Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
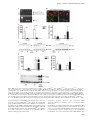
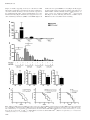
Carcinogenesis vol.32 no.10 pp.1441–1449, 2011 doi:10.1093/carcin/bgr134 Advance Access publication July 18, 2011 Deletion of cyclooxygenase 2 in mouse mammary epithelial cells delays breast cancer onset through augmentation of type 1 immune responses in tumors Nune Markosyan, Edward P.Chen, Victoire N.Ndong, Yubing Yao, Christopher J.Sterner1, Lewis A.Chodosh1, John A.Lawson, Garret A.FitzGerald and Emer M.Smyth Department of Pharmacology, University of Pennsylvania, Institute for Translational Medicine and Therapeutics, Philadelphia, PA 19104, USA and 1 Department of Cancer Biology, University of Pennsylvania, Abramson Family Cancer Research Institute, Philadelphia, PA 19104, USA To whom correspondence should be addressed. University of Pennsylvania, 421 Curie Boulevard, 808 Biomedical research Building II/III, Philadelphia, PA 19104, USA. Tel: þ215 573 2323; Fax: þ215 573 9004; Email: [email protected] Inhibition of cyclooxygenase (COX) 2, which is associated with >40% of breast cancers, decreases the risk of tumorigenesis and breast cancer recurrence. To study the role of COX-2 in breast cancer, we engineered mice that lack selectively mammary epithelial cell (MEC) COX-2 (COX-2 KOMEC). Compared with wild type (WT), MEC from COX-2 KOMEC mice expressed >90% less COX2 messenger RNA (mRNA) and protein and produced 90% less of the dominant pro-oncogenic COX-2 product, prostaglandin (PG) E2. We confirmed COX-2 as the principle source of PGE2 in MEC treated with selective COX-2 and COX-1 inhibitors. Tumors were induced in mice using medroxyprogesterone acetate and 7,12-dimethylbenz[a]anthracene. Breast cancer onset was significantly delayed in COX-2 KOMEC compared with WT (P 5 0.03), equivalent to the delay following systemic COX-2 inhibition with rofecoxib. Compared with WT, COX-2 KOMEC tumors showed increased mRNA for Caspase-3, Ki-67 and common markers for leukocytes (CD45) and macrophages (F4/80). Analysis of multiple markers/ cytokines, namely CD86, inducible nitric oxide synthase (iNOS), interleukin-6, tumor necrosis factor a (TNFa) and Tim-3 indicated a shift toward antitumorigenic type 1 immune responses in COX-2 KOMEC tumors. Immunohistochemical analysis confirmed elevated expression of CD45, F4/80 and CD86 in COX-2 KOMEC tumors. Concordant with a role for COX-2 in restraining M1 macrophage polarization, CD86 and TNFa expression were offset by exogenous PGE2 in bone marrow-derived macrophages polarized in vitro to the M1 phenotype. Our data reveal the importance of epithelial COX-2 in tumor promotion and indicate that deletion of epithelial COX-2 may skew tumor immunity toward type 1 responses, coincident with delayed tumor development. Introduction Cyclooxygenase (COX)-2, the inducible COX isoform, has been convincingly linked with colon, prostate, lung and breast cancer (1,2) across basic, animal and human studies. In breast tumors, increased COX-2 expression correlates with parameters of aggressiveness and poor outcome (3). Inhibition or deletion of COX-2 reduced tumor incidence in animal models of mammary tumorigenesis (4–6), whereas targeted overexpression of COX-2 to the mammary epithelium was sufficient to induce tumor formation (7). Concordantly, preventive effects of nonsteroidal anti-inflammatory drugs, which inhibit both the COX-1 and COX-2 isoforms (8), or COX-2 selective inhibitors (1,9) have been consistently reported. These and other studies have raised interest in Abbreviations: BMDM, bone marrow-derived macrophages; COX, cyclooxygenase; LPS, lipopolysaccharide; MEC, mammary epithelial cell; mmtv, mouse mammary tumor virus; mRNA, messenger RNA; PCR, polymerase chain reaction; PG, prostaglandin; Q-PCR, quantitative polymerase chain reaction; TAM, tumor-associated macrophages; TNFa, tumor necrosis factor a; WT, wild type. targeting COX metabolic pathways, COX-2 in particular, for chemoprevention and/or therapy. Indeed, celecoxib, a selective COX-2 inhibitor, is approved for the prevention and treatment of familial adenomatous polyposis, a condition that frequently proceeds to colon cancer (10). However, clinical use of the COX-2 inhibitors in chemoprevention and/or therapy is limited because of an increased cardiovascular risk associated with this class of drugs (11,12). COX-2 uses arachidonic acid as a substrate to catalyze the production of the prostaglandins (PGs) and thromboxane A2. PGE2, a ubiquitous and well-studied product of COX-2, has proven pro-oncogenic roles in many tissues (1). Thus, PGE2 enhanced intestinal adenoma growth via activation of the Ras–MAPK cascade (13), whereas cell migration was stimulated through activation of the epidermal growth factor receptor (14). PGE2 also promoted survival and growth of human colon cancer cells via induction of the anti-apoptotic protein Bcl-2 (15) and the Gs–axin–ß-catenin signaling axis (16). Recent studies in mice with enhanced mammary COX-2 expression established the role of PGE2 in the ’angiogenic switch’ that is necessary for cancer development, growth and invasion (17). Primary tumor cells are surrounded and strongly influenced by stromal cells, including fibroblasts, adipocytes and immune cells, and a supportive microenvironment appears critical for the tumor to progress toward malignancy (18,19). Environmental cues, tumor-derived stimuli and contact with tumor cells direct stromal cell differentiation and function (20,21). COX-2 contributes to this epithelial-stromal interplay. For example, the pro-tumorigenic effects of fibroblasts and macrophages