis of bone remodelling. Calcified Tissue Research, 3.21 1-237. 11. Owen, M. (1985) Lineage of osteogenic cells and their relationship to the stromal system. In: Bone and Mineral Research Annual3, pp. 1-25. Ed. Peck, W. Excerpta Medica, Amsterdam. 12. Ericsson, J.L.E. (1980) Origin and structure of the osteoclast. In: Mononuclear Phagocyres: Functional Aspects, part 1, p. 203. Ed. Van Furth, R. Martinus Nijhoff, The Hague/Boston/London. 13. Horton, M.A., Rimmer, E.F., Moore, A. & Chambers, T.J. (1985) On the origin of the osteoclast: the cell surface phenotype of rodent osteoclasts. Calcified Tissue International, 37,46-50. 14. Rasmussen, H. & Bordier. P. (1974) In: The Physiological and Cellular Basis of Metabolic Bone Disease. pp. 41-48. Williams and Wilkins, Baltimore. 15. Baron, R., Vignery, A. & Horowitz, M. (1984) Lymphocytes, macrophages and the regulation of bone remodelling. In: Bonc and Mineral Research Annual 2, pp. 175-243. Ed. Peck, W.A. Elsevier, Amsterdam. 16. Eriksen, E.F., Melsen, F. & Mosekilde, L. (1984) Reconstruction of the resorptive site in iliac trabecular bone: a kinetic model for bone resorption in 20 normal individuals. Metabolic Bone Disease and Related Research, 5,235-242. 17. Parfitt, A.M., Mathews, C., Rao, D., Frame, B., Kleerekoper, M. & Villanueva, A.R. (1981) Impaired osteoblast function in metabolic bone disease. In: Osteoporosis - Recent Advances in Pathogenesis and Treatment, pp. 231-330. Ed. DeLuca, H.F. et al. University Park Press, Baltimore. 18. Melsen, F. & Mosekilde, L. (1978) Tetracycline double-labeling of iliac trabecular bone in 41 normal adults. Calcified Tissue Research, 26.99-102. 19. Parfitt, A.M. (1976) The actions of parathyroid hormone on bone: relation to bone remodeling and turnover, calcium homeostasis. and metabolic bone disease. Part I Mechanisms of calcium transfer between blood and bone and their cellular basis: morphological and kinetic approaches to bone turnover. Metabolism, 25,809-844. 20. Parfitt, A.M. (1976) The actions of parathyroid hormone on bone: relation to bone remodeling and turnover, calcium homeostasis, and metabolic bone disease. Part II: PTH and bone cells: bone turnover and plasma calcium regulation. Merabolism, 25, 909-955. 21. Parfitt, A.M. (1976) The actions of parathyroid hormone on bone: relation to bone remodelling and turnover, calcium homeostasis, and metabolic bone disease. Part Ill: PTH and osteoblasts, the relationship between bone turnover and bone loss, and the state of the bones in primary hyperparathyroidism. Metabolism, 25,1033-1069. 22. Parfitt, A.M. (1976) The actions of parathyroid hormone on bone: relation to bone remodeling and turnover, calcium homeostasis, and metabolic bone disease. Part IV: The state of the bones in uremic hyperparathyroidism - the mechanisms of skeletal resistance to PTH in renal failure and pseudohypoparathyroidism and the role of PTH in osteoporosis, osteopetrosis, and osteofluorosis. Metabolism, 25,1157-1188. 23. Parsons, J.A. & Potts, J.T., Jr (1972) Physiology and chemistry of parathyroid hormone. Clinics in Endocrinology and Metabolism, 1,33-78. 24. Barnicot, N.A. (1948) The local action of the parathyroid and other tissues on bone in intracerebral grafts. Journal Of AllalOmJ (London),82,233-248. 25. Chang, H.Y. (1951) Grafts of parathyroid and other tissues to bone. Anaromical Record, 111,23-47. 26. Gaillard, P.J. (1955) Parathyroid gland tissue and bone in vitro (I) ExperimentalCell Research, Suppl. 3,154-169. 27. Gaillard, P.J. (1961) Parathyroid and bone in tissue culture. In: The Parathyroids, DP. 20-45. Ed. GreeD, R.O. & Talmage, R.V. Charles C. ThomaH,Springfield, Ill. . 