Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
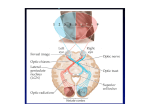
Review of Visual System We can’t go over every aspect of the visual system in a single lecture, so I’m going to assume you have covered this in general in previous courses, and will review some elements that are key to this course. 1) Anatomy, Lateralization, and Depth. Stimulation at the retina leads to activation of retinal ganglion cells, which project to a part of the thalamus called the lateral geniculate nucleus (LGN). Remember that the optics of the eye cause light from the right visual field to strike the left retina and vice versa. Ganglion cells from the medial retinas (the parts close to the nose) cross over to the opposite LGN. As a result, you have everything from the left visual field activating the right LGN, and vice versa. But at this point, information from the two eyes is not yet combined, but instead is separated in different layers of magnocellular (M) and parvocellular (P) cells.. The right LGN then projects to the right side of the primary visual cortex (V1), located along the calcarine fissure at the back of occipital cortex (and left to left). Thus this lateralization remains in the cortex and is seen through higher levels of visual cortex as well (thought question: so why don’t we see things backwards?). Information from the left and right eyes is combined at the level of V1, giving rise to a form of depth perception called binocular disparity. However, other monocular cues (e.g., occlusion, parallax, size constancy, linear convergence and texture) are equally important. 2) Retinotopy. Because of the structure of the eye and the orderly way in which light falls on the back of the eye, activation of cells in the retina likewise forms an orderly, i.e. topographic map of space known as a retinotopic map. The map is over-represented (and color) at the fovea, where cone cells are packed closely together, and under-represneted (and relatively color instenstive) in the periphery where one has more scattered receptors, mainly of the rod type. These project indirectly to local gangion cells, which we have seen already project to the LGN. This distorted retinotopic map is preserved in LGN and V1, but starts to break down at higher levels (V2, V3, V4) and its controversial if any retinotopy remains at the highest levels (see below), except in terms of general lateralization. Retinotopy does not exist so that we have a picture of the world in our heads, rather it may have several advantages: if most connections between cells are local, then retinotopy reduces connection lengths (and thus brain volume) while increasing speed of interactions. It may also play a role in certain ‘wave’-like electrical phenomena that my coordinate activation across populations of neurons. Such population phenomena are not so obvious when one records action potentials, but are more evident when one records ‘local field potentials’ that reveal the sum of local synaptic activity, or images visual cortex in other ways. 3) Receptive Fields (RF). The RF is the area in space where a stimulus can modulate activity of a particular neuron. This activity is normally quantified by measuring the rate of action potentials from a single neuron. This modulation could be either excitatory or inhibitory. (There are also ‘non classical’ RFs: points in space where a stimulus will not change activity, but will change the effect of a stimulus in the classic RF.) Visual RFs are generally located in a particular part of retinotopic space and vary in size and shape. In the classic story, Retinal Ganglion cells and LGN cells essentially have donut-like RFs with excitatory centres and inhibitory surrounds, which are small near the fovea and bigger toward the periphery. Hubel and Weisel won the Nobel Prize for showing that in V1, these RFs assume more linear characteristics, e.g., ‘simple cells’ that respond to lines with a certain orientation in a certain part of space, ‘complex cells’ that require the line to be moving one way or the other othrogonal to its long axis, or hypercomplex cells that require a certain line length. Moder techniques tend to be more computational, for example looking at frequency and principle component analysis of the information contained in cells. 4) Functional Specialization. Even at the level of the retina, ganglion cells have different functional properties, e.g. comparing p and m ganglion cells. Confusingly, p cells project to M cells in the LGN and m cells project to P cells, where from hereon they are called the M and P pathway (based on the LGN nomenclature). P cells tend to have smaller, colorsensitive receptive fields and less temporal resolution (and thus are thought to be good for discriminating certain features of objects), whereas M cells have larger color-insensitive receptive fields and are very temporally sensitive (and thus are thought to be more useful for spatial perception and motion processing). This dichotomy tends to be preserved in cortex, although in more abstract ways. Ungerleider and Mishkin pointed out that a ‘dorsal stream’ of vision, running from occipital cortex to parietal cortex, contained mostly spatiallyresponsive neurons with little feature responsiveness, whereas the ‘ventral stream’, running from occipital cortex through temporal cortex has neurons that are highly sensitive to features. Further as one goes further along this stream the features become more complex, e.g. coding for rudimentary shape components like corners in V4 to (sometimes) specific objects, independent of location, in IT (inferotemporal). An intermediate area in the human, LO, seems particularly important for object analysis. This gave rise to the notion of a ‘where’ and ‘what’ pathways. Goodale and Milner revised this to ‘how’ and ‘what’ or action and perception pathways. This is based on observations such as deficits in patients with these areas, or the fact that many dorsal stream neurons also show action responses and motor response fields for different parts of the body (we will return to this next week). The main difference between these two accounts is that the first considers the type of visual input to the stream, whereas the second considers the output: what this information is used for. Not everything fits neatly within these schemes; for example V5 (MT, or MT+ in humans) is a major motion sensitive area that contributes to both perception and action, so it might be considered an input module to both. Further, each of the streams has many different functional modules (some of which we will consider in this course), each having different complex neuron types. Neverthless, the dorsal-ventral distinction has proven over the years to be a powerful heuristic to understand the major aspects of high-level vision.