have been linked to COX-2 upregulation in tumor cells (22,23), whereas COX-2-derived PGE2, in turn, contributed to matrix remodeling and suppressed immune surveillance (24). Tumor-associated immune responses can be generalized to type 1, in which Th1 lymphocytes and M1-polarized macrophages limit tumor progression, and type 2, in which Th 2 lymphocytes and M2 macrophages favor immune escape and disease progression (25). PGE2 has been reported to augment protumorigenic type 2 lymphocyte and myeloid cell functions (26,27). However, the precise contribution of COX-2 to tumor-immune responses within the microenvironment remains ill-defined. Strategies that interrupt global COX-2 function, either via inhibitors or systemic deletion of the COX-2 gene, make it difficult to decipher its role in orchestration of tumor cell’s interaction with its microenvironment. In the present study, we report generation of mice that lack COX-2 only in mammary epithelium. Selective deletion of mammary epithelial COX-2 was sufficient to delay the onset of breast cancer in mice and equivalent in benefit to systemic COX-2 inhibition. This was associated with elevated expression of type 1 inflammatory and immune cell markers/cytokines, indicating a role for epithelial COX-2 in limiting antitumorigenic immune responses in mammary tumors. Materials and methods Mice All procedures involving mice were conducted in accordance with National Institutes of Health regulations and were approved by the Institutional Animal Care and Use Committee of the University of Pennsylvania. Floxed COX-2 mice, generated by flanking the region of the COX-2 gene between introns 5 and 8 with loxP sites (COX-2flox/flox) (28), were backcrossed fully (.9 generations) onto an FVB background (29). COX-2flox/flox mice were crossed with FVB mice expressing Cre-recombinase under control of the mouse mammary tumor virus (mmtv) promoter (Cremmtv), which is used widely to target transgene expression to the mammary epithelial cells (MEC) (29). The resulting mice were termed COX-2 KOMEC. Genotyping Genomic DNA was extracted from mouse tails and used for polymerase chain reaction (PCR). Primers are listed in supplementary Materials and Methods (available at Carcinogenesis online). Ó The Author 2011. Published by Oxford University Press. All rights reserved. For Permissions, please email: [email protected] 1441 N.Markosyan et al. MEC, peritoneal macrophage and bone marrow-derived macrophage isolation and culture Mouse MEC were isolated and cultured according to the protocol from StemCell Technologies (Vancouver, BC Canada). Reagents used were from StemCell Technologies, unless otherwise stated. Cells were plated on 6- or 12-well plates (50 000 or 100 000 cells/well, respectively) in EpiCult-B medium, supplemented with recombinant human epidermal growth factor, recombinant human basic fibroblast growth factor, Heparin and 5% fetal bovine serum; 24 h later, cells were transferred to serum-free EpiCult-B medium, to prevent overgrowth by stromal cells. Cells, at 80% confluency, were or were not treated with 5 lg/ml lipopolysaccharide (LPS; Sigma, St Louis, MO) for 24 h. For inhibitor studies, rofecoxib (COX-2 selective inhibitor, 107 M; Sequoia Research, Berkshire, UK) or FR122047 (COX-1 selective inhibitor, 107 M; Cayman Chemicals, Ann Arbor, MI) were added at the beginning of the LPS treatment. RNA was extracted from cells in 12well plates (see below); 1 ml conditioned medium from each well of the six-well plates was stored at 80°C for prostanoid measurements, and cells were lysed with radio-immunoprecipitation assay buffer (Sigma) for western blotting. Ice-cold sterile phosphate-buffered saline was injected into the peritoneal cavity for peritoneal macrophage collection. Peritoneal fluid was collected through a small midline incision. After centrifugation, cells were resuspended and plated on a 60-mm dish in 3 ml RPMI1640 medium (Invitrogen) with 10% fetal bovine serum, 1% Pen/Strep and 1% L-glutamate. After an overnight incubation at 37°C, the cells were placed in serum-free medium for 24 h and then treated with or without 5 lg/ml LPS for an additional 24 h. Media were harvested for prostanoid measurements. Cells were lysed for RNA and protein extraction. Bone marrow-derived macrophages (BMDM) were isolated as described (30). Femurs and tibias were removed from female mice and flushed with DMEM. Cells were pelleted (300g) and incubated at 37°C for 24 h, in DMEM medium containing 10% fetal bovine serum, 1% Pen/Strep and 1% L-glutamate. L929 cells (American Type Culture Collection, Manassas, VA) were plated in DMEM; 24 h later, L929 cell conditioned medium was collected and mixed 1:5 with DMEM (L-cell conditioned medium). Non-adherent cells from BMDM cultures were collected and plated in L-cell conditioned medium. Purity (99%) was verified by flow cytometry for F4/80 and CD11b (data not shown). Once confluent, conditioned media were collected and BMDM were detached using 10 mM lidocaine in DMEM and lysed for RNA extraction. Conditions for polarization of BMDM are described below. Measurements of eicosanoids PGs and their metabolites were measured by ultra high-pressure liquid chromatography/tandem mass spectrometry, using negative electrospray ionization and multiple reaction monitoring (MRM) techniques, as described previously (31). Additional details are given in the supplementary Materials and Methods (available at Carcinogenesis online). Real-time RT-PCR Total RNA from MEC, macrophages and tumors was isolated (RNeasy; Qiagen, Germantown, MD) and reverse transcribed (TaqMan Reverse Transcriptase; Applied