28. Raisz, L.G. (1963) Stimulation of bone resorption by parathyroid hormone in tissueculture. Nature(London),197, 1015-1017. 29. Raisz. L.G. (1965) Bone resomtion in tissue culture. Factors influencing the response to parahyroid hormone. Journal of Clinicallnvestigation, 44, 103-1 16. 30. Goldhaber, P. (1966) Remodelling of bone in tissue culture. Journalof Dental Research, 45,490-499. 31. Vaes, G. (1968) On the mechanisms of bone resorption. Jourtul O f Cell Bioloby, 39,676-697. 32. Zanelli, J.M., Lea, D.J. & Nisbet, J.A. (1969) A bioassay method in vitro for parathyroid hormone. Journal of Endocrinology, 43, 33-46. 33. Stern, P.H., Orr, M.F. & Brull, E. (1982) Ionophore A23187 pro34. 35. 36. 37. 38. 39. motes osteoclast formation in bone organ culture. Calcified Tissue International, 34, 31-36. Lowik, C.W.G.M., van Leeuwen, J.P.T.M., van der Meer, J.M., van Zeeland, J.K., Scheven, B.A.A. & Herrmann-Erlee, M.P.M. (1986) A two receptor model for the action of parathyroid hormone on osteoblasts: a role for intracellular free calcium and CAMP. Cell Calcium (In press). Marcus, R. & Aurbach, G.D. (1969) Bioassay of parathyroid hormone in virro with a stable preparation of adenylate cyclase from rat kidney. Endocrinology, 85,801-810. Chase, L.R., Fedak, S.A. & Aurbach, G.D. (1966) Activation of skeletal adenyl cyclase by parathyroid hormone in vitro. Endocrinology, 84, 761-768. Crisp, A.J., McGuire-Goldring, M.B. & Goldring, S.R. (1984) A system for culture of human trabecular bone and hormone response profiles of derived cells. British Journal of ExperimentalPathology, 65,645-654. Peck, W.A., Kohler, G. & Burr, S. (1981) Calcium mediated enhancement of the cyclic AMP response in cultured bone cells. Calcified Tissue International, 33,409-416. Nissenson, R.A., Mann, E., Winer, J., Teitelbaurn, A.P. & Arnaud, C.D. (1986) Solubilization of the renal receptor for parathyroid hormone. Journal of Bone and Mineral Research, 1, 160. 40. Herrmann-Erlee, M.P.M., Gaillard, P.J. & Hekkelman, J.W. (1978) Regulation of the response of embryonic bone to PTH 41. 42. 43. 44. 45. 46. and PTH fragments. A morphological and biochemical study. In: Endocrinology of Calcium Metabolism, pp. 253-261. Ed. Copp, D.H. & Talmage, R.V. Excerpta Medica, Amsterdam. Howard, G.A., Botterniller, B.L., Turner, R.T., Rader, J.1. & Baylink, D.J. (1981) Parathyroid hormone stimulates bone formation and resorption in organ culture: evidence for a coupling mechanism. Proceedings of the National Academy of Sciences U.S.A.,78, 3204-3208. Farley, J.R. & Baylink, D.J. (1984) Evidence that skeletal growth factor may be a paracrine effector of bone volume. In: Osteoporosis I, pp. 423-430 (Proceedings of Copenhagen International Symposium on Osteoporosis, 3-8 June 1984). Ed. Christiansen, C., Arnaud, C.D., Nordin, B.E.C., Parfitt, A.M., Peck, W.A. & Riggs, B.L. Department of Clinical Chemistry, Glostrup Hospital, Copenhagen. Bingham, P.J., Brazell, LA. & Owen, M. (1969) The effect of parathyroid extract on cellular activity and plasma calcium levels in vivo. Journal ofEndocrinology, 45,387-410. Chambers, T.J., Revell, P.A., Fuller, K. & Athanosou, N.A. (1984) Resorption of bone by isolated osteoclasts. Journal of Cell Science, 66,383-399. Chambers, T.J., McSheehy, P.M.J., Thomson, B.M. & Fuller, K. (1985) The effect of calcium-regulating hormones and prostaglandins on bone resorption by osteoclasts disaggregated from neonatal rabbit bones. Endocrinology, 116,234-239. Braidman,I.P., Anderson, D.C., Jones,C.J.P. & Weiss, J.B.(1983) Separation of two bone cell populations from foetal rat calvaria and a study of their responses to parathyroid hormone and calcitonin. Journal of Endocrinology, 99, 387-399. Parathyroid hormone and bone 47. Wong, G.L. & Cohn, D.V. (1974) Separation of uarathvroid hormone and calcitonin-sensitive cellc from non-;espo&ve bone cells. Nature(London),252,713-715. 48. Wong, G.L. & Cohn, D.V. (1975) Target cells in bone for parathormone and calcitonin are different: enrichment for each cell type by sequential digestion of mouse calvaria and selective adhesion lo polymeric surfaces. Proceedings of the National AcademyofSciences U.S.A.,72,3167-3171. 49. Luben, R.A., Wong, G.L. & Cohn, D.V. (1976) Biochemical characterization with parathormone and calcitonin of isolated bone cells: provisional identification of osteoclasts and osteoblasts. Endocrinology, 99, 525-534. 50. Friedenstein, A.J., Chailakhyan, R.K., Latsinik, N.V., Panasyuk, A.F. & Keiliss-Borok, I.V. (1974) Stromal cells responsible for transferring the microenvironment of the hemopoietic tissues. Transplantation, 17,331-340. 51. Ashton, B.A., Abdullah, F., Cave, J., Williamson, M., Sykes, B.C., Couch, M. & Poser, J.W. (1985) Characterization of cells with high alkaline phosphatase activity derived from human bone and marrow: Preliminary assessment of their osteogenicity. Bone, 6, 313-320. 52. Ashton, B.A., Allen, T.D., Howlett, C.R., Eaglesom, C.C., Hattori, A. & Owen, M. (1980) Formation of bone and cartilage by marrow stromal cells in diffusion chambers in vivo. Clinical Orthopaedics and Related Research, 151,294-307. 53. Bab, I., Howlett, C.R., Ashton, B.A. & Owen, M.E. (1984)Ultrastructure of bone and cartilage formed in vivo in diffusion chambers. Clinical Orthopaedics and Related Research, 187, 243-254. 54. Friedenstein, A.J. (1980) Stromal mechanisms of bone marrow: cloning in vitro and retransplantation in vivo. In: lmmunobiology of Bone Marrow Transplantation, pp. 19-29. Ed. Thierenfelder, S. Springer-Verlag,Berlin. 55. Aubin, J.E., Heersche, J.N.M., Merrilees, M.J. & Sodek, J.(1982) Isolation of bone cell clones with differences in growth, hormone responses and extracellular matrix proteins. Journal of Cell Biology, 92,452-461. 56. Tyrrell, D.A.J., Mika-Johnson, M. & Chapple, P.J. (1979) Clones of cells from a human embryo lung: their growth and susceptibility to respiratory viruses. Archives of Virology, 6 1 , 6 9 4 5 , 57. Martin, T.J., Ingleton, P.M., Underwood, JE.C., Michelangeli, V.P., Hunt, N.H. & Melick, R.A. (1976) Parathyroid hormoneresponsive adenylate cyclase in induced transplantable osteogenic rat sarcoma. Nature (London),260,436-438. 58. Livesey, S.A., Kemp, B.E., Re, C.A., Partridge, N.C. & Martin, T.J. (1982) Selective hormonal activation of cyclic AMP-dependent protein kinase isohormones in normal and malignant osteoblasts. Journalof BiologicalChemistry, 257,14983-14985. 59. Majeska, R.J., Rodan, S.B. & Rodan, G.A. (1978) Maintenance of parathyroid hormone response in clonal rat osteosarcoma lines. Experimental Cell Research, 11 1,465-468. 60. Rodan, G.A. & Rodan, S.B. (1984) Expression of the osteoblast phenotype. In: Bone and Mineral Research Annual 2, pp. 244-285. Ed. Peck, W.A. Elsevier, Amsterdam. 61. Hamilton, J.A., Lingelbach, S.R., Partridge, N.C. & Martin, T.J. (1984) Stimulation of plasminogen activator in osteoblast-like cells by bone resorbing hormones. Biochemical and Biophysical Research Communications, 122,230-236. 62. Beresford, J.N., Gallagher, J.A., Poser, J.W. & Russell, R.G.G. (1984) Production of osteocalcin by human bone cells in vitro. Effects of 1,25(OH),D,, 24,25(OH),D,, parathyroid hormone and glucocorticoids. Metabolic Bone Disease and Related Research, 5,229-234. 63. Chambers, T.J. (1985)The pathology of the osteoclast. Journal of Clinical Pathology, 38, 241-252. 