Biosystems, Carlsbad, CA), according to the manufacturer’s instructions. Real-time quantitative polymerase chain reaction (Q-PCR) of all genes, including 18S ribosomal RNA, was performed using inventoried gene expression assays and TaqMan Universal PCR Master Mix from Applied Biosystems. PCR products were detected in ABI-PRISM 7900 sequence detection systems (Applied Biosystems). Results were analyzed using the comparative Ct method (32) and normalized to 18S RNA. Western blotting MEC and macrophages were lysed and homogenized in radio-immunoprecipitation assay buffer. Proteins were quantified (Pierce BCA Assay; Thermo Scientific, Rockford, IL), resolved by SDS-PAGE (4–12% Bis-Tris gel; Invitrogen, Carlsbad, CA) and electrophoretically transferred to nitrocellulose membranes. After blocking, the membranes were incubated with primary antibodies followed by horseradish peroxidase-conjugated secondary antibodies. Immune complexes were detected by enhanced chemiluminescence (GE Healthcare, Piscataway, NJ). Details of the antibodies used are given in the supplementary Methods (available at Carcinogenesis online). Tumor induction Subcutaneous inter-scapular implants of medroxyprogesterone pellets (75 mg/ pellet, 21 day release; Innovative Research of America, Sarasota, FL) were administered to 6-week-old virgin female mice. Starting at 9 weeks, 1 mg/100 ll of 7,12-dimethylbenz[a]anthracene (Sigma) in sesame oil was given, via gavage, weekly for 4 weeks. This regime causes predominantly mammary tumors (33). Mice were fed either with regular chow, or starting from the implant date, diets (Harlan Laboratories, Inc., Madison, WI) containing 10 mg/kg/day rofecoxib or 3 mg/kg/day SC560 (COX-1 selective inhibitor; Tocris Biosciences, Ellisville, MO). Mice were checked weekly and considered tumor bearing if a palpable mammary mass persisted for .1 week. 1442 Immunocytochemistry and immunohistochemistry MEC cultured in Lab-Tek chamber slides (Nalge Nunc International, Naperville, IL) were fixed with 10% buffered formalin, blocked with 3% donkey serum (Jackson Immunoresearch, West Grove, PA) and incubated overnight with primary antibodies followed by a fluorophore-conjugated secondary antibody (as detailed in supplementary Materials and Methods, available at Carcinogenesis online). Cells were visualized by fluorescent microscopy. For immunohistochemistry, after deparaffinization and rehydration, endogenous peroxidase was blocked with 3% hydrogen peroxide, on 5-lm thick tumor sections. Heat-induced epitope retrieval was performed with 1 mM EDTA. Primary antibody treatments were for 1 h at 37°C, followed by immune complex visualization using the Polink-2 HRP Plus AEC System (Golden Bridge International, Inc., Mukillteo, WA). Additional details are given in supplementary Materials (available at Carcinogenesis online). BMDM polarization BMDM were plated (0.5 106 cells/well) in L-cell conditioned medium, which was replaced, 24 h later, with BMDM medium. After a further 24 h, cells received either vehicle or M1 polarizing mix of 100 ng/ml LPS (Sigma) and 20 ng/ml interferon c (Peprotech, Rocky Hill, NJ) with or without 10, 250 and 1000 nM PGE2 (Cayman Chemicals). Supernatants were removed, 18 h later, and cells were lysed for RNA isolation (see above). Tumor necrosis factor a (TNFa) in supernatants was measured by ELISA (R&D Systems, Minneapolis, MN). Statistical analysis Statistical analyses were performed using Prism (GraphPad Software, Inc., La Jolla, CA). As appropriate, comparisons were made using Mann–Whitney test, log-rank analysis, Student’s t-test, or, for multiple group comparisons, analysis of variance followed by Dunnet’s multiple comparison test. Results Characterization of COX-2 KOMEC mice Mouse genotypes—loxP null (COX-2þ/þ), heterozygous (COX-2flox/þ) or homozygous (COX-2flox/flox), and Cremmtv positive or negative—were determined by PCR (Figure 1A). COX-2flox/flox mice lacking Cre expression (Figure1A, lane 4) were considered wild type (WT), whereas COX-2flox/flox mice positive for Cremmtv (Figure 1A, lane 3) were considered COX-2 KOMEC. We estimated that .95% of cells cultured from the single-cell suspensions obtained from the mammary glands of WT and COX-2 KOMEC mice expressed either a luminal epithelial cell marker CK18 or a basal/myoepithelial cell marker CK14, or both, (Figure 1B) and were negative for a fibroblast marker vimentin (supplementary Figure 1 is available at Carcinogenesis online), confirming their predominantly epithelial identity. COX-2 expression in cultured MEC appeared to be constitutive and marginally responsive to LPS induction (Figure 1C). This was in contrast to peritoneal macrophages in which COX-2 expression was strongly induced by LPS treatment (Figure 1E). In MEC from COX-2 KOMEC mice, COX-2 messenger RNA (mRNA) and protein expression were dramatically decreased under vehicle or LPS-treated conditions (Figure 1C and D, left panels). COX-1 mRNA and protein were unaltered (Figure 1C and D, right panels). Importantly, COX-2 and COX-1 expression was not different between peritoneal macrophages obtained from WT and COX-2 KOMEC mice, with or without LPS (Figure 1E). COX-2 protein in peritoneal macrophages similarly was not affected by COX-2 deletion in MEC (Figure 1F). These data confirmed selective loss of COX-2 in MEC. Genomic or pharmacological inhibition of PGE2 production delays mammary tumor onset PGE2 in conditioned media from vehicle or LPS-treated WT MEC exceeded other COX-2 products by 100-fold (Figure 2A). Genomic deletion of COX-2 in MEC led to a 90% reduction in PGE2 production (Figure 2B), despite the presence of COX-1 (Figure 1D). Similarly, treatment with COX-2 selective inhibitor rofecoxib markedly decreased PGE2 production by untreated or LPS-treated MEC (Figure 2B). In contrast, the COX-1 selective inhibitor FR122047 did not affect PGE2 production, whether alone or in combination with rofecoxib, in MEC from WT or COX-2KOMEC mice. Together these data indicate that COX-1 does not contribute to PGE2 generation in MEC. Systemic COX function, as indicated by urinary prostanoid metabolite levels, was unaltered in COX-2 KOMEC mice compared with WT (Figure 2C). Deletion of mammary epithelial COX-2 delays tumors Fig. 1. MEC-specific COX-2 gene deletion. (A) PCR detection of COX-2þ/þ (lane 1), COX-2flox/þ (lane 2), COX-2flox/flox (lanes 3 and 4), Cremmtv/ (lower panel, lanes 1 and 4) and Cre mmtvþ/ (lower panel, lanes 2 and 3) in tail DNA. (B) Immunocytochemistry for CK14 (myoepithelial marker, red) and CK18 (luminal epithelial marker, green) in MEC isolated from WT and COX-2 KOMEC (KO) mice. Background staining in the absence of primary antibody is shown in KO cells (inset) and was similarly negative in WT cells. Nuclei are stained blue with DAPI. Data are representative of similar results from n 5 3. (C) Real-time Q-PCR quantification of COX-2 (left panel) and COX-1 (right panel) mRNA expression in untreated and LPS-treated MEC isolated from WT or COX-2 KOMEC (KO) mice (n 5 6). (D) COX-2 protein in untreated or LPS-treated MEC (left panel) or COX-1 protein in LPS-treated MEC (right panel) from WT and COX-2 KOMEC (KO) mice. (E) COX-2 (left panel) and COX-1 (right panel) mRNA in untreated and LPS-treated peritoneal macrophages from WT and COX-2 KOMEC (KO) mice (n 5 5–6). (F) COX-2 protein in LPS-treated peritoneal macrophages from WT and COX-2 KOMEC (KO) mice. In (C) and (E) data are mean ± SE. P , 0.005. Despite unchanged systemic PGE2 levels, mammary tumor onset was delayed by 8 weeks in COX-2 KOMEC mice (Figure 2D-1). In the WT group, treatment with the COX-2 selective inhibitor rofecoxib, but not the COX-1 selective inhibitor SC560, similarly delayed tumorigenesis (Figure 2D-2). Pharmacological inhibition of either COX isoform did not modify the delayed tumor onset in COX-2 KOMEC mice (Figure 2D-3). These data indicate a dominant role for epithelial COX-2 in mammary tumorigenesis in this model. Proliferative, apoptotic, angiogenic and extracellular matrix remodeling pathways in mammary tumors from WT and COX-2 KOMEC mice To identify the potential mechanism through which COX-2 in MEC promotes mammary tumorigenesis, we next examined expression of a range of genes in WT and COX-2 KOMEC tumors. Messenger RNA levels for both the apoptotic enzyme Caspase 3 and a proliferation marker, Ki67, were increased in tumors from COX-2 KOMEC mice 1443 N.Markosyan et al. (Figure 3A and B), suggesting an increase in cell turnover in KO tumors but not indicating a mechanism for the delay in tumorigenesis in these animals. No changes were detected in the expression of the endothelial marker CD31, either by Q-PCR (Figure 3C) or immunohistochemistry (Figure 5K and L), two angiogenic factors vascular endothelial growth factor (VEGF) A and VEGFC (Figure 3D and E) or their receptors VEGFR 1, 2 and 3 (Figure 3F–H), suggesting that modified angiogenesis did not underlie reduced disease in COX-2 KOMEC animals. Similarly, matrix metalloproteinase enzymes 2 and 9 were unaltered in COX-2 KOMEC compared with WT (Figure 3I and J), indicating that deletion of COX-2 only in MEC did not alter extracellular matrix remodeling. Fig. 2. Inhibition of PGE2 production in MEC delays tumor onset in mice. (A) Prostanoids in conditioned media from cultured MEC treated with or without LPS (n 5 6). (B) PGE2 in conditioned media from MEC obtained from WT and COX-2 KOMEC (KO) mice and incubated with either rofecoxib, or FR12207, or both, with or without LPS (n 5 4–6). (C) Urinary prostanoid metabolites measured in WT and COX-2 KOMEC (KO) disease-free 6-week-old female mice (n 5 10–12). For (A–C), data are mean ± SE, P , 0.05, P , 0.0002. (D) Percent of tumor-free WT and COX-2 KOMEC (KO) mice on regular chow (1, n 5 24–30) or chow containing rofecoxib (2, n 5 17) or SC560 (3, n 5 13–14). 1444 Deletion of mammary epithelial COX-2 delays tumors COX-2 deletion in MEC modifies the tumor-immune microenvironment We observed, by Q-PCR and immunohistochemistry, significant increases in expression of a common leukocyte marker, CD45 (Figures 4A-1 and 5E and F), and a common macrophage marker, F4/80 (Figures 4A-2 and Fig. 5G and H) in COX-2 KOMEC tumors compared with WT. Macrophages can be anti- (M1 phenotype) or pro- (M2 phenotype) tumorigenic, with tumor-associated macrophages (TAM) displaying predominantly M2 characteristics; thus, we examined a range of phenotypic macrophage markers in WT and COX-2 KOMEC tumors. Expression of M2-type macrophage markers Scaf1 (Figure 4B-1) and Arginase-1 (Figure 4B-2), as well as the M2-type cytokine transforming growth factor b (Figure 4B-3), were not different between WT and COX-2 KOMEC tumors. Another M2-type macrophage cytokine, interleukin-10, was low/undetectable in tumors from both animal groups. In contrast, expression of the M1-type marker CD86 was increased in COX-2 KOMEC tumors as assessed by Q-PCR (Figure 4C-1) and immunohistochemistry (Figure 5I and J). Similarly, the M1 macrophage cytokine TNFa was significantly higher in COX-2 KOMEC tumors (Figure 4C-2). Two additional characteristic M1 macrophage-associated genes, inducible nitric oxide synthase and interleukin-6, were not significantly altered (Figure 4C-3 and 4C-4) although interleukin-6 correlated strongly with CD86 or F4/80 in COX-2 KOMEC, but not WT, animals (supplementary Figure 2 is available at Carcinogenesis online). Taken together, these data are consistent with polarization of macrophages associated with COX-2 KOMEC tumors toward the antitumorigenic M1 phenotype. We also examined markers for other tumor-associated immune cells. Significantly higher levels of CD2, a shared marker for T-lymphocytes and natural killer (NK) cells, were evident in COX-2 KOMEC tumors (Figure 4D-1). No significant changes were seen in expression of the NK-specific marker CD49b (Figure 4D-2), the regulatory T cell marker CD25 (Figure 4D-3) or the cytotoxic T cell marker CD8 (Figure 4D-4). In contrast, CD4 was increased in COX-2 KOMEC tumors (Figure 4E-1), indicating increased presence of CD4expressing cells. Among CD4þ cells, Th1 cells are known to direct macrophage polarization toward M1 (34) leading us to speculate that the apparent increase in M1-type responses in COX-2 KOMEC tumors could be secondary to the influence of CD4þ Th1-derived mediators. Indeed, although expression of Th1 markers, TIM3 and TRANCE (35,36) was not significantly different in tumors from COX-2 KOMEC mice compared with WT (Figure 4E-2 and E-3), TIM-3 levels showed almost perfect correlation with both F4/80 and CD86 in COX2MECKO, but not WT, tumors (Figure 4F). Together, these data are consistent with a shift toward type 1 innate and adaptive immune responses in COX-2MECKO tumors. PGE2 limits in vitro M1 Polarization of BMDM Our study suggests that delayed tumorigenesis in COX-2MECKO mice is at least partially due to interruption of the limit placed by COX-2derived PGE2 on type 1 macrophage polarization. We examined whether PGE2 modifies M1 polarization of isolated macrophages. As expected, compared with non-polarized cells, significant increases in mRNA for CD86 and TNFa were evident in M1-polarized BMDM (Figure 6A). PGE2 did not significantly alter M1 markers in nonpolarized cells (supplementary Figure 3 is available at Carcinogenesis online). The M2 marker Arginase-1 expression was at or below the detection level in control and PGE2-treated non-polarized BMDM. However, concordant with our in vivo data, when PGE2 was included at the beginning of the polarization procedure, the M1-associated CD86 and TNFa mRNA expression, as well as TNFa protein secretion, were substantially reduced in dose-dependent manner consistent with a PGE2-mediated restraint of M1 polarization (Figure 6B and C). Discussion Tumorigenesis involves a complex and intricate interplay between tumor and stromal cells (21). Although multiple pro-tumorigenic events are associated with COX-2 (2,37), the contribution of COX2-derived prostanoids to autocrine and paracrine signaling in tumor cells and the surrounding microenvironment are poorly understood. We used a novel model of COX-2 deletion in mammary epithelium to examine the role of COX-2 in epithelial-stromal interplay and its consequence for mammary cancer. We found that deletion of MEC COX-2 induced a substantial delay in tumorigenesis that was equivalent to systemic inhibition of the COX-2 enzyme. We detected no change in proliferative, apoptotic, angiogenic or matrix remodeling pathways to explain the reduced disease in COX-2 KOMEC mice. Fig. 3 Expression of genes associated with proliferation, apoptosis, angiogenesis and matrix remodeling. Real-time Q-PCR quantification of mRNA for all genes are relative to 18S in tumors harvested from WT and COX-2 KOMEC (KO) mice. Data are mean ± SE; n 5 15–16, P , 0.01. 1445 N.Markosyan et al. Fig. 4. Increased association of cells expressing M1-type macrophage markers and cytokines with tumors from COX-2 KOMEC mice. (A–E) Real-time Q-PCR quantification of mRNA for all genes is relative to 18S mRNA. Data presented are mean ± SE; n 5 15–16, P , 0.05. (F) Correlations of TIM3 mRNA expression with F4/80 and CD86 genes in COX-2 KOMEC (KO) and WT tumors (n 5 14). Rather, we observed an increase in markers/cytokines for antitumorigenic M1 macrophages, and possibly Th1 lymphocytes, in COX-2 KOMEC tumors suggesting augmentation of type 1 polarized inflammatory/immune cells in the absence of epithelial COX-2. Consistent with this hypothesis, PGE2, the dominant MEC COX-2-derived prostanoid, restrained M1 polarization of BMDM in vitro. Thus, we conclude that deletion of COX-2 in mouse MECs delayed tumor onset, at least in part, by shifting the tumor inflammatory/immune microenvi- 1446 ronment toward antitumorigenic type 1 responses. We propose a role for epithelial COX-2-derived prostanoids in modulating the tumorimmune microenvironment to favor tumor growth and progression. Multiple prostanoids can be generated via COX-2, and each has been implicated in tumorigenesis. PGE2 is the dominant pro-tumorigenic prostanoid (2,14) although pro-tumorigenic effects of thromboxane A2 have been reported (38). PGI2 and PGD2 are generally considered antitumorigenic (38,39), whereas the role of PGF2a in cancer remains Deletion of mammary epithelial COX-2 delays tumors Fig. 5. COX-2 KOMEC tumors show more M1 macrophage infiltration by immunohistochemistry. Tumors, originated from luminal epithelial cells and harvested from WT and COX-2 KOMEC (KO) mice, were selected based on hematoxylin and eosin staining and immune staining for myoepithelial (red staining for CK14, A and B) and luminal epithelial (red staining for CK18, C and D) cells. Red-brown color represents specific staining for CD45 (E and F), F4/80 (G and H), CD86 (I and J) and CD31 (K and L), in tumors from WT (left column) and COX-2 KOMEC (KO, right column) animals. In panels (E–J), areas within dashed squares are shown as insets with arrowheads, indicating