64. Braidman, I.P., Anderson, D.C. & Robertson, W.R. (1985) Hormone responses mediated via interactions between different cell types, effects of physiological concentrations of parathyroid hormone on cultured bone cells. Journal of Endocrinology, 104 (Suppl.), 47. Abstract 61. 65. Raisz, L.G. & Kraem, B.A. (1983) Regulation of bone formation 1. New England Journal of Medicine, 309, 29-35. 66. Kahn, A.J., Fallon, M.D. & Teitelbaum, S.L. (1984) Structure-function relationships in bone: an examination of events at the cellular level. In: Bone and Mineral Research Annual 2, pp. 125-174. Ed. Peck, W.A. Elsevier, Amsterdam. 67. Charhon, S.A., Edouard, C.M., Arlot, M.E. & Meunier, P.J. (1982) Effects of parathyroid hormone on remodeling of iliac trabecular bone packets in patients with primary hyperparathyroidism. Clinical Orthopaedics and Related Research, 162, 255-263. 68. Peterson, D.R., Heideger, W.J. & Beach, K.W. (1985) Calcium homeostasis: the effect of PTH on bone membrane electrical potential differences. Calcified Tissue International, 37, 307-311. 69. Grubb, S.A., Edwards, G. & Talmage, R.V. (1977) Effect of endogenous and infused parathyroid hormone on plasma con- 237 centrations of recently administered “Ca. Calcified Tissue Research, 24,209-214. 70. Talmage, R.V., Cooper, C.W. & Toverud, S.U. (1983)The physiological significance of calcitonin. In: Bone and Mineral Research Annual I, pp. 74-143. Ed. Peck, W.A. Excerpta Medica, Amsterdam. 71. Parfitt, A.M. (1979) Equilibrium and disequilibrium hypercalcaemia. New light on an old concept. Metabolic Bone Disease and Related Research. 1. 279-293. 72. Parsons, J.A. & Robikbn, C.J (1971) Calcium shift into bone causing transient hypocalcaemia after injection of parathyroid hormone. Nature (London),230,581-582. 73. Parsons, J.A., Reit, B. & Robinson, C.J. (1973) A bioassay for parathyroid hormone using chicks. Endocrinology, 92,454-462. 74. Mazzuoli, G.F., D’Erasmo, E., Scarda, A,, Minisola, S., Mancini, D. & Malaguti Aliberti, L. (1979)Significance of early increase in stable and radioactive plasma calcium after parathyroidectomy in primary hyperparathyroidism. Calcified Tissue International, 29, 185-191. 75. Stevenson, R.W., Parsons, J.A., Podbesek, R.D. & Reeve, J. (1983) Changes in plasma calcium and in radiocalcium kinetics after termination of five-week infusions of synthetic parathyroid peptide in dogs. Acta Endocrinologica, 104,462-467. 76. Atkinson, M.J., Vido, I.,Keck, E. & Hesch, R.-D. (1983)Hepalic osteodystrophy in primary biliary cirrhosis: a possible defect of Kupffer cell mediated cleavage of parathyroid hormone. Clinical Endocrinology, 18.21-28. 77. Atkinson, M.J., Schettler, T., Bodenstein, H. & Hesch, R.-D. (1984) Osteoporosis: a bone turnover defect resulting from an elevated parathyroid hormone concentration within the bone marrow cavity? Klinische Wochenschrift, 62,129-132. 78. Hesp, R., Tellez, M. & Davidson, L. (1986)Trabecular and cortical bone in the radii of female patients with primary hyperparathyroidism. ClinicalScience, 70 (Suppl. 13), 2OP-21P. 79. Parfitt, A.M. (1986) Accelerated cortical bone loss in primary and secondary hyperparathyroidism. In: Bone Fragility in Ortho paedics and Medicine. Ed. Uhthoff, H.K. & ,Jaworski, Z.F.G. Springer Verlag, Berlin, Heidelberg, New York. (In press). 80. Malluche, H.H., Sherman, D., Meyer, W., Ritz, E., Norman, A.W. & Massry, S.G. (1982) Effects of long-term infusion of physiologic doses of 1-34 PTH on bone. American Journal of Physiology, 242 (Renal, Fluid and Electrolyte Physiology, 1l ) , F197-FZ01. 81 Podbesek, R., Edouard, C., Meunier, P.J., Parsons, J.A., Reeve, J., Stevenson, R.W. & Zanelli, J.M. (1983) Effects of two treatment regimes with synthetic human parathyroid hormone fragment on bone formation and the tissue balance of trabecular bone in erevhounds. Endocrinology, 112,1000-1006. 82. Reeve, J., Meunier, P.J., Parsons, J.A., Bernat, M., Bijvoet, O.L.M., Courpron, P., Edouard, C., Klenerman, L., Neer, R.M., Renier, J.C., Slovik, D., Vismans, F.J.F.E. & Potts, J.T., Jr (1980) The anabolic effect of human parathyroid hormone fragment (hPTH 1-34) therapy on trabecular bone in involutional osteoporosis: report of a multi-centre trial. British Medical Journal, 280,1340-1344. 83. Tam, C.S., Heersche, J.N.M., Murray, T.M. & Parsons, J.A. (1982) Parathyroid hormone stimulates the bone apposition rate indeaendentlv of its resorative action: differential effects of intermittdnt and. continuous’ administration. Endocrinology, 506-512. 84. Walker, D.G. (1971) The induction of osteopetrotic changes in hypophysectomized, thyroparathyroidectomized, and intact rats of various ages. Endocrinology, 89, 1389-1406. 85. Kalu, D.N., Doyle, F.H., Pennock, J. & Foster, G.V. (1970) Parathyroid hormone and experimental osteosclerosis. Lancet, i, 1363-1366. 86. Slovik, D.M., Rosenthal, D.I., Doppelt, S.H., Daly, M.A., Murray, J.A. & Neer, R.M. (1985) Reversal of idiopathic osteoporosis by treatment with hPTH (1-34) and 1,25(OH),D,. Clinical Research, 33,444A. 87. Martin, K.J., Hruska, K., Bellorin-Font, E. & Slatopolsky, E. (1981) The peripheral metabolism of parathyroid hormone: activation or inactivation? In: Hormonal Control of Calcium Metabolism, pp. 64-69. Ed. Cohn, D.V., Talmage, R.V. & Matthews, J.L. Excerpta Medica, Amsterdam. 88. ZaneUi, J.M., Lane, E., Kimura, T. & Sakakibara, S . (1985) Biological activities of synthetic human parathyroid hormone 1-84 relative to natural bovine parathyroid hormone in two different in vivo bioassay systems. Endocrinology, 117,1962-1967. 89. Herrmann-Erlee, M.P.M., Nijweide, P.J., Van der Meer, J.M. & Ooms, M.A.C. (1983) Action of bPTH and bPTH fragments on embryonic bone in vitro: dissociation of the cyclic AMP and bone resorbing response. Calcified Tissue International, 35, 70-77. 90. Hefti, E., Trechsel, U., Bonjour, J.-P., Fleisch, H. & Schenk, R. (1982) Increase of whole body calcium and skeletal mass in I . 238 91. 92. 93. Y4. 95. 96. 97. 98. 99. J. Reeve and J. M.Zanelli normal and osteoporotic adult rats treated with parathyroid hormone. Clinical Science, 62, 389-396. Kent, G.N., Loveridge, N., Reeve, J. & Zanelli, J.M. (1985) Pharmacokinetics of synthetic human parathyroid hormone (134) in man measured with cytochemical bioassay and radioimmunoassay. Clinical Science, 68, 171-177. Reeve, J., Tregear, G.W. & Parsons, J A . (1976) Preliminary trial of low doses of human parathyroid hormone 1-34 peptide in treatment of osteoporosis. Calcified Tissue Research, 21 (Suppl.), 469-477. Kent, G.N. & Zanelli, J.M. (1983) Parathyroid hormone. In: C y l o chemical Bioassays: Techniques and Clinical Applications, pp. 255-307. Ed. Chayen, J. & Bitensky, L. Marcel Dekker Inc., New York, Basel/Butterworths, London. Slovik, D.M., Neer, R.M. & Potts, J.T., Jr (1981) Short-term effects of synthetic human parathyroid hormone-( 1-34) administration on bone mineral metabolism in osteoporotic patients. Journalof Clinical Investigation, 68, 1261-1271. Meunier, P.J., S.