positively stained cells. The scale bar in all images indicates 40 magnification. Images are representative of at least n 5 3. poorly examined (40). In cultured MEC, PGE2 is the most abundant COX-2-derived mediator (Figure 2A), with little or no contribution by COX-1 (Figure 2B). It is highly likely, therefore, that loss of COX2-derived PGE2 in MEC underlies the delayed disease we observed. Deletion of COX-2 in MEC was equivalent to systemic COX-2 inhibition in delaying carcinogen-induced mammary tumorigenesis (Figure 2D). This is perhaps surprising given the theoretical advantage afforded by depression of PGE2 generation not only in MEC but also in surrounding stromal cells in rofecoxib-treated animals. Further, reported COX-independent anticarcinogenic effects of rofecoxib (37) would be expected in drug-treated, but not COX-2 KOMEC, mice. The degree to which COX-2 pathways in stromal cells, or COX-independent events, contribute to mammary tumor progression has yet to be elucidated although the pro-tumorigenic effects of fibroblasts (22) and macrophages (23) may be realized through induction of epithelial cell COX-2. The equivalence of mammary epithelial COX-2 deletion with systemic COX-2 inhibition indicates a dominant role of COX-2 in the epithelial cell itself in tumor promotion. The pro-tumorigenic effects of PGE2, which include increased cell proliferation, cell survival, angiogenesis and decreased apoptosis, are well described (2,37). In our COX-2 KOMEC mice, mRNA for the proliferation marker Ki67 and apoptotic enzyme Caspase-3 were both upregulated, indicating increased cell turnover in tumors lacking epithelial COX-2. We sought to determine, by immunohistochemistry, whether epithelial or specific stromal cells were undergoing proliferation or apoptosis. However, Ki67 and Caspase-3 protein localization and expression intensity were very variable across WT and KO tumors, preventing definitive conclusions. Given the established pro-angiogenic role of COX-2, we were surprised to observe similar expression of genes for angiogenic factors and their receptors in WT and COX-2 KOMEC tumors. It may be that mRNA quantitation in heterogeneous tumor samples did not reveal localized changes in angiogenesis. We do not believe this to be the case, however, because no difference was observed between WT and COX-2 KOMEC tumor levels of CD31, an endothelial marker, by PCR (Figure 3C) or immunohistochemistry (Figure 5K and L). Similarly, expression of extracellular matrix remodeling enzyme matrix metalloproteinase 2, regulated by COX-2 in breast cancer (41), was not different between WT and COX-2 KOMEC tumors (Figure 3I). These data imply that non-epithelial cell COX-2 in COX-2 KOMEC tumors was sufficient to maintain angiogenesis and matrix remodeling at levels similar to WT tumors. Further examination revealed an apparent shift in the immune microenvironment in COX-2 KOMEC tumors. Increased expression of mRNA for the common leukocyte marker CD45, the macrophage marker F4/80, the T cell markers CD2 and CD4 and the M1 macrophage marker CD86 and cytokine TNFa, was evident in COX-2 KOMEC tumors (Figure 4A1, -2, C-1, -2, D-1 and E-1). Elevated CD45, F4/80 and CD86 in the KO tumors were also evident by immunohistochemistry (Figure 5E-J). Confirmation of the M1 versus M2 status of TAM, as well as phenotypic analysis of other tumor associated immune cells by flow cytometry, was not possible in this carcinogen-induced model of mammary tumorigenesis because of limitations in the size and number of tumors obtained and the cellular yield per tumor. However, our data from Q-PCR and immunohistochemistry were consistent with the augmented presence of M1-type macrophages in COX-2 KOMEC tumors. We verified a PGE2dependent restraint on in vitro M1 polarization of BMDM, consistent with the lower levels of M1 markers/cytokines in WT versus COX2MECKO tumors. One putative source of a principle M1 polarizing cytokine, interferon c, is the pool of CD4þ Th1 cells (34,42). In the present study, COX-2 KOMEC tumors also showed increased expression of T cell markers CD2 and CD4, whereas TIM3, a Th1 marker, trended toward elevation (P 5 0.062). It may be that a slight increase in the Th1 cell population in COX-2 KOMEC tumors was sufficient to shift TAM polarization to the M1 phenotype, although this remains to be verified experimentally. Alternatively, M1 polarization of macrophages in COX2 KOMEC may be Th1 independent, relying instead on mediators elaborated from other cells. A permissive microenvironment is necessary for the tumors to progress fully to the malignancy (43). The tumor microenvironment consists of many cells and the composition varies depending on the tumor type. Macrophages populate the microenvironment of most if not all tumors. Independent studies have demonstrated a correlation of TAM with poor prognosis, particularly when associated with high microvascular density (44). Further, the local tumor microenvironment may educate macrophages to perform non-immune, trophic functions similar to the 1447 N.Markosyan et al. Fig. 6. PGE2 limits M1 polarization of BMDM. CD86 and TNFa mRNA expression change, measured by real-time Q-PCR in (A) M1-polarized BMDM (M1) versus non-polarized (control—ctrl) or (B) M1-polarized BMDM treated with 10, 250 and 1000 nM PGE2 versus untreated (0 nM), M1-polarized BMDM. (C) TNFa protein measured by ELISA in conditioned media of non-polarized and untreated (control—ctrl), M1-polarized and untreated (0 nM PGE2) and M1polarized and PGE2-treated (10, 250 and 1000 nM PGE2) BMDM. For all panels, data are mean ± SE, n 5 5, P , 0.05, P , 0.005, P , 0.0005. promotion