-Bianchi, G.G., Edouard, C.M., Bernard, J.C., Courpron, P. & Vignon, G.E. (1972) Bony manifestations of thyrotoxicosis. In: Orthopedic Clinics of North America, vol. 3, pp. 745-774. Ed. Hohl, J.C. W.B. Saunders, Philadelphia, London, Toronto. Frost, H.M. (1979) Treatment of osteoporoses by manipulation of coherent bone cell populations. Clinical Orthopaedics and Related Research, 143,227-244. Reeve, J., Podbesek, R.D., Price, T.R., Arlot, M., Bartlett, C., Deacon, A.A., Edouard, C., Green, J.R., Hesp, R., Hulme, P., Katz, D., Tellez, M.. Zanelli, G.D., Zanelli, J.M. & Meunier, P.J. (1984) Studies of a ‘short-cycle’ADFR regime using parathyroid peptide hPTH 1-34 in idiopathic osteoporosis and in a dog model. In: Osteoporosis (Proceedings of the Copenhagen International Symposium, 3-8 June 1984), vol. 2, pp. 567-573. Ed. Christiansen, C., Amaud, C.D., Nordin, B.E.C., Parfitt, A.M., Peck, W.A. & Riggs, B.L. Department of Clinical Chemistry, Glostrup Hospital, Copenhagen. Gallagher, J.C. (1981) Biochemical effects of estrogen and progesterone on calcium metabolism. In: Osteoporosis: Recent Advances in Pathogenesis and Treatment, pp. 231-238. Ed. De Luca, H.F., Frost, H.M., Jee, W.S.S.,Johnston, C.C. & Parfitt, A.M. University Park Press, Baltimore. Mosekilde, L. & Melsen, F. (1978) A tetracycline-based histomorphometric evaluation of bone resorption and bone turnover in hyperthyroidism and hyperparathyroidism. Acta Medica Scandinavica, 204,97-102. 100. Bressot, C., Meunier, P.J., Chapuy, M.C., Lejeune, E., Edouard, C. & Darby, A.J. (1979) Histomorphometric profile, patho- physiology and reversibility of carticosteroid-induced osteoporosis. Metabolic Bone Disease and Related Research, 1, 303-31 1. 101. Mundy, G.R. & Martin, T.J. (1982) The hypercalcemia of malignancy: Pathogenesis and management. Metabolism, 3 1, 1247-1277. 102. Broadus, A.E. & Stewart, A.F. (1983) Humoral hypercalcaemia 103. 104. 105. 106. of malignancy. In: Clinical Disorders of Bone and Mineral Metabolism, pp. 284-287. Ed. Frame, B. & Potts, J.T., Jr. Excerpta Medica. Amsterdam. Dunham, J. & Chayen, J. (1983) An effect of parathyroid hormone on the epiphyseal plate and osteoblasts: studies towards a cytochemical bioassay. Journal of Imrnunoassay, 4,329-338. Mundy, G.R., Ibbotson, K.J., DSouza, S.M., Jacobs, J.W. & Martin, T.J. (1984) The hypercalcaemia of cancer: clinical implications and oathoeenic mechanisms. New Ennland Journal of Medicine, 316, 17r8-1727. Rodan, G.A. & Martin, T.J. (1981) Role of osteoblasts in hormonal control of bone resorption - an hypothesis. Calcified Tissue International, 33, 349-351. Jones, S.J. & Boyde, A. (1976) Experimental study of changes in osteoblastic shape induced by calcitonin and parathyroid extract in an ore.an culture system. Cell and Tissue Research, 169, 449-4651 107. Chambers, T.J. & Fuller, K. (1986) Osteoblasts initiate bone resorption by exposing bone mineral lo osteoclastic contact (Abstract). Bone (In press). 108. Frost, H.M. (1985) The skeletal intermediary organisation: a synthesis. In: Bone and Mineral Research Annual3, pp. 49-108. Ed. Peck, W.A. Elsevier, Amsterdam. 109. Reeve, J. (1985) A stochastic analysis of iliac trabecular bone dynamics. Clinical Orthopaedics and Related Research (In press). 110. Eriksen, E.F., Mosekilde, E.F. & Melsen, F. (1984) Bone balance at the BMU level in 16 osteoporotic patients before and after 5 years treatment with calcium, vitamin D, and sodium fluoride. In: Osteoporosis (Proceedings of the Copenhagen International Symposium, 3-8 June 1984). vol. 1, pp. 493-498. Ed. Christiansen, C., Arnaud, C.D., Nordin, B.E.C., Parfitt, A.M., Peck, W.A. & Riggs, B.L. Department of Clinical Chemistry, Glostrup Hospital, Copenhagen. 111. Arlot, M., Edouard, C., Meunier, P.J., Neer, R.M. & Reeve, J. (1984) Imoaired osteoblast function in osteovorosis: comvarison between chcium balance and dynamic histomorphometry.. British Medicallournal, 289,517-520.