of epithelial outgrowth and invasion during development (21,45). It appears that once they have infiltrated the tumor, the macrophage phenotype is switched from an activated, immunological M1 type to an alternatively activated M2 type that promotes tumor progression (46). However, clinical studies have shown that, in some cases, elevated macrophage infiltration is associated with a beneficial outcome. Thus, resident and newly recruited macrophages displaying M1 characteristics limited the development of peritoneal metastases from colon carcinoma (47). Similarly, in patients with resected nonsmall cell lung carcinoma, tumor islet macrophage infiltration was identified as a strong favorable independent prognostic marker for survival (48). Further, it was shown that antigen-presenting and cytotoxic functions of M1-polarized macrophages can destroy tumor cells (49). The tumor milieu can direct macrophage polarization to hinder tumor growth and progression. Indeed, in the PyMT mouse model of breast cancer, augmented type 1 polarization of both Th cells and TAM was associated with reduced tumor invasiveness and limited metastasis of the primary disease (42). Our data suggest a putative novel role for COX-2 in contributing to macrophage phenotype in mammary tumors. We believe that MEC-derived COX-2 products, most probably PGE2, restrain M1 polarization of macrophages in tumors, limiting their antitumorigenic functions and promoting tumor progression. It is worth noting that macrophages, whose COX-2 expression was strongly inducible by LPS (Figure 1E), may contribute significantly to tumor microenvironmental PGE2 levels with unexplored paracrine and autocrine effects. The contribution of macrophage COX-2 to mammary tumorigenesis is currently under investigation in our laboratory. The mechanism through which mammary epithelial COX-2derived products impact immune cell polarization in mammary tumors is not clear. There is evidence that prostanoids can modulate T cell and macrophage differentiation and polarization (27,50,51). Our data in isolated BMDM argue for a direct effect of PGE2 on macrophages although this remains to be verified in TAM. Our current findings support the concept that the COX-2/PGE2 biosynthetic pathway may provide novel targets for therapeutic modulation of the tumor-immune microenvironment. Further, in our head-to-head comparison of mammary epithelial versus systemic interruption of COX-2 1448 activity, similar benefit was seen using the targeted approach. Refined approaches to inhibit specific functions of COX-2 in the tumor microenvironment may yield beneficial antitumorigenic effects while avoiding deleterious systemic events that limit clinical use of systemic COX-2 inhibition in cancer prevention or treatment. Supplementary material Supplementary Figures 1–3 and other Materials and Methods can be found at http://carcin.oxfordjournals.org/ Funding American Cancer Society [RSG-08-024-01-CNE to E.M.S.]. Acknowledgements The assistance of Dr Robert Cardiff, UC Davis, with immunohistochemistry is gratefully acknowledged. We also thank Ms Jenny Bruce for her technical assistance. Conflict of Interest Statement: None declared. References 1. Harris,R.E. (2009) Cyclooxygenase-2 (cox-2) blockade in the chemoprevention of cancers of the colon, breast, prostate, and lung. Inflammopharmacology, 17, 55–67. 2. Howe,L.R. (2007) Inflammation and breast cancer. Cyclooxygenase/prostaglandin signaling and breast cancer. Breast Cancer Res., 9, 210. 3. Arun,B. et al. (2004) The role of COX-2 inhibition in breast cancer treatment and prevention. Semin. Oncol., 31, 22–9. 4. Howe,L.R. et al. (2002) Celecoxib, a selective cyclooxygenase 2 inhibitor, protects against human epidermal growth factor receptor 2 (HER-2)/neuinduced breast cancer. Cancer Res., 62, 5405–5407. 5. Lanza-Jacoby,S. et al. (2003) The cyclooxygenase-2 inhibitor, celecoxib, prevents the development of mammary tumors in Her-2/neu mice. Cancer Epidemiol. Biomarkers Prev., 12, 1486–1491. Deletion of mammary epithelial COX-2 delays tumors 6. Howe,L. et al. (2005) HER2/neu-induced mammary tumorigenesis and angiogenesis are reduced in cyclooxygenase-2 knockout mice. Cancer Res., 65, 10113–10119. 7. Liu,C.H. et al. (2001) Overexpression of cyclooxygenase-2 is sufficient to induce tumorigenesis in transgenic mice. J. Biol. Chem., 276, 18563– 18569. 8. Cuzick,J. et al. (2009) Aspirin and non-steroidal anti-inflammatory drugs for cancer prevention: an international consensus statement. Lancet Oncol., 10, 501–507. 9. Harris,R. et al. (2006) Reduction in the risk of human breast cancer by selective cyclooxygenase-2 (COX-2) inhibitors. BMC Cancer, 6, 27. 10. Solomon,S. et al. (2005) Cardiovascular risk associated with celecoxib in a clinical trial for colorectal adenoma prevention. N. Engl. J. Med., 352, 1071–80. 11. Solomon,S. et al. (2006) Effect of celecoxib on cardiovascular events and blood pressure in two trials for the prevention of colorectal adenomas. Circulation, 114, 1028–1035. 12. Grosser,T. et al. (2010) Emotion recollected in tranquility: lessons learned from the COX-2 saga. Annu. Rev. Med., 61, 17–33. 13. Wang,D. et al. (2005) Prostaglandin E2 enhances intestinal adenoma growth via activation of the Ras-mitogen-activated protein kinase cascade. Cancer Res., 65, 1822–1829. 14. Buchanan,F.G. et al. (2003) Prostaglandin E2 regulates cell migration via the intracellular activation of the epidermal growth factor receptor. J. Biol. Chem., 278, 35451–35457. 15. Sheng,H. et al. (1998) Modulation of apoptosis and Bcl-2 expression by prostaglandin E2 in human colon cancer cells. Cancer Res., 58, 362– 366. 16. Castellone,M. et al. (2005) Prostaglandin E2 promotes colon cancer cell growth through a Gs-axin-beta-catenin signaling axis. Science, 310, 1504– 1510. 17. Chang,S. et al. (2004) Role of prostaglandin E2-dependent angiogenic switch in cyclooxygenase 2-induced breast cancer progression. Proc. Natl Acad. Sci. USA, 101, 591–596. 18. Joyce,J.A. et al. (2009) Microenvironmental regulation of metastasis. Nat. Rev. Cancer, 9, 239–252. 19. Hanahan,D. et al. (2011) Hallmarks of cancer: the next generation. Cell, 144, 646–674. 20. Erez,N. et al. (2010) Cancer-associated fibroblasts are activated in incipient neoplasia to orchestrate tumor-promoting inflammation in an NF-kappaBdependent manner. Cancer Cell, 17, 135–147. 21. Pollard,J. (2004) Tumour-educated macrophages promote tumour progression and metastasis. Nat. Rev. Cancer, 4, 71–78. 22. Hu,M. et al. (2009) Role of COX-2 in epithelial-stromal cell interactions and progression of ductal carcinoma in situ of the breast. Proc. Natl Acad. Sci. USA, 106, 3372–3377. 23. Hou,Z. et al. (2011) Macrophages induce COX-2 expression in breast cancer cells: role of IL-1{beta} auto-amplification. Carcinogenesis. 24. Okano,M. et al. (2006) E prostanoid 2 (EP2)/EP4-mediated suppression of antigen-specific human T-cell responses by prostaglandin E2. Immunology, 118, 343–352. 25. DeNardo,D.G. et al. (2010) Interactions between lymphocytes and myeloid cells regulate pro- versus anti-tumor immunity. Cancer Metastasis Rev., 29, 309–316. 26. Wang,D. et al. (2010) Eicosanoids and cancer. Nat. Rev. Cancer, 10, 181– 193. 27. Eruslanov,E. et al. (2010) Pivotal advance: tumor-mediated induction of myeloid-derived suppressor cells and M2-polarized macrophages by altering intracellular PGE2 catabolism in myeloid cells. J. Leukoc. Biol., 88, 839–848. 28. Wang,D. et al. (2009) Cardiomyocyte cyclooxygenase-2 influences cardiac rhythm and function. Proc. Natl Acad. Sci. USA, 106, 7548–7552. 29. Narko,K. et al. (2005) COX-2 inhibitors and genetic background reduce mammary tumorigenesis in cyclooxygenase-2 transgenic mice. Prostaglandins Other Lipid Mediat., 76, 86–94. 30. Sinha,P. et al. (2005) Reduction of myeloid-derived suppressor cells and induction of M1 macrophages facilitate the rejection of established metastatic disease. J. Immunol., 174, 636–645. 31. Song,W. et al. (2007) Noninvasive assessment of the role of cyclooxygenases in cardiovascular health: a detailed HPLC/MS/MS method. Methods Enzymol., 433, 51–72. 32. Bookout,A. et al. (2003) Quantitative real-time PCR protocol for analysis of nuclear receptor signaling pathways. Nucl. Recept. Signal, 1, e012. 33. Yan,D. et al. (2004) A delayed chemically induced tumorigenesis in Brca2 mutant mice. Oncogene, 23, 1896–1901. 34. Martinez,F.O. et al. (2008) Macrophage activation and polarization. Front. Biosci., 13, 453–461. 35. Nakayama,M. et al. (2009) Tim-3 mediates phagocytosis of apoptotic cells and cross-presentation. Blood, 113, 3821–3830. 36. Nakae,S. et al. (2007) Phenotypic differences between Th1 and Th17 cells and negative regulation of Th1 cell differentiation by IL-17. J. Leukoc. Biol., 81, 1258–1268. 37. Grösch,S. et al. (2006) Cyclooxygenase-2 (COX-2)-independent anticarcinogenic effects of selective COX-2 inhibitors. J. Natl. Cancer Inst., 98, 736–747. 38. Pradono,P. et al. (2002) Gene transfer of thromboxane A(2) synthase and prostaglandin I(2) synthase antithetically altered tumor angiogenesis and tumor growth. Cancer Res., 62, 63–66. 39. Kim,J. et al. (2005) Suppression of prostate tumor cell growth by stromal cell prostaglandin D synthase-derived products. Cancer Res., 65, 6189–6198. 40. Vergote,I. et al. (1985) Prostaglandin F2 alpha in benign and malignant breast tumours. Br. J. Cancer, 51, 827–836. 41. Sivula,A. et al. (2005) Association of cyclooxygenase-2 and matrix metalloproteinase-2 expression in human breast cancer. Breast Cancer Res. Treat., 89, 215–220. 42. DeNardo,D.G. et al. (2009) CD4(þ) T cells regulate pulmonary metastasis of mammary carcinomas by enhancing protumor properties of macrophages. Cancer Cell, 16, 91–102. 43. Mantovani,A. et al. (2006) Role of tumor-associated macrophages in tumor progression and invasion. Cancer Metastasis Rev., 25, 315–322. 44. Leek,R. et al. (2002) Tumor-associated macrophages in breast cancer. J. Mammary Gland Biol. Neoplasia, 7, 177–189. 45. Gouon-Evans,V. et al. (2002) Requirement of macrophages and eosinophils and their cytokines/chemokines for mammary gland development. Breast Cancer Res., 4, 155–164. 46. Mantovani,A. et al. (2002) Macrophage polarization: tumor-associated macrophages as a paradigm for polarized M2 mononuclear phagocytes. Trends Immunol., 23, 549–555. 47. van der Bij,G. et al. (2008) Tumor infiltrating macrophages reduce development of peritoneal colorectal carcinoma metastases. Cancer Lett. 48. Kim,D. et al. (2008) High tumour islet macrophage infiltration correlates with improved patient survival but not with EGFR mutations, gene copy number or protein expression in resected non-small cell lung cancer. Br. J. Cancer, 98, 1118–1124. 49. Jadus,M. et al. (1996) Macrophages can recognize and kill tumor cells bearing the membrane isoform of macrophage colony-stimulating factor. Blood, 87, 5232–5241. 50. Yao,C. et al. (2009) Prostaglandin E2-EP4 signaling promotes immune inflammation through Th1 cell differentiation and Th17 cell expansion. Nat. Med., 15, 633–640. 51. Boniface,K. et al. (2009) Prostaglandin E2 regulates Th17 cell differentiation and function through cyclic AMP and EP2/EP4 receptor signaling. J. Exp. Med., 206, 535–548. Received April 28, 2011; revised June 20, 2011; accepted July 5